

Patterns of Seed Dispersal in Coastal Dune Plant Communities

Abstract
:1. Introduction
2. Materials and Methods
Data Analyses
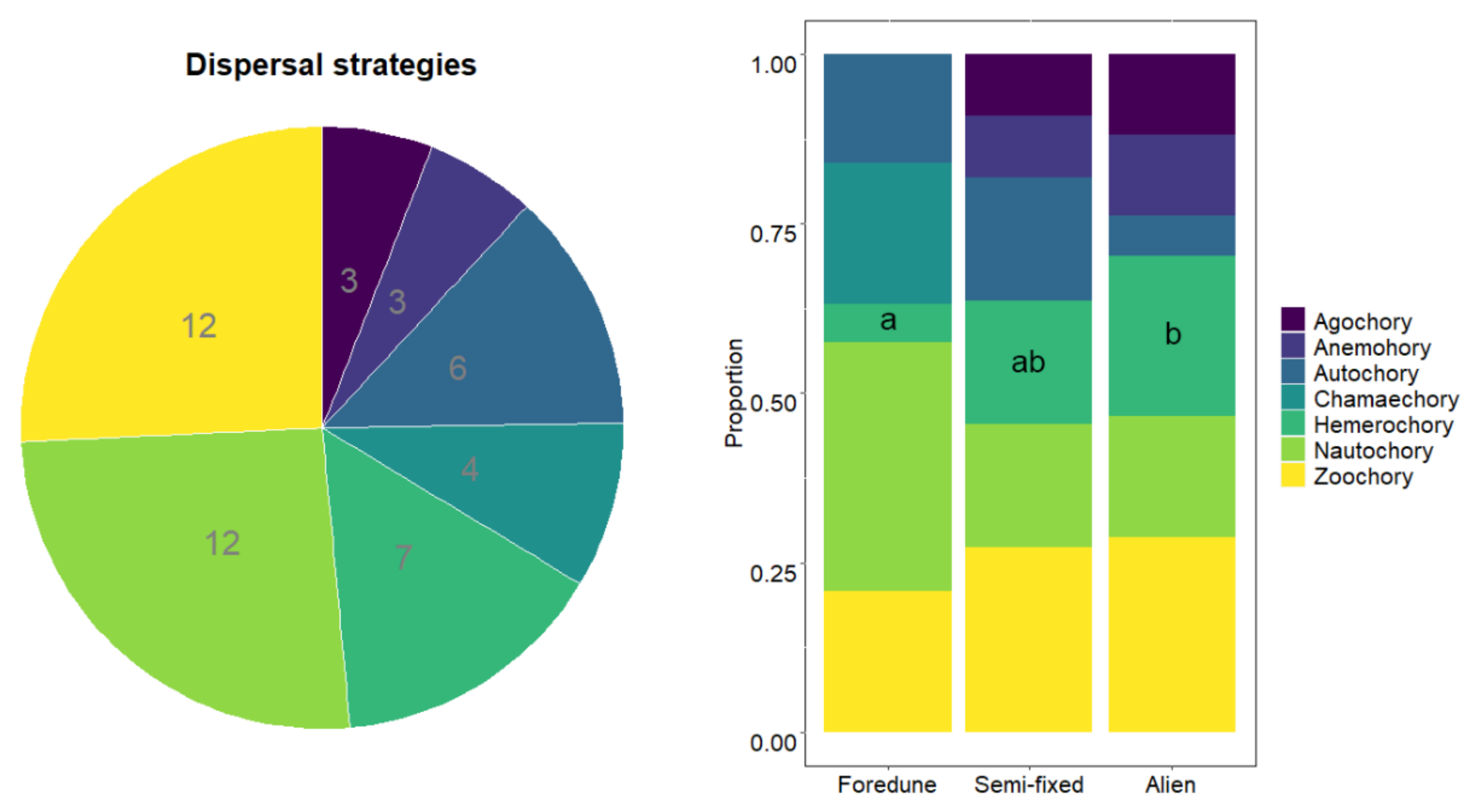
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, D.R. Seed Dispersal; Murray, D.R., Ed.; Academic Press: San Diego, CA, USA, 1986; ISBN 0125119003. [Google Scholar]
- Seale, M.; Nakayama, N. From passive to informed: Mechanical mechanisms of seed dispersal. New Phytol. 2020, 225, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.M.; Lawton, J.H.; Eversham, B.C.; Wood, S.N. The biogeography of scarce vascular plants in Britain with respect to habitat preference, dispersal ability and reproductive biology. Biol. Conserv. 1994, 70, 149–157. [Google Scholar] [CrossRef]
- Eriksson, O.; Jakobsson, A. Recruitment trade-offs and the evolution of dispersal mechanisms in plants. Evol. Ecol. 1999, 13, 411–423. [Google Scholar] [CrossRef]
- Johnson, J.S.; Cantrell, R.S.; Cosner, C.; Hartig, F.; Hastings, A.; Rogers, H.S.; Schupp, E.W.; Shea, K.; Teller, B.J.; Yu, X.; et al. Rapid changes in seed dispersal traits may modify plant responses to global change. AoB Plants 2019, 11, plz020. [Google Scholar] [CrossRef] [PubMed]
- Poschlod, P.; Abedi, M.; Bartelheimer, M.; Drobnik, J.; Rosbakh, S.; Saatkamp, A. Seed ecology and assembly rules in plant communities. Veg. Ecol. Second Ed. 2013, 2, 164–202. [Google Scholar] [CrossRef]
- Van Leeuwen, C.H.A.; Sarneel, J.M.; van Paassen, J.; Rip, W.J.; Bakker, E.S. Hydrology, shore morphology and species traits affect seed dispersal, germination and community assembly in shoreline plant communities. J. Ecol. 2014, 102, 998–1007. [Google Scholar] [CrossRef]
- Cochrane, A.; Yates, C.J.; Hoyle, G.L.; Nicotra, A.B. Will among-population variation in seed traits improve the chance of species persistence under climate change? Glob. Ecol. Biogeogr. 2015, 24, 12–24. [Google Scholar] [CrossRef]
- Guitián, J.; Sánchez, J.M. Seed dispersal spectra of plant communities in the Iberian Peninsula. Vegetatio 1992, 98, 157–164. [Google Scholar] [CrossRef]
- Jara-Guerrero, A.; de la Cruz, M.; Méndez, M. Seed Dispersal Spectrum of Woody Species in South Ecuadorian Dry Forests: Environmental Correlates and the Effect of Considering Species Abundance. Biotropica 2011, 43, 722–730. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological assembly rules in plant communities-approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Rasran, L.; Vogt, K.; Jensen, K. Hydrochorous seed transport in a small river in Northern Germany as trait-dependent filter of plant dispersal and recruitment. Int. Rev. Hydrobiol. 2021, 106, 277–286. [Google Scholar] [CrossRef]
- Nilsson, C.; Brown, R.L.; Jansson, R.; Merritt, D.M. The role of hydrochory in structuring riparian and Wetland vegetation. Biol. Rev. 2010, 85, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Schwab, A.; Stammel, B.; Kiehl, K. Seed dispersal via a new watercourse in a reconnected floodplain: Differences in species groups and seasonality. Restor. Ecol. 2018, 26, S103–S113. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 1998, 9, 113–122. [Google Scholar] [CrossRef]
- Cortés-Flores, J.; Andresen, E.; Cornejo-Tenorio, G.; Ibarra-Manríquez, G. Fruiting phenology of seed dispersal syndromes in a Mexican Neotropical temperate forest. For. Ecol. Manag. 2013, 289, 445–454. [Google Scholar] [CrossRef]
- Pak, D.; Swamy, V.; Alvarez-Loayza, P.; Cornejo, F.; Queenborough, S.A.; Metz, M.R.; Terborgh, J.; Valencia, R.; Wright, S.J.; Garwood, N.C.; et al. Multi-scale phenological niches in diverse Amazonian plant communities. bioRxiv 2021. [Google Scholar] [CrossRef]
- Du, Y.; Mi, X.; Liu, X.; Chen, L.; Ma, K. Seed dispersal phenology and dispersal syndromes in a subtropical broad-leaved forest of China. For. Ecol. Manag. 2009, 258, 1147–1152. [Google Scholar] [CrossRef]
- Escobar, D.F.E.; Silveira, F.A.O.; Morellato, L.P.C. Timing of seed dispersal and seed dormancy in Brazilian savanna: Two solutions to face seasonality. Ann. Bot. 2018, 121, 1197–1209. [Google Scholar] [CrossRef]
- Del Vecchio, S.; Mattana, E.; Ulian, T.; Buffa, G. Functional seed traits and germination patterns predict species coexistence in Northeast Mediterranean foredune communities. Ann. Bot. 2021, 127, 361–370. [Google Scholar] [CrossRef]
- Lõhmus, K.; Balke, T.; Kleyer, M. Spatial and temporal patterns of initial plant establishment in salt marsh communities. J. Veg. Sci. 2020, 31, 1124–1134. [Google Scholar] [CrossRef]
- Beckman, N.G.; Aslan, C.E.; Rogers, H.S.; Kogan, O.; Bronstein, J.L.; Bullock, J.M.; Hartig, F.; HilleRisLambers, J.; Zhou, Y.; Zurell, D.; et al. Advancing an interdisciplinary framework to study seed dispersal ecology. AoB Plants 2020, 12, plz048. [Google Scholar] [CrossRef] [PubMed]
- Beckman, N.G.; Aslan, C.E.; Rogers, H.S. Introduction to the special issue: The role of seed dispersal in plant populations: Perspectives and advances in a changing world. AoB Plants 2020, 12, plaa010. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.; Ercole, S. Gli Habitat Delle Coste Sabbiose Italiane: Ecologia e Problematiche di Conservazione; Acosta, A.T.R., Ercole, S., Eds.; Serie Rapporti; ISPRA: Rome, Italy, 2015; Volume 215, ISBN 9788578110796. [Google Scholar]
- Del Vecchio, S.; Fantinato, E.; Silan, G.; Buffa, G. Trade-offs between sampling effort and data quality in habitat monitoring. Biodivers. Conserv. 2019, 28, 55–73. [Google Scholar] [CrossRef]
- Doing, H. Coastal foredune zonation and succession in various parts of the world. Vegetatio 1985, 61, 65–75. [Google Scholar] [CrossRef]
- Torca, M.; Campos, J.A.; Herrera, M. Changes in plant diversity patterns along dune zonation in south Atlantic European coasts. Estuar. Coast. Shelf Sci. 2019, 218, 39–47. [Google Scholar] [CrossRef]
- Buffa, G.; Gaetan, C.; Piccoli, S.; del Vecchio, S.; Fantinato, E. Using fine-scale field data modelling for planning the management of invasions of Oenothera stucchii in coastal dune systems. Ecol. Indic. 2021, 125, 107564. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Meier, U. (Ed.) Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium: Quedlinburg, Germany, 2018; ISBN 978-3-95547-071-5. [Google Scholar]
- Pignatti, S. Introduzione allo studio fitosociologico della pianura veneta orientale con particolare riguardo alla vegetazione litoranea. Arch. Bot. 1952, 28, 265–329. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Garnier, S.; Ross, N.; Rudis, R.; Camargo, P.A.; Sciaini, M.; Scherer, C. Viridis—Colorblind-Friendly Color Maps for R. 2021. R package version 0.6.1. Available online: https://CRAN.R-project.org/package=viridis (accessed on 1 July 2022).
- Katsnelson, A. Colour me better: Fixing figures for colour blindness. Nature 2021, 598, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Childs, D.Z.; Metcalf, C.J.E.; Rees, M. Evolutionary bet-hedging in the real world: Empirical evidence and challenges revealed by plants. Proc. R. Soc. B Biol. Sci. 2010, 277, 3055–3064. [Google Scholar] [CrossRef]
- Cohen, D. Optimizing Reproduction in a Randomly Varying Environment. J. Theor. Biol. 1966, 12, 119–129. [Google Scholar] [CrossRef]
- Venable, D.L.; Brown, J.S. The selective interactions of dispersal, dormancy, and seed size as adaptations for reducing risk in variable environments. Am. Nat. 1988, 131, 360–384. [Google Scholar] [CrossRef]
- Maun, M. The Biology of Coastal Sandy Dunes; Oxford University Press: Oxford, UK, 2009; ISBN 9788578110796. [Google Scholar]
- McLachlan, A.; Defeo, O. (Eds.) The Ecology of Sandy Shores, 3rd ed.; Academic Press: London, UK, 2017. [Google Scholar]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 2008, 23, 638–647. [Google Scholar] [CrossRef]
- Levin, S.A.; Muller-Landau, H.C.; Nathan, R.; Chave, J. The ecology and evolution of seed dispersal: A theoretical perspective. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 575–604. [Google Scholar] [CrossRef]
- Garrison, W.J.; Miller, G.L.; Raspet, R. Ballistic seed projection in two herbaceous species. Am. J. Bot. 2000, 87, 1257–1264. [Google Scholar] [CrossRef]
- Stamp, N.E.; Lucas, J.R. Ecological correlates of explosive seed dispersal. Oecologia 1983, 59, 272–278. [Google Scholar] [CrossRef]
- Banks, N.C.; Paini, D.R.; Bayliss, K.L.; Hodda, M. The role of global trade and transport network topology in the human-mediated dispersal of alien species. Ecol. Lett. 2015, 18, 188–199. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Kučera, T. Patterns of invasion in temperate nature reserves. Biol. Conserv. 2002, 104, 13–24. [Google Scholar] [CrossRef]
- Yang, Q.; Weigelt, P.; Fristoe, T.S.; Zhang, Z.; Kreft, H.; Stein, A.; Seebens, H.; Dawson, W.; Essl, F.; König, C.; et al. The global loss of floristic uniqueness. Nat. Commun. 2021, 12, 7290. [Google Scholar] [CrossRef] [PubMed]
- Fenner, M. The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, NY, USA, 2000; ISBN 0851994326. [Google Scholar]
- Obseso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L. The physiology of invasive plants in low-resource environments. Conserv. Physiol. 2013, 1, cot026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; ISBN 9783540773405. [Google Scholar]



{kind=link}
{kind=link}
| Dispersal Strategies | Vector | Description/Seed Morphology |
|---|---|---|
| Agochory | Human activities | The vector of transmission, in this case, is human activities (e.g., agriculture). Seeds of species adopting this strategy exploit anthropogenic means such as operating and/or agricultural machinery, truckloads, or ships |
| Anemochory | Wind | Plants produce light seeds, often with thin expansions to favor dispersal by wind |
| Autochory | Autonomous dispersal (no vector) | Plants simply drop seeds on the ground or hurl them through explosive mechanisms (in this case, the strategy is called “bolochory”). Autochory has the advantage of being independent of biotic or abiotic vectors, although the extent of dispersal is not very far from the mother plant |
| Chamaechory | Wind | When the individual reaches senescence, the aerial portion detaches itself from the roots and rolls away. The plant tends to have a rounded shape, which allows it to roll for considerable distances |
| Hemerochory | Humans and domestic animals | In this case, the transmission vector is humans. This strategy is similar to zoochory and allows seeds to attach themselves to shoes and clothes or domestic animals in order to be transported; this strategy is also long-range, as humans also move hundreds of kilometers |
| Nautochory | Water | It occurs through water (e.g., marine currents, run-offs, rivers, and canals). The fruits produced have to be able to float and survive in water for a long time without damaging the seeds |
| Zoochory | Animals | It occurs with the help of the most diverse species, such as birds, insects, worms, and mammals, above all, when the seeds remain attached to their hairy bodies. Fruits can be eaten by animals as well; in this case, they ought to be fragrant and attractive |
| Species | Number of Strategies | Dispersal Strategy | Evenness Index | |
|---|---|---|---|---|
| Foredune foundation species | Cakile maritima subsp. maritima | 2 | Chamaechory; Nautochory | 0.57 |
| Calamagrostis arenaria subsp. arundinacea | 2 | Nautochory; Zoochory | 0.55 | |
| Calystegia soldanella | 1 | Nautochory | 0.28 | |
| Echinophora spinosa | 1 | Chamaechory | 0.09 | |
| Elymus farctus | 2 | Nautochory; Zoochory | 0.55 | |
| Eryngium maritimum | 5 | Autochory; Chamaechory; Hemerochory; Nautochory; Zoochory | 0.31 | |
| Euphorbia paralias | 2 | Autochory; Nautochory | 0.54 | |
| Euphorbia peplis | 1 | Autochory | 0.34 | |
| Salsola tragus | 3 | Chamaechory; Nautochory; Zoochory | 0.16 | |
| Semi-fixed dune | Festuca fasciculata | 3 | Anemochory; Nautochory; Zoochory | 0.44 |
| Cyperus capitatus | 1 | Autochory | 0.44 | |
| Hypochaeris radicata | 4 | Agochory; Hemerochory; Nautochory; Zoochory | 0.39 | |
| Medicago littoralis | 2 | Hemerochory; Zoochory | 0.18 | |
| Silene colorata | 1 | Autochory | 0.37 | |
| Alien | Ambrosia psilostachya | 2 | Hemerochory; Zoochory | 0.28 |
| Cenchrus longispinus | 2 | Hemerochory; Zoochory | 0.46 | |
| Erigeron canadensis | 5 | Agochory; Anemochory; Hemerochory; Nautochory; Zoochory | 0.39 | |
| Oenothera stucchii | 6 | Agochory; Anemochory; Autochory; Hemerochory; Nautochory; Zoochory | 0.47 | |
| Xanthium orientale | 2 | Nautochory; Zoochory | 0.69 |
| Df | Deviance | Resid. Df | Resid. Dev | Pr(>Chi) | |
|---|---|---|---|---|---|
| NULL | 227 | 4043.6 | |||
| Month | 11 | 1301.34 | 216 | 2742.3 | <0.0001 |
| Species group | 2 | 149.15 | 214 | 2593.2 | <0.0001 |
| Month × Species group | 22 | 1217.79 | 192 | 1375.4 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdesolo, T.; Del Vecchio, S.; Buffa, G. Patterns of Seed Dispersal in Coastal Dune Plant Communities. Sustainability 2022, 14, 10983. https://doi.org/10.3390/su141710983
Valdesolo T, Del Vecchio S, Buffa G. Patterns of Seed Dispersal in Coastal Dune Plant Communities. Sustainability. 2022; 14(17):10983. https://doi.org/10.3390/su141710983
Chicago/Turabian StyleValdesolo, Tommaso, Silvia Del Vecchio, and Gabriella Buffa. 2022. "Patterns of Seed Dispersal in Coastal Dune Plant Communities" Sustainability 14, no. 17: 10983. https://doi.org/10.3390/su141710983
APA StyleValdesolo, T., Del Vecchio, S., & Buffa, G. (2022). Patterns of Seed Dispersal in Coastal Dune Plant Communities. Sustainability, 14(17), 10983. https://doi.org/10.3390/su141710983