
Soil Respiration Characteristics and Influencing Factors for Apple Orchards in Different Regions on the Loess Plateau of Shaanxi Province

Abstract
:1. Introduction
2. Materials and Methods
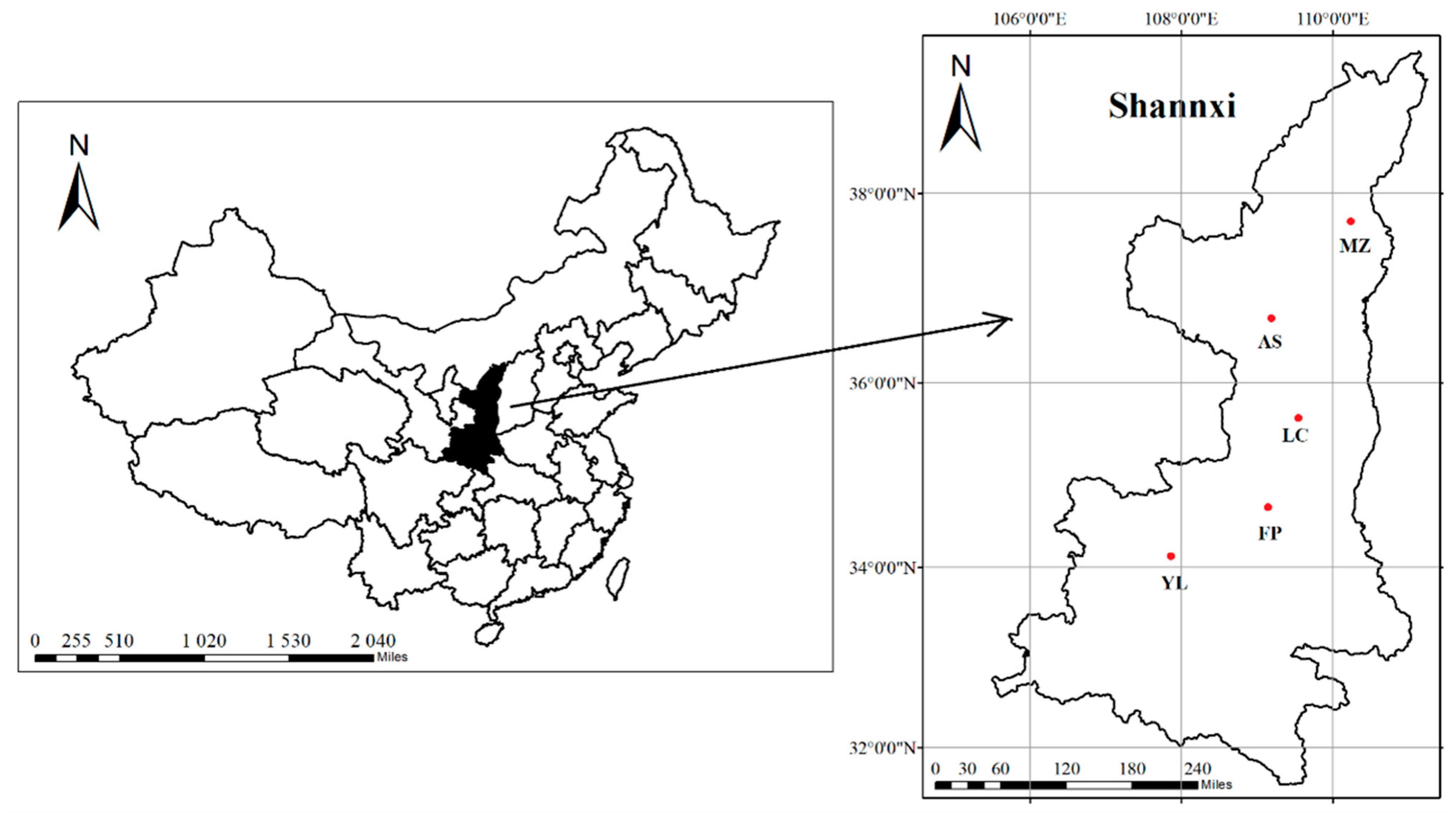
2.1. Area Description
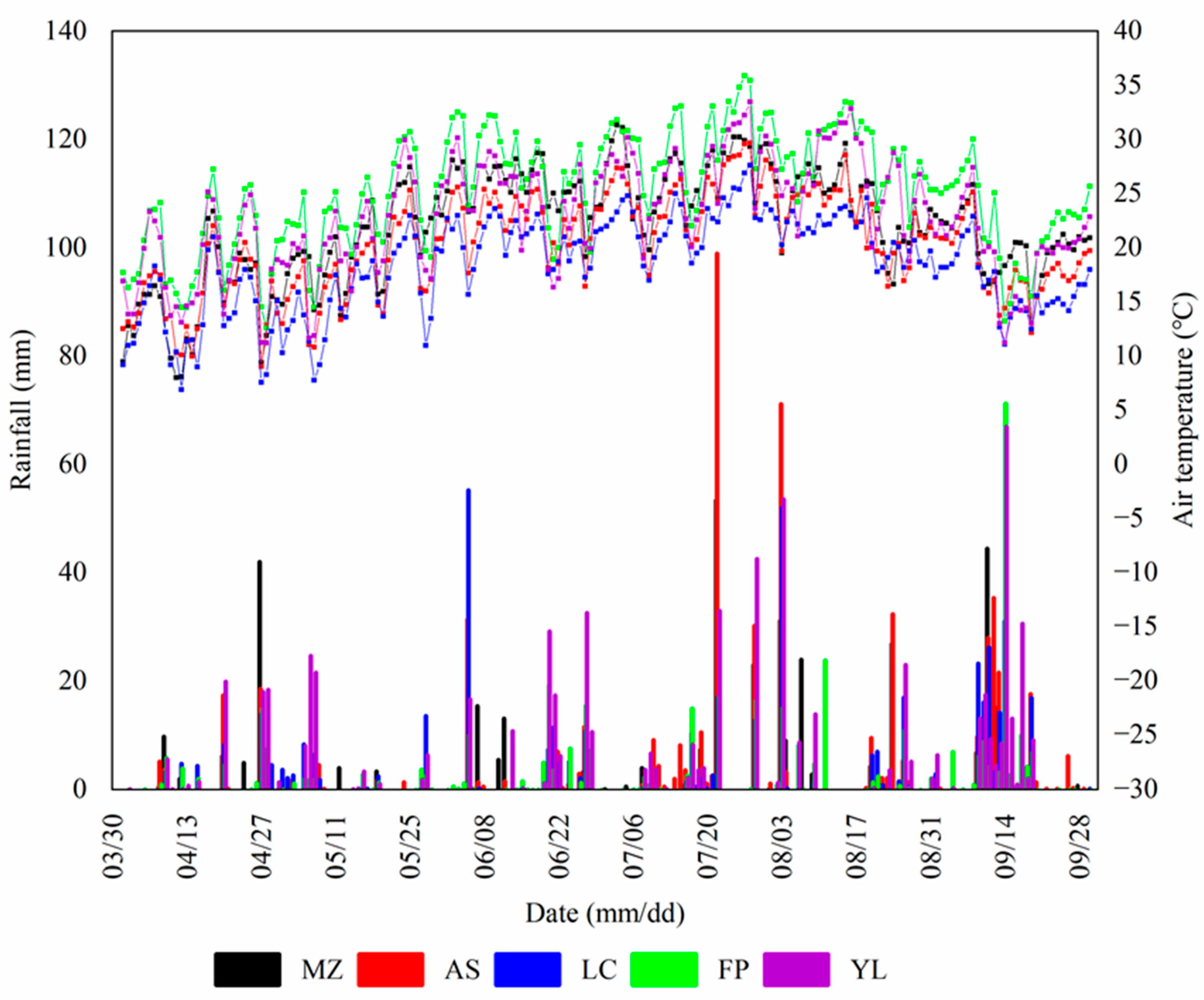
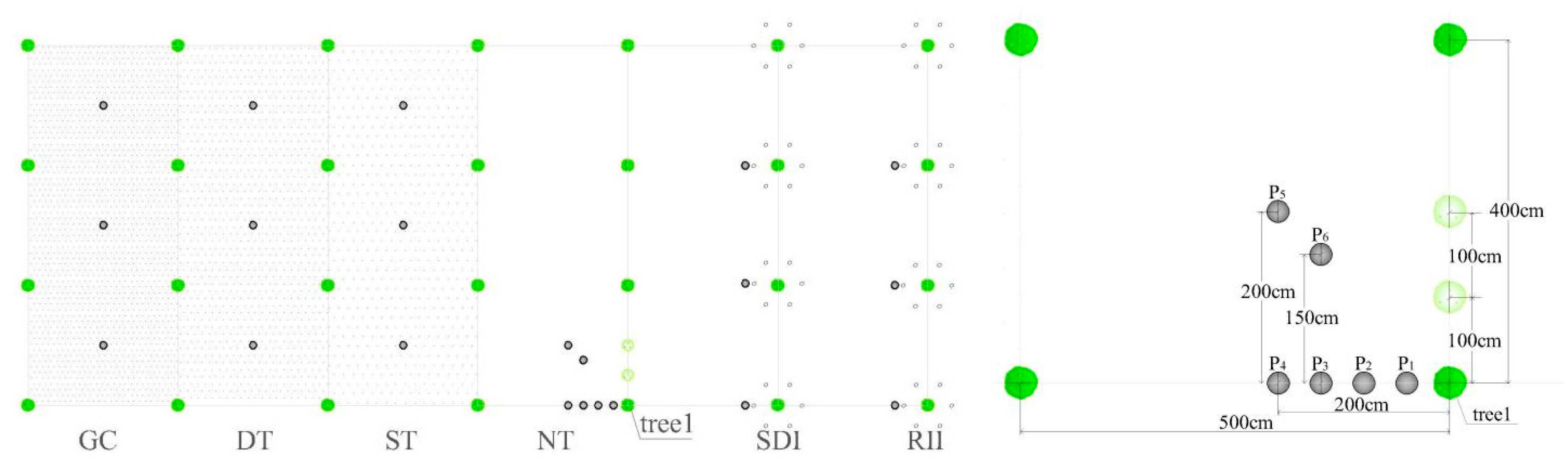
2.2. Experimental Design and Measurement
2.3. Data Analysis
3. Result
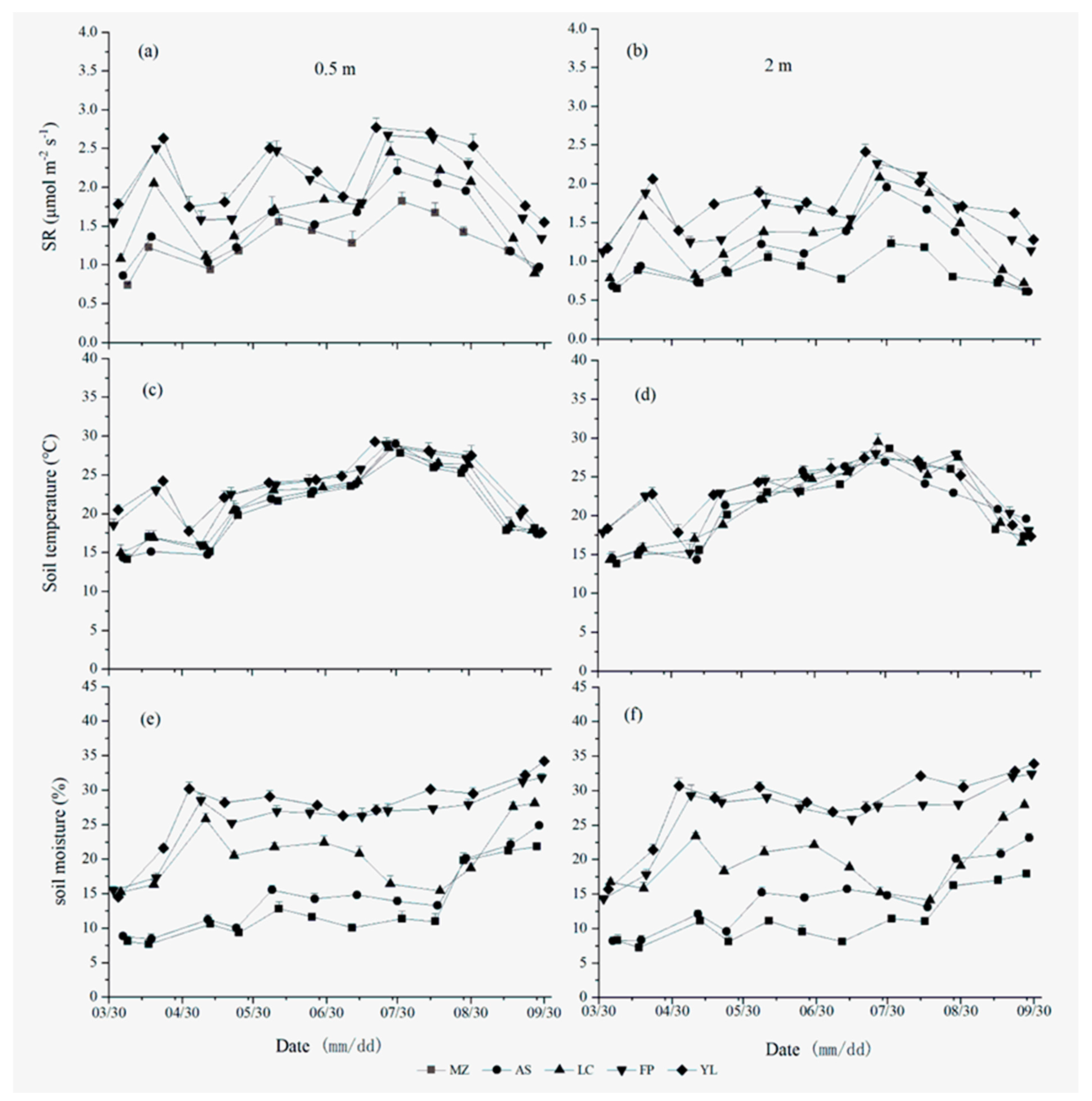
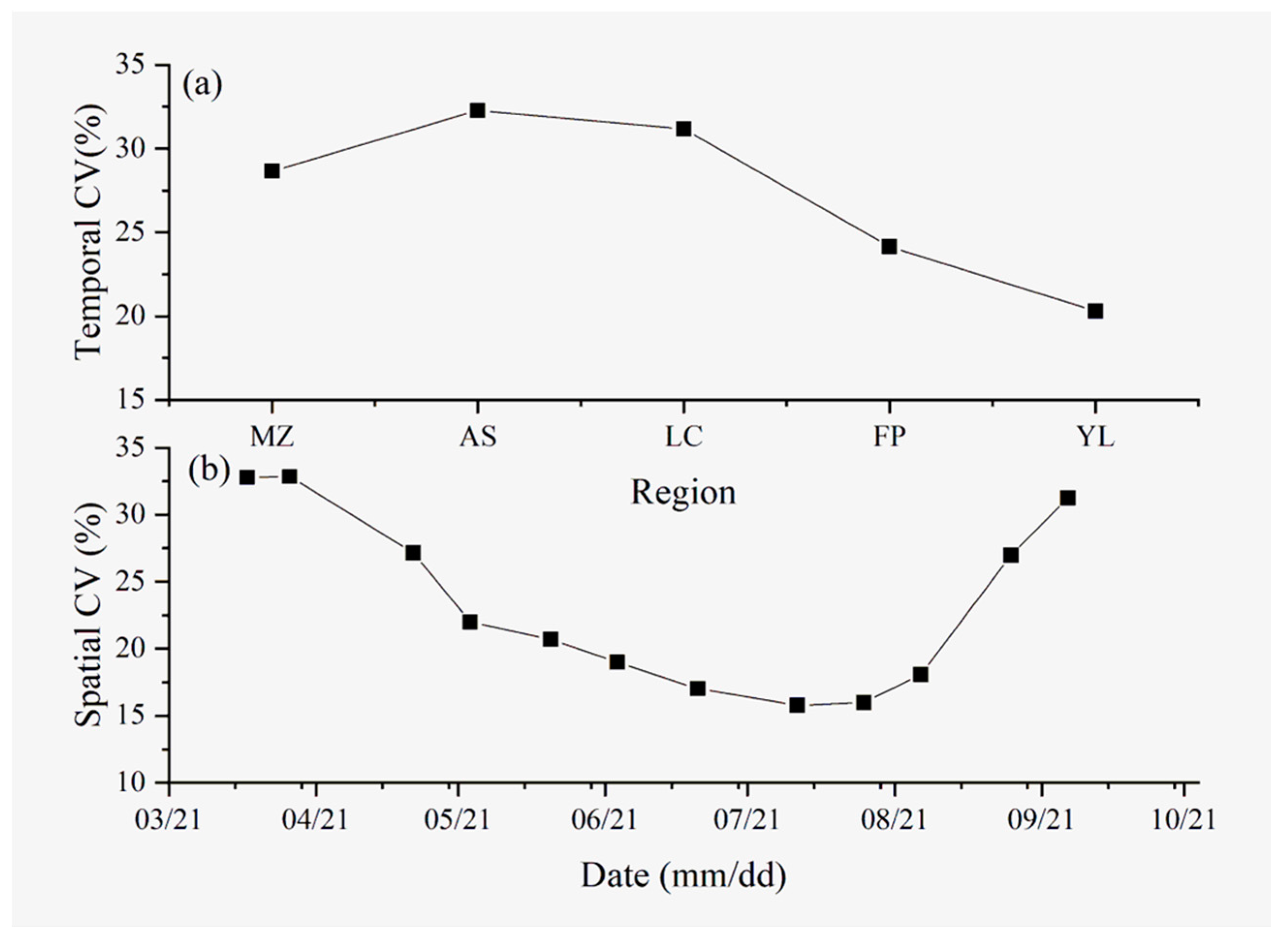
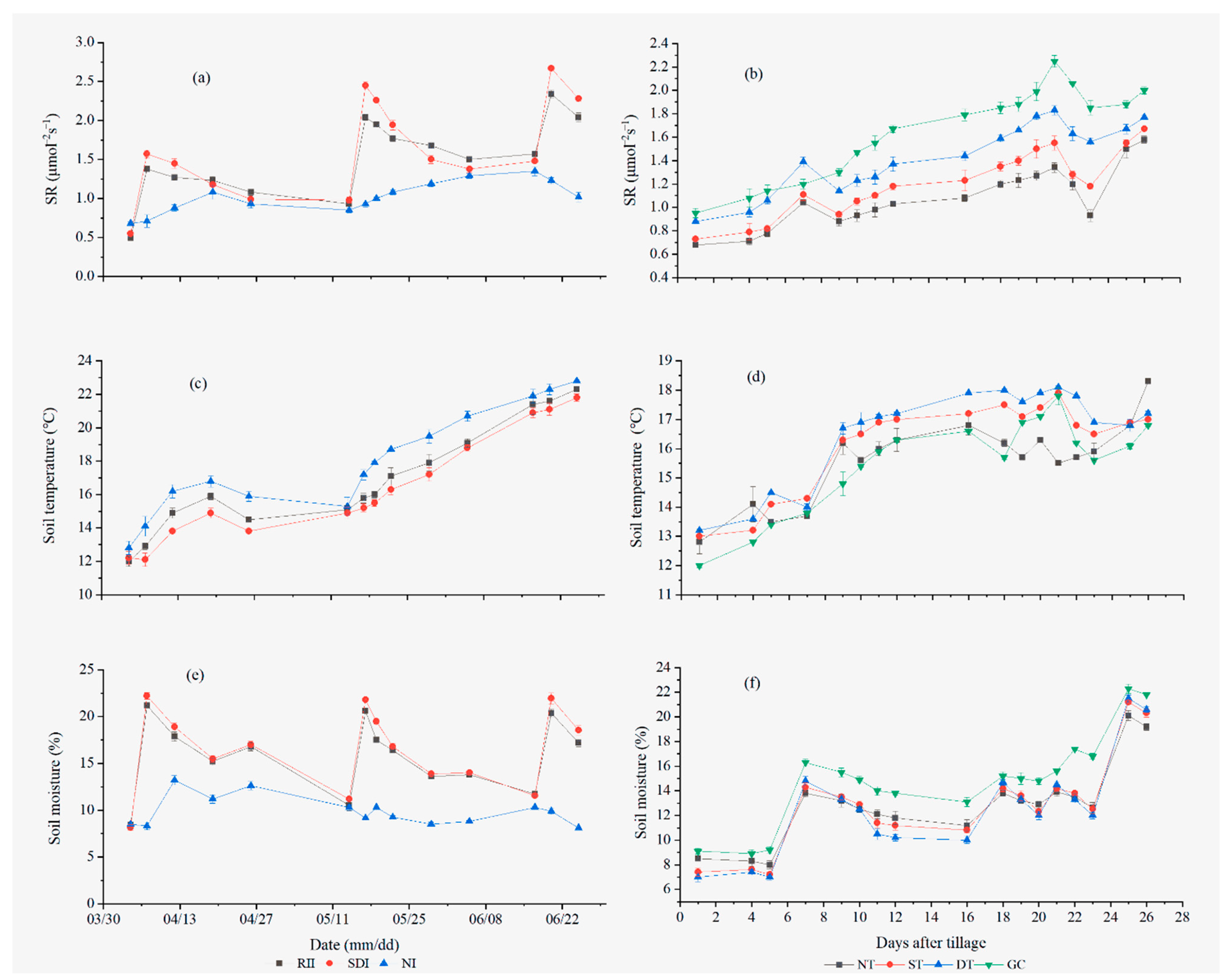
3.1. Differences in Temporal Variation in the SR in the Apple Orchards in Different Areas
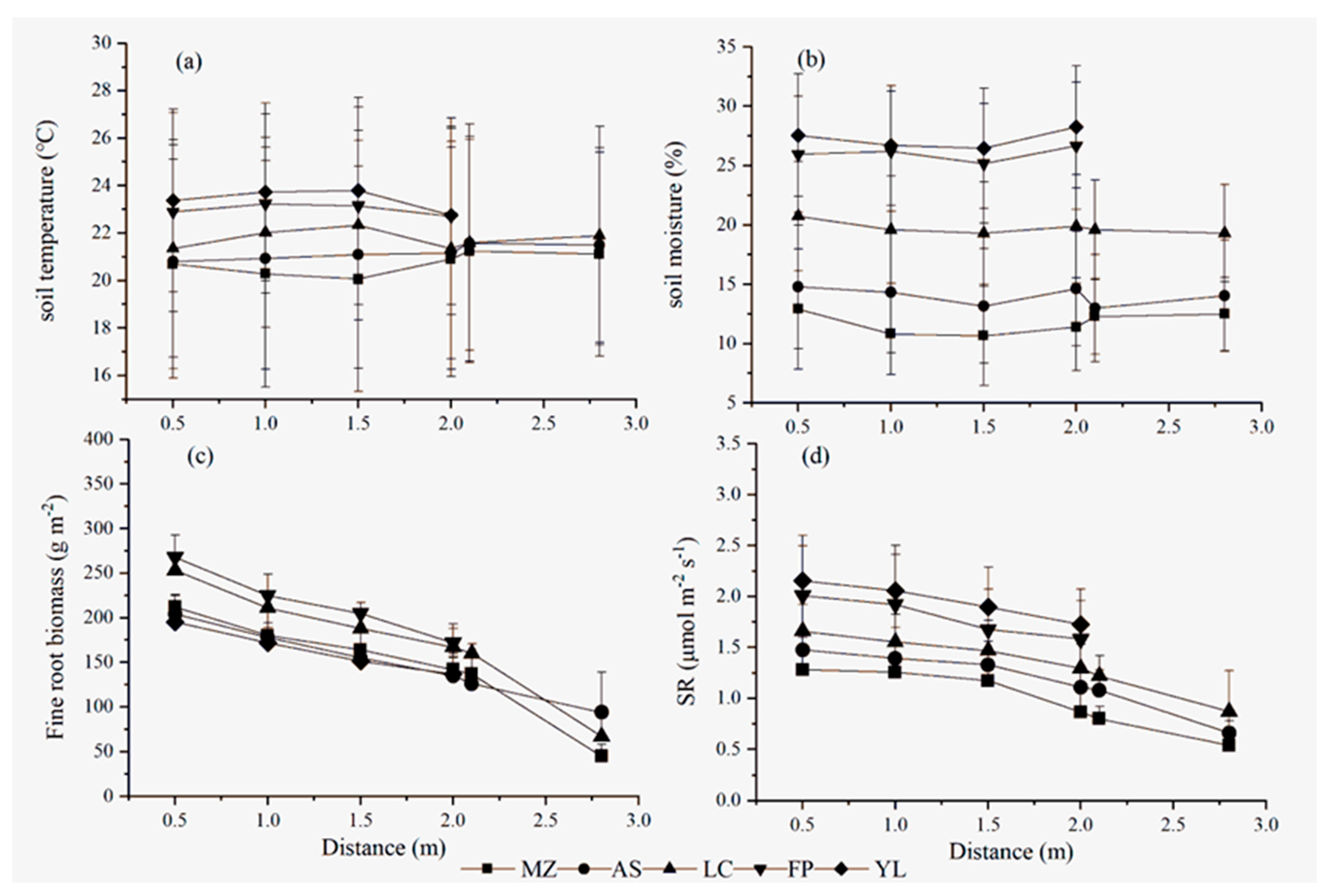
3.2. Differences in Spatial Variation in the Soil Respiration in the Apple Orchards in Different Areas
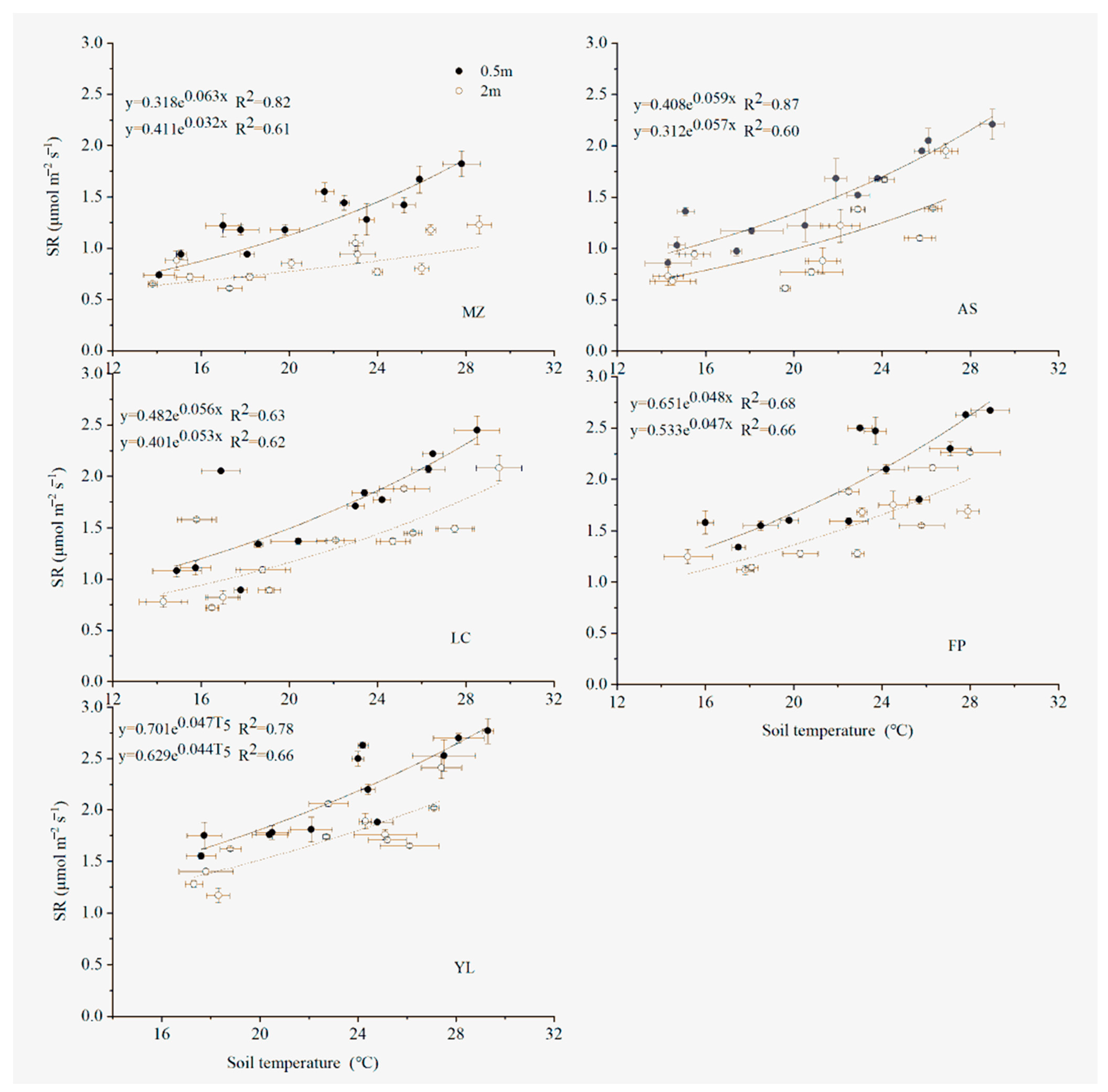
3.3. Temporal SR Related to Soil Temperature and Soil Moisture
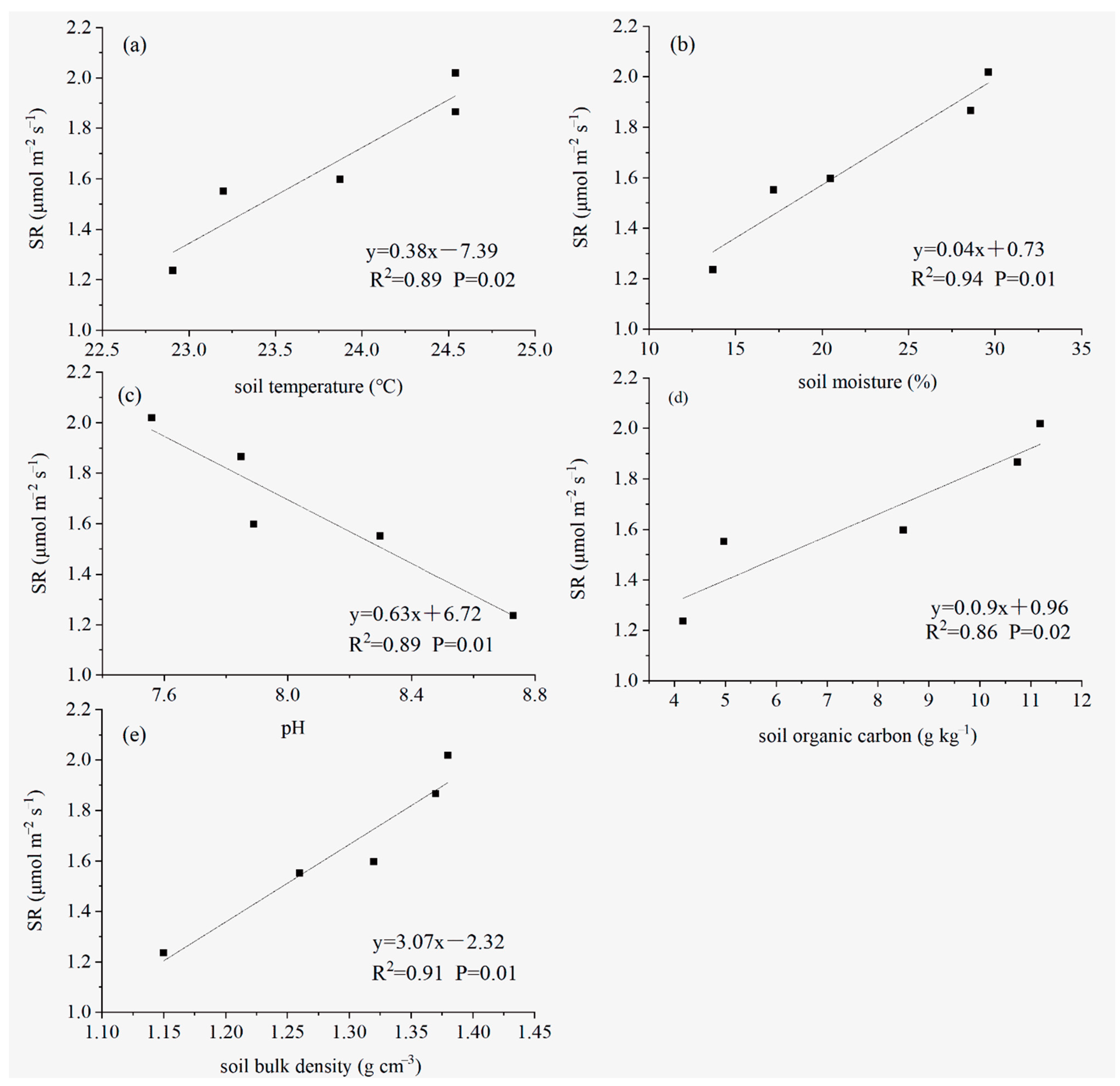
3.4. The Regional Variation in SR Is Related to the Soil Temperature, Soil Moisture, Soil Organic Carbon, pH and Soil Bulk Density
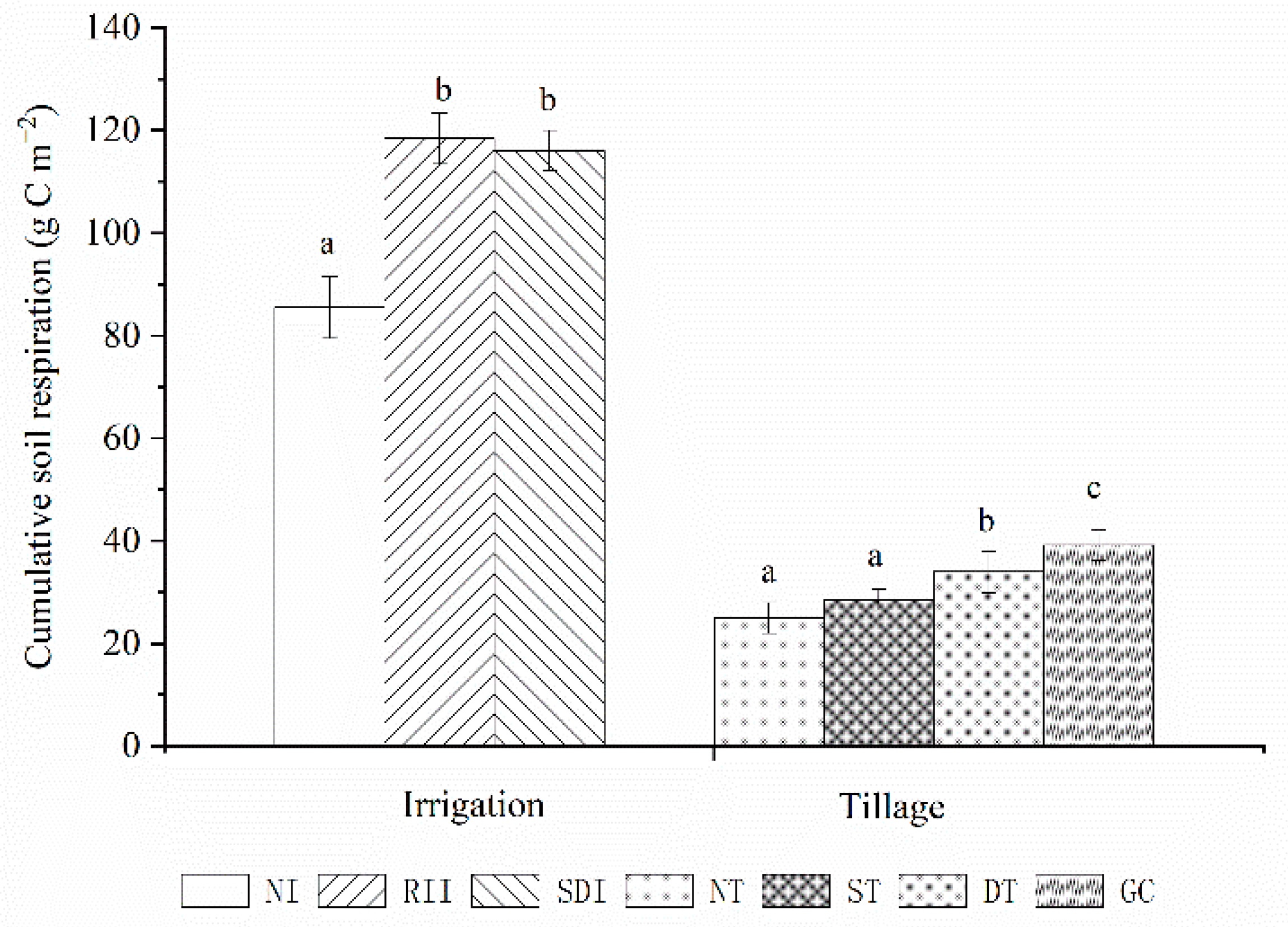
3.5. Effects of Agricultural Management Practices on SR
4. Discussion
4.1. Temporal Variability of SR
4.2. Spatial Variability of SR
4.3. SR under Different Soil Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Guo, S.; Zhang, Y.; Liu, Q.; Wang, R.; Wang, Z.; Li, N.; Li, R. Changes in temperature sensitivity of soil respiration in the phases of a three-year crop rotation system. Soil Tillage Res. 2015, 150, 139–146. [Google Scholar] [CrossRef]
- Adachi, M.; Ito, A.; Yonemura, S.; Takeuchi, W. Estimation of global soil respiration by accounting for land-use changes derived from remote sensing data. J. Environ. Manag. 2017, 200, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Song, Y.; Zhang, W.; He, S. Spatio-temporal Variation of Soil Respiration and Its Driving Factors in Semi-arid Regions of North China. Chin. Geogr. Sci. 2018, 28, 12–24. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.D. Spatial–temporal variation in soil respiration in an oak–grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 2005, 73, 183–207. [Google Scholar] [CrossRef]
- Ngao, J.; Epron, D.; Delpierre, N.; Bréda, N.; Granier, A.; Longdoz, B. Spatial variability of soil CO2 efflux linked to soil parameters and ecosystem characteristics in a temperate beech forest. Agric. For. Meteorol. 2012, 154–155, 136–146. [Google Scholar] [CrossRef]
- Zhou, X.-G.; Zhang, Y.-J.; Nan, Y.-F.; Liu, Q.-F.; Guo, S.-L. Differences in soil respiration between cropland and grassland ecosystems and factors influencing soil respiration on the Loess Plateau. Environ. Sci. 2013, 34, 1026–1033. [Google Scholar]
- Gong, J.; Xu, S.; Wang, Y.; Luo, Q.; Liu, M.; Zhang, W. Effect of irrigation on the soil respiration of constructed grasslands in Inner Mongolia, China. Plant Soil 2015, 395, 159–172. [Google Scholar] [CrossRef]
- Abido, W.; Hadházy, Á.; Henzsel, I. Effect of tillage method on carbon-dioxide emission andsoil properties under two soil surface levels. Acta Ecol. Sin. 2020, 40, 210–213. [Google Scholar] [CrossRef]
- Wang, R.; Guo, S.; Jiang, J.; Wu, D.; Li, N.; Zhang, Y.; Liu, Q.; Li, R.; Wang, Z.; Sun, Q.; et al. Tree-scale spatial variation of soil respiration and its influence factors in apple orchard in Loess Plateau. Nutr. Cycl. Agroecosyst. 2015, 102, 285–297. [Google Scholar] [CrossRef]
- María, M.G.R.; Gregorio, E.; Bernardo, B.M.G.; Pedro, A.N.; Alain, B. Spatial variability of soil CO2 efflux in drip-irrigated old and young citrus orchards and its dependence on biotic and abiotic factors. Geoderma 2017, 294, 29–37. [Google Scholar] [CrossRef]
- González-Real, M.M.; Martin-Gorriz, B.; Egea, G.; Nortes, P.A.; Baille, A. Characterization and modelling of soil CO2 efflux in old and young irrigated citrus orchards. Catena 2018, 162, 376–385. [Google Scholar] [CrossRef]
- Tedeschi, V.; Rey, A.; Manca, G.; Valentini, R.; Jarvis, P.G.; Borghetti, M. Soil respiration in a Mediterranean oak forest at different developmental stages after copping. Glob. Chang. Biol. 2006, 12, 110–121. [Google Scholar] [CrossRef]
- Ceccon, C.; Panzacchi, P.; Scandellari, F.; Prandi, L.; Ventura, M.; Russo, B.; Millard, P.; Tagliavini, M. Spatial and temporal effects of soil temperature and moisture and the relation to fine root density on root and soil respiration in a mature apple orchard. Plant Soil 2011, 342, 195–206. [Google Scholar] [CrossRef]
- Luo, Y.Q.; Zhou, X.H. Soil Respiration and the Environment; Academic Press: London, UK, 2006; pp. 257–305. [Google Scholar]
- Wang, C.; Huang, R.; Yang, Z.-J.; Liu, Q.; Chen, G.-S.; Wan, X.-H. Dynamic changes of soil respiration in Citrus reticulata and Castanea henryi orchards in Wanmulin Nature Reserve, Fujian Province of East China. Chin. J. Appl. Ecol. 2012, 23, 1469–1475. [Google Scholar]
- Yan, J.-X.; Hao, Z.; Jing, X.-K.; Li, H.-J. Interannual Variations of Soil Respiration and Its Temperature Sensitivity in an Orchard in Jinci Region of Taiyuan City. Environ. Sci. 2016, 37, 3625–3633. [Google Scholar]
- Scandellari, F.; Zanotelli, D.; Ceccon, C.; Bolognesi, M.; Montagnani, L.; Cassol, P.; Melo, G.; Tagliavini, M. Enhancing prediction accuracy of soil respiration in an apple orchard by integrating photosynthetic activity into a temperature-related model. Eur. J. Soil Biol. 2015, 70, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.S.; Li, L.H.; Han, X.G.; Yan, Z.D.; Wang, Y.F.; Zhang, Y.; Xiong, X.G.; Chen, S.P.; Zhang, L.X.; Gao, Y.Z.; et al. Temperature sensitivity of soil respiration in relation to soil moisture in 11 communities of typical temperate steppe in Inner Mongolia. Acta Ecol. Sin. 2004, 24, 831–836. [Google Scholar] [CrossRef]
- Millikin, C.S.; Bledsoe, C.S. Biomass and distribution of fine and coarse roots from blue oak (Quercus douglasii) trees in the northern Sierra Nevada foothills of California. Plant Soil 1999, 214, 27–38. [Google Scholar] [CrossRef]
- Gan, Z.-T.; Zhou, Z.-C.; Liu, W.-Z. Vertical Distribution and Seasonal Dynamics of Fine Root Parameters for Apple Trees of Different Ages on the Loess Plateau of China. Agric. Sci. China 2010, 9, 46–55. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; Shi, Z.; Lu, L.; Zeng, J.; Ming, A.; Tang, J.; Yu, H. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. For. Ecol. Manag. 2013, 300, 4–13. [Google Scholar] [CrossRef]
- Wang, R.; Guo, S.-L.; Liu, Q.-F.; Zhang, Y.-J.; Jiang, J.-S.; Guo, H.-M.; Li, R.-J. Variation characteristic in soil respiration of apple orchard and its biotic and abiotic influencing factors. Environ. Sci. 2014, 35, 1915–1921. [Google Scholar]
- Sheng, H.; Yang, Y.; Yang, Z.; Chen, G.; Xie, J.; Guo, J.; Zou, S. The dynamic response of soil respiration to land-use changes in subtropical China. Glob. Chang. Biol. 2010, 16, 1107–1121. [Google Scholar] [CrossRef]
- Wiseman, P.; Seiler, J.R. Soil CO2 efflux across four age classes of plantation loblolly pine (Pinus taeda L.) on the Virginia Piedmont. For. Ecol. Manag. 2004, 192, 297–311. [Google Scholar] [CrossRef]
- Sainju, U.M.; Jabro, J.D.; Stevens, W.B. Soil Carbon Dioxide Emission and Carbon Content as Affected by Irrigation, Tillage, Cropping System, and Nitrogen Fertilization. J. Environ. Qual. 2008, 37, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichstein, M.R.; Ubke, J.E.S.; Angeli, A.C.; Tenhunen, J.D. Does the temperature sensitivity of decomposition of soil organic matter depend upon water content, soil horizon, or incubation time? Glob. Chang. Biol. 2005, 11, 1754–1767. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Wang, T.; Sun, J.; Shen, Z. Temperature sensitivity of soil respiration in different ecosystems in China. Soil Biol. Biochem. 2009, 41, 1008–1014. [Google Scholar] [CrossRef]
- Lavigne, M.B.; Boutin, R.; Foster, R.J.; Goodine, G.; Bernier, P.Y.; Robitaille, G. Soil respiration responses to temperature are controlled more by roots than by decomposition in balsam fir ecosystems. Can. J. For. Res. 2003, 33, 1744–1753. [Google Scholar] [CrossRef]
- Zheng, Z.-M.; Yu, G.-R.; Fu, Y.-L.; Wang, Y.-S.; Sun, X.-M.; Wang, Y.-H. Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study. Soil Biol. Biochem. 2009, 41, 1531–1540. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Y.; Zou, J.; Shen, Q.; Hu, Z.; Qin, Y.; Chen, H.; Pan, G. Modeling interannual variability of global soil respiration from climate and soil properties. Agric. For. Meteorol. 2010, 150, 590–605. [Google Scholar] [CrossRef]
- Shi, P.; Qin, Y.; Liu, Q.; Zhu, T.; Li, Z.; Li, P.; Ren, Z.; Liu, Y.; Wang, F. Soil respiration and response of carbon source changes to vegetation restoration in the Loess Plateau, China. Sci. Total. Environ. 2020, 707, 135507. [Google Scholar] [CrossRef]
- Yang, J.-F.; Yang, X.-N.; Wang, J.-H.; Duan, Y.-M.; Qi, X.-N.; Zhang, L.-S. Characteristics of CO2 Flux in a Mature Apple (Malus demestica) Orchard Ecosystem on the Loess Plateau. Environ. Sci. 2018, 39, 2339–2350. [Google Scholar]
- Zhang, H.; Huang, Y.-M.; Qi, J.-H.; An, S.-S. Effects of temperature and moisture on soil CO2 release potential under three typical land use types in the hill-gully area of the Loess Plateau. Chin. J. Eco-Agric. 2011, 19, 731–737. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, S.; Liu, Q.; Jiang, J.; Wang, R.; Li, N. Responses of soil respiration to land use conversions in degraded ecosystem of the semi-arid Loess Plateau. Ecol. Eng. 2015, 74, 196–205. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Q.; Wang, Y.; Zheng, W.; Yao, L.; Hu, Y.; Guo, S. Contrasting responses of soil respiration and temperature sensitivity to land use types: Cropland vs. apple orchard on the Chinese Loess Plateau. Sci. Total Environ. 2018, 621, 425–433. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Q.; Han, X.; Wan, S.; Li, L. Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Buchmann, N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands. Soil Biol. Biochem. 2000, 32, 1625–1635. [Google Scholar] [CrossRef]
- Chen, G.S.; Yang, Y.S.; Lü, P.P.; Zhang, Y.P.; Qian, X.L. Regional patterns of soil respiration in China’s forests. Acta Ecol. Sin. 2008, 28, 1748–1760. [Google Scholar] [CrossRef]
- Iqbal, J.; Hu, R.G.; Du, L.J.; Lu, L.; Lin, S.; Chen, T.; Ruan, L.L. Differences in soil CO2 flux between different land use types in mid-subtropical China. Soil Biol. Biochem. 2008, 40, 2324–2333. [Google Scholar] [CrossRef]
- Zhao, G.D.; Wang, B.; Yang, J.; Zhang, Z.J. LI-8100 Automated Soil CO2 Flux System and Its Application. Meteorol. Sci. Technol. 2005, 33, 363–366. [Google Scholar] [CrossRef]
- Lu, R.K. Analytical Methods for Soil Agrochemistry; Chinese Agricultural Science and Technology Publishing House: Beijing, China, 2000. [Google Scholar]
- Liu, G.S. Soil Physical and Chemical Analysis and Profile Description; China Standard Press: Beijing, China, 1996. [Google Scholar]
- Wang, Y.; Zhang, L.; Mu, Y.; Liu, W.; Guo, F.; Chang, T. Effect of a root-zone injection irrigation method on water productivity and apple production in a semi-arid region in north-western China. Irrig. Drain. 2019, 69, 74–85. [Google Scholar] [CrossRef]
- Tian, X.-Y.; Tu, L.-H.; Hu, T.-X.; Zhang, J.; He, Y.-Y.; Xiao, Y.-L. Characteristics of soil respiration components and their temperature sensitivity in a Pleioblastus amarus plantation in rainy area of West China. Chin. J. Appl. Ecol. 2012, 23, 293–300. [Google Scholar] [CrossRef]
- Gnanamoorthy, P.; Selvam, V.; Ramasubramanian, R.; Nagarajan, R.; Chakraborty, S.; Burman, P.K.D.; Karipot, A. Diurnal and seasonal patterns of soil CO2 efflux from the Pichavaram mangroves, India. Environ. Monit. Assess. 2019, 191, 258. [Google Scholar] [CrossRef]
- Luo, J.; Chen, Y.; Wu, Y.; Shi, P.; She, J.; Zhou, P. Temporal-Spatial Variation and Controls of Soil Respiration in Different Primary Succession Stages on Glacier Forehead in Gongga Mountain, China. PLoS ONE 2012, 7, e42354. [Google Scholar] [CrossRef] [Green Version]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Zanotelli, D.; Montagnani, L.; Manca, G.; Scandellari, F.; Tagliavini, M. Net ecosystem carbon balance of an apple orchard. Eur. J. Agron. 2015, 63, 97–104. [Google Scholar] [CrossRef]
- Saiz, G.; Green, C.; Butterbach-Bahl, K.; Kiese, R.; Avitabile, V.; Farrell, E.P. Seasonal and spatial variability of soil respiration in four Sitka spruce stands. Plant Soil 2006, 287, 161–176. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, P.; Wang, H.; Zhou, G.; Wu, J.; Yang, F.; Qian, X. Seasonal soil CO2 efflux dynamics after land use change from a natural forest to Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2011, 262, 1131–1137. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J.; Zhang, Q. Soil respiration in six temperate forests in China. Glob. Chang. Biol. 2006, 12, 2103–2114. [Google Scholar] [CrossRef]
- Jia, B.; Zhou, G.; Wang, Y.; Wang, F.; Wang, X. Effects of temperature and soil water-content on soil respiration of grazed and ungrazed Leymus chinensis steppes, Inner Mongolia. J. Arid. Environ. 2006, 67, 60–76. [Google Scholar] [CrossRef]
- Xie, H.-H.; Fan, J.; Qi, L.-B.; Hao, M.-D. Seasonal characteristics of soil respiration and affecting factors under typical vegetations in the water-wind erosion crisscross region of the Loess Plateau. Environ. Sci. 2010, 31, 164–172. [Google Scholar] [CrossRef]
- Zhao, Z.M.; Zhao, C.Y.; Yan, Y.Y.; Li, J.Y.; Li, J.; Shi, F.Z. Interpreting the dependence of soil respiration on soil temperature and moisture in an oasis cotton field, central Asia. Agric. Ecosyst. Environ. 2013, 168, 46–52. [Google Scholar] [CrossRef]
- Hanson, P.; Edwards, N.; Garten, C.; Andrews, J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Smith, D.L.; Johnson, L. Vegetation-mediated changes in microclimate reduce soil respiration as woodlands expand into grasslands. Ecology 2004, 85, 3348–3361. [Google Scholar] [CrossRef]
- da Cunha, J.M.; Campos, M.C.C.; Gaio, D.C.; de Souza, Z.M.; Soares, M.D.R.; da Silva, D.M.P.; Simões, E.L. Spatial variability of soil respiration in Archaeological Dark Earth areas in the Amazon. Catena 2018, 162, 148–156. [Google Scholar] [CrossRef]
- Yim, M.H.; Joo, S.J.; Shutou, K.; Nakane, K. Spatial variability of soil respiration in a larch plantation: Estimation of the number of sampling points required. For. Ecol. Manag. 2003, 175, 585–588. [Google Scholar] [CrossRef]
- Barba, J.; Yuste, J.C.; Martínez-Vilalta, J.; Lloret, F. Drought-induced tree species replacement is reflected in the spatial variability of soil respiration in a mixed Mediterranean forest. For. Ecol. Manag. 2013, 306, 79–87. [Google Scholar] [CrossRef]
- Hernández-Montes, E.; Escalona, J.; Tomás, M.; Medrano, H. Influence of water availability and grapevine phenological stage on the spatial variation in soil respiration. Aust. J. Grape Wine Res. 2017, 23, 273–279. [Google Scholar] [CrossRef]
- Wang, W.; Zeng, W.; Chen, W.; Yang, Y.; Zeng, H. Effects of Forest Age on Soil Autotrophic and Heterotrophic Respiration Differ between Evergreen and Deciduous Forests. PLoS ONE 2013, 8, e80937. [Google Scholar] [CrossRef] [Green Version]
- Rodeghiero, M.; Cescatti, A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps. Glob. Chang. Biol. 2005, 11, 1024–1041. [Google Scholar] [CrossRef]
- Merbold, L.; Ziegler, W.; Mukelabai, M.M.; Kutsch, W.L. Spatial and temporal variation of CO2 efflux along a disturbance gradient in a miombo woodland in Western Zambia. Biogeosciences 2011, 8, 147–164. [Google Scholar] [CrossRef] [Green Version]
- Smith, V. Soil respiration and its determinants on a sub-Antarctic island. Soil Biol. Biochem. 2003, 35, 77–91. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Y.; Jia, B.; Wang, F.; Zhou, G. Comparison of soil respiration among three temperate forests in Changbai Mountains, China. Can. J. For. Res. 2010, 40, 788–795. [Google Scholar] [CrossRef]
- Chen, H.; Tian, H.-Q. Does a General Temperature-Dependent Q10 Model of Soil Respiration Exist at Biome and Global Scale? J. Integr. Plant Biol. 2005, 47, 1288–1302. [Google Scholar] [CrossRef]
- Conant, R.T.; Dalla-Betta, P.; Klopatek, C.C.; Klopatek, J.M. Controls on soil respiration in semiarid soils. Soil Biol. Biochem. 2004, 36, 945–951. [Google Scholar] [CrossRef]
- Smith, V. Moisture, carbon and inorganic nutrient controls of soil respiration at a sub-Antarctic island. Soil Biol. Biochem. 2005, 37, 81–91. [Google Scholar] [CrossRef]
- Janssens, I.A.; Pilegaard, K. Large seasonal changes in Q10 of soil respiration in a beech forest. Glob. Chang. Biol. 2003, 9, 911–918. [Google Scholar] [CrossRef]
- Yang, Q.P.; Xu, M.; Liu, H.S.; Wang, J.S.; Liu, L.X.; Chi, Y.G.; Zheng, Y.P. Impact factors and uncertainties of the temperature sensitivity of soil respiration. Acta Ecol. Sin. 2011, 31, 2301–2311. [Google Scholar]
- Parkin, T.B.; Kaspar, T.C. Temperature Controls on Diurnal Carbon Dioxide Flux. Soil Sci. Soc. Am. J. 2003, 67, 1763–1772. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Andersen, M.N.; Jensen, C.R. Carbon retention in the soil–plant system under different irrigation regimes. Agric. Water Manag. 2010, 98, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef]
- Pawlowski, M.N.; Crow, S.E.; Meki, M.N.; Kiniry, J.R.; Taylor, A.D.; Ogoshi, R.; Youkhana, A.; Nakahata, M. Field-Based Estimates of Global Warming Potential in Bioenergy Systems of Hawaii: Crop Choice and Deficit Irrigation. PLoS ONE 2017, 12, e0168510. [Google Scholar] [CrossRef]
- Rochette, P.; Flanagan, L.B. Quantifying rhizosphere respiration in a corn crop under field conditions. Soil Sci. Soc. Am. 1997, 61, 466–474. [Google Scholar] [CrossRef]
- Tu, C.; Li, F.-D.; Qiao, Y.-F.; Zhu, N.; Gu, C.-K.; Zhao, X. Effect of experimental warming on soil respiration under conventional tillage and no-tillage farmland in the North China Plain. J. Integr. Agric. 2017, 16, 967–979. [Google Scholar] [CrossRef]
- de OliveiraSilva, B.; Moitinho, M.R.; de Araújo Santos, G.A.; De Bortoli Teixeira, D.; Fernandes, C.; La Scala, N., Jr. Soil CO2 emission and short-term soil pore class distribution after tillage operations. Soil Tillage Res. 2019, 186, 224–232. [Google Scholar] [CrossRef]
- Dossou-Yovo, E.R.; Brüggemann, N.; Ampofo, E.; Igue, A.M.; Jesse, N.; Huat, J.; Agbossou, E.K. Combining no-tillage, rice straw mulch and nitrogen fertilizer application to increase the soil carbon balance of upland rice field in northern Benin. Soil Tillage Res. 2016, 163, 152–159. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Latitude (N) | Longitude (E) | Elevation (m) | Terrain | Soil Types | Annual Mean Precipitation (mm) | Annual Average Temperature (°C) | Tree-Age | Space (m × m) | Cultivation Modes |
|---|---|---|---|---|---|---|---|---|---|---|
| MZ | 37°47′8″ | 110°5′29″ | 1030 | Platform | Loessi-Orthic Primosols | 451.6 | 8.5 | 10 | 5 × 4 | Standard |
| AS | 36°47′23″ | 109°22′42″ | 1220 | Platform | Loessi-Orthic Primosols | 506.6 | 9.1 | 11 | 5 × 4 | Standard |
| LC | 35°46′2″ | 109°26′28″ | 1160 | Highland | Cumulic Haplustolls | 622 | 9.2 | 16 | 5 × 4 | Standard |
| FP | 34°50′17″ | 109°0′47″ | 570 | Plain | Eum-Orthric Anthrosols | 533.2 | 13.1 | 13 | 4 × 2 | Dwarfing |
| YL | 34°18′28″ | 108°0′15″ | 550 | Plain | Eum-Orthric Anthrosols | 649.5 | 12.9 | 9 | 4 × 1 | Dwarfing |
| Regions | SR (μ mol m−2s−1) | Cumulative Soil Respiration (g C m−2) | Soil Temperature (°C) | Soil Moisture (%) | Soil Organic Carbon (g kg−1) | Fine Root Biomass (g m−2) |
|---|---|---|---|---|---|---|
| MZ | 1.14 (0.38) c | 205 (35) c | 20.49 (4.58) b | 11.46 (4.11) d | 4.18 (0.17) d | 174.5 (29) b |
| AS | 1.33 (0.44) b | 244 (28) bc | 21.00 (4.56) b | 14.23 (4.88) c | 4.98 (0.36) c | 168 (30) b |
| LC | 1.49 (0.47) b | 278 (28) b | 21.76 (4.31) ab | 19.88 (4.33) b | 8.50 (0.50) b | 204.75 (37) b |
| FP | 1.80 (0.46) a | 338 (39) a | 22.99 (3.94) a | 25.99 (4.95) a | 10.75 (0.37) a | 217.5 (40) b |
| YL | 1.96 (0.43) a | 369 (36) a | 23.41 (3.72) a | 27.25 (4.98) a | 11.19 (0.50) a | 224 (25) a |
| Area | Distance (m) | Functions | R2 | p |
|---|---|---|---|---|
| MZ | 0.5 | SR = −0.014 θ2 + 0.422 θ − 1.499 | 0.50 | <0.05 |
| 2 | SR = −0.013 θ2 + 0.309 θ − 0.839 | 0.49 | <0.05 | |
| AS | 0.5 | SR = −0.013 θ2 + 0.428 θ − 1.691 | 0.56 | <0.05 |
| 2 | SR = −0.013 θ2 + 0.406 θ − 1.732 | 0.49 | <0.05 | |
| LC | 0.5 | SR = −0.007 θ2 − 0.233 θ − 0.016 | 0.46 | =0.06 |
| 2 | SR = 0.002 θ2 − 0.170 θ + 3.716 | 0.52 | <0.05 | |
| FP | 0.5 | SR = −0.011 θ2 + 0.499 θ − 3.256 | 0.31 | >0.05 |
| 2 | SR = −0.009 θ2 + 0.437 θ − 3.092 | 0.44 | =0.07 | |
| YL | 0.5 | SR = −0.008 θ2 + 0.377 θ − 1.982 | 0.36 | >0.05 |
| 2 | SR = −0.008 θ2 + 0.413 θ − 3.233 | 0.45 | =0.07 |
| Area | Distance/m | α | β | tα | tβ | R2 | p | Q10 |
|---|---|---|---|---|---|---|---|---|
| MZ | 0.5 | 0.318 ± 0.052 | 0.063 ± 0.008 | −2.419 | 0.489 | 0.82 | <0.05 | 1.88 |
| 2 | 0.411 ± 0.054 | 0.060 ± 0.007 | 0.61 | <0.05 | 1.82 | |||
| AS | 0.5 | 0.408 ± 0.072 | 0.059 ± 0.008 | 0.879 | 0.142 | 0.87 | <0.05 | 1.80 |
| 2 | 0.312 ± 0.175 | 0.057 ± 0.023 | 0.60 | <0.05 | 1.77 | |||
| LC | 0.5 | 0.482 ± 0.144 | 0.056 ± 0.012 | 0.726 | 0.294 | 0.63 | <0.05 | 1.75 |
| 2 | 0.401 ± 0.129 | 0.053 ± 0.013 | 0.62 | <0.05 | 1.70 | |||
| FP | 0.5 | 0.651 ± 0.120 | 0.048 ± 0.008 | 1.230 | 0.120 | 0.68 | <0.05 | 1.62 |
| 2 | 0.533 ± 0.115 | 0.047 ± 0.012 | 0.66 | <0.05 | 1.60 | |||
| YL | 0.5 | 0701 ± 0.147 | 0.047 ± 0.008 | 0.561 | 0.382 | 0.78 | <0.05 | 1.60 |
| 2 | 0.629 ± 0.167 | 0.044 ± 0.011 | 0.66 | <0.05 | 1.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, T.; Wang, Y.; Guo, F.; Jia, Q.; Wu, X.; Wang, E.; Hong, J. Soil Respiration Characteristics and Influencing Factors for Apple Orchards in Different Regions on the Loess Plateau of Shaanxi Province. Sustainability 2021, 13, 4780. https://doi.org/10.3390/su13094780
Hou T, Wang Y, Guo F, Jia Q, Wu X, Wang E, Hong J. Soil Respiration Characteristics and Influencing Factors for Apple Orchards in Different Regions on the Loess Plateau of Shaanxi Province. Sustainability. 2021; 13(9):4780. https://doi.org/10.3390/su13094780
Chicago/Turabian StyleHou, Tingting, Yanping Wang, Fuxing Guo, Qiong Jia, Xinnan Wu, Enguang Wang, and Jingbo Hong. 2021. "Soil Respiration Characteristics and Influencing Factors for Apple Orchards in Different Regions on the Loess Plateau of Shaanxi Province" Sustainability 13, no. 9: 4780. https://doi.org/10.3390/su13094780
APA StyleHou, T., Wang, Y., Guo, F., Jia, Q., Wu, X., Wang, E., & Hong, J. (2021). Soil Respiration Characteristics and Influencing Factors for Apple Orchards in Different Regions on the Loess Plateau of Shaanxi Province. Sustainability, 13(9), 4780. https://doi.org/10.3390/su13094780