

Alfalfa Cover Crops Influence the Soil Fungal Community and Function in Apple Orchards in Arid Desert Oases in Northwest China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling Collection
2.2. Collection and Treatment of Soil Samples
2.3. Determination of Soil Chemical Analysis
2.4. Fungal DNA Extraction and NGS
2.5. Data Processing and Analysis
3. Results
3.1. Diversity and Richness of the Soil Fungal Community
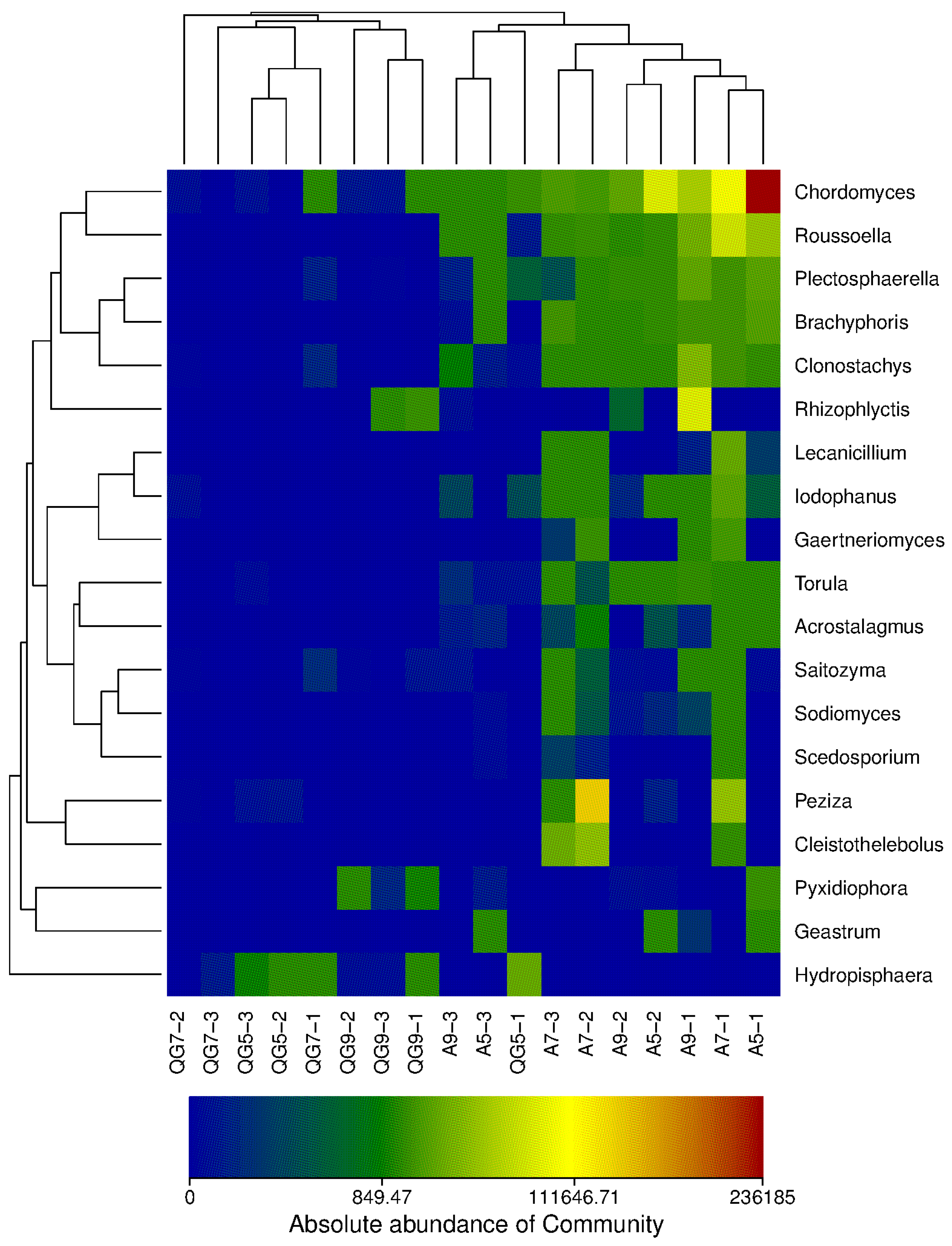
3.2. Distribution of the Soil Fungal Community
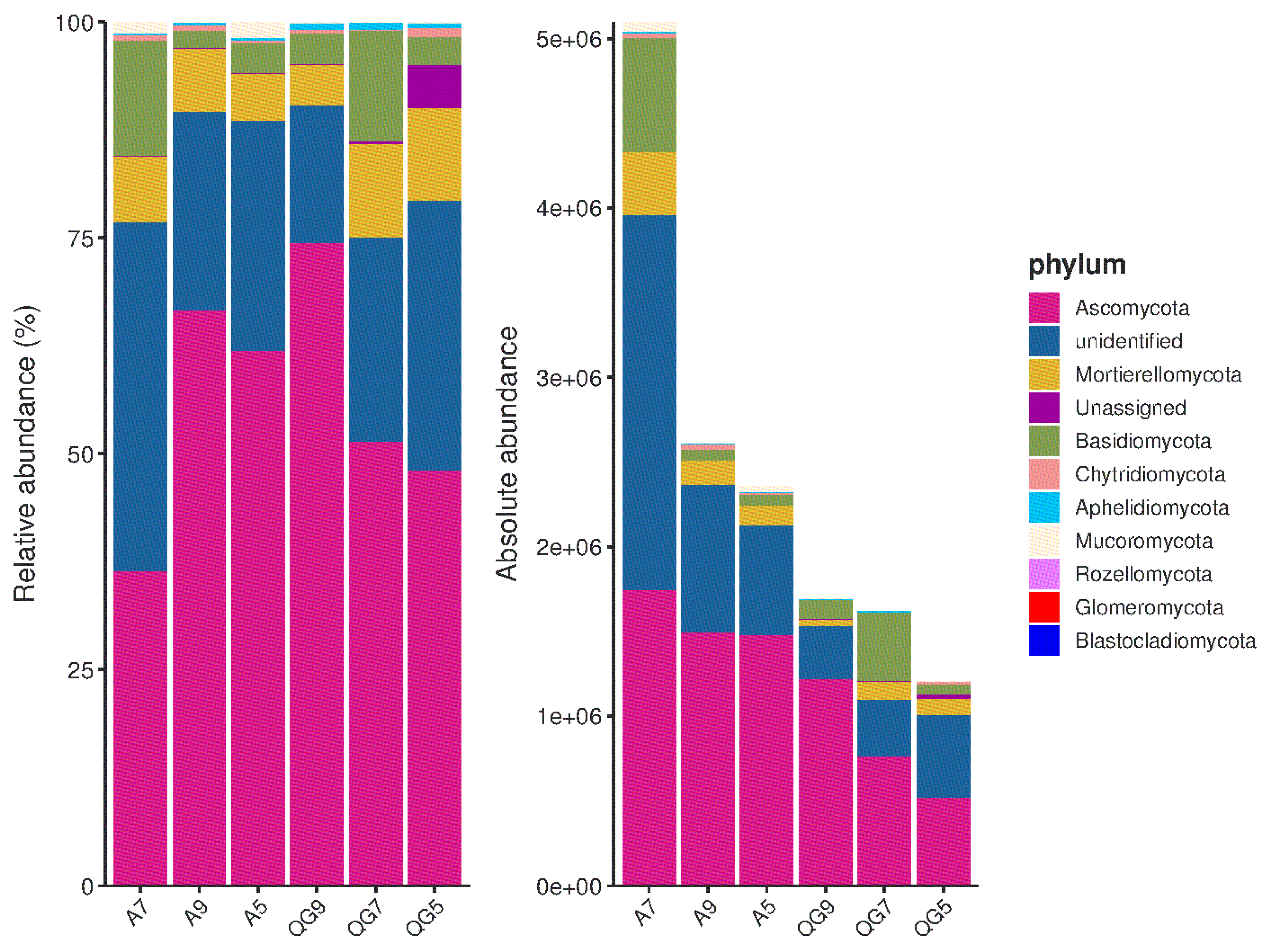
3.3. Composition and Structure of the Fungal Community
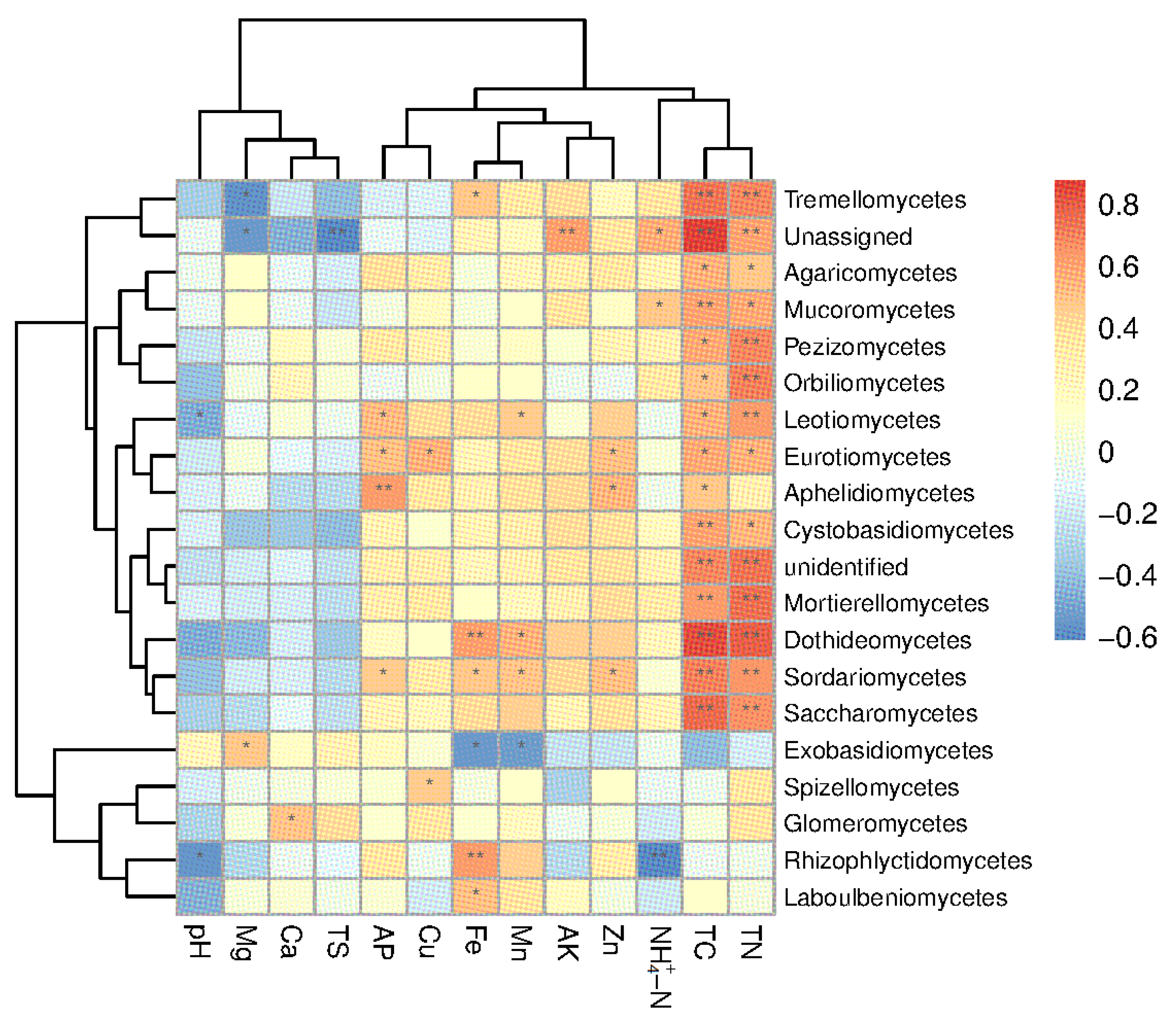
3.4. Correlation between Soil Fungi and Soil Factors
3.5. Functional Groups of Soil Fungi
4. Discussion
4.1. Impact of Cover Crops on Microbial Diversity
4.2. Impact of Cover Crops on Fungal Community Structure
4.3. Impact of Cover Crops on Fungal Function
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, W.H.; Yang, L. Xinjiang Statistical Yearbook; China Statistics Press: Urumqi, China, 2018; p. 358. [Google Scholar]
- Daryanto, S.; Fu, B.; Wang, L.; Jacinthe, P.-A.; Zhao, W. Quantitative synthesis on the ecosystem services of cover crops. Earth-Sci. Rev. 2018, 185, 357–373. [Google Scholar] [CrossRef]
- Du, S.; Bai, G.; Yu, J. Soil properties and apricot growth under intercropping and mulching with erect milk vetch in the loess hilly-gully region. Plant Soil 2015, 390, 431–442. [Google Scholar] [CrossRef]
- Yao, S.R.; Merwin, I.A.; Bird, G.W.; Abawi, G.S.; Thies, J.E. Orchard floor management practices that maintain vegetative or biomass groundcover stimulate soil microbial activity and alter soil microbial community composition. Plant Soil 2005, 271, 377–389. [Google Scholar] [CrossRef]
- Blesh, J. Functional traits in cover crop mixtures: Biological nitrogen fixation and multifunctionality. J. Appl. Ecol. 2018, 55, 38–48. [Google Scholar] [CrossRef]
- Bénédicte, A.; Bruno, M.; Claire, C.; May, B.; Cyril, G.; Michel, B.; Gilles, G.; Nicolas, B. Alternative arable cropping systems: A key to increase soil organic carbon storage, Results from a 16 year field experiment. Agric. Ecosyst. Environ. 2016, 232, 150–164. [Google Scholar]
- Prosdocimi, M.; Tarolli, P.; Cerdà, A. Mulching practices for reducing soil water erosion. Earth-Sci. Rev. 2016, 161, 191–203. [Google Scholar] [CrossRef]
- Reeve, J.; Culumber, C.; Black, B.; Tebeau, A.; Ransom, C.; Alston, D.; Rowley, M.; Lindstrom, T. Establishing peach trees for organic production in Utah and the Intermountain West. Sci. Hortic. 2017, 214, 242–251. [Google Scholar] [CrossRef]
- Pignataro, A.; Moscatelli, M.C.; Mocali, S.; Grego, S.; Benedetti, A. Assessment of soil microbial functional diversity in a coppiced forest system. Appl. Soil Ecol. 2012, 62, 115–123. [Google Scholar] [CrossRef]
- Srivastava, A.; Huchche, A.; Ram, L.; Singh, S. Yield prediction in intercropped versus monocropped citrus orchards. Sci. Hortic. 2007, 114, 67–70. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, W.; Wang, R.; Wang, W.; Shangguan, Z. Decreased occurrence of carbon cycle functions in microbial communities along with long-term secondary succession. Soil Biol. Biochem. 2018, 123, 207–217. [Google Scholar] [CrossRef]
- Gabhane, J.; William, S.P.; Bidyadhar, R.; Bhilawe, P.; Anand, D.; Vaidya, A.N.; Wate, S.R. Additives aided composting of green waste: Effects on organic matter degradation, compost maturity, and quality of the finished compost. Bioresour. Technol. 2012, 114, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Duchene, O.; Vian, J.F.; Celette, F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Ramos, M.E.; Robles, A.B.; Navarro, A.S.; González-Rebollar, J.L. Soil responses to different management practices in rainfed orchards in semiarid environments. Soil Tillage Res. 2011, 112, 85–91. [Google Scholar] [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.D. Analysis of Soil Agrochemical, 3rd ed.; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Lei, H.J.; Xiao, Z.Y.; Zhang, Z.H.; Jin, C.C.; Pan, H.W.; Sun, K.P.; Xi, H.P. Effects of Water-nitrogen coupled oxygen Irrigation on soil Fertility and bacterial Community of greenhouse Pepper. Trans. Chin. Soc. Agric. Eng. 2021, 37. [Google Scholar]
- Spalding, E.P.; Miller, N.D. Image analysis is driving a renaissance in growth measurement. Curr. Opin. Plant Biol. 2013, 16, 100–104. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Jiang, S.-Q.; Yu, Y.-N.; Gao, R.-W.; Wang, H.; Zhang, J.; Li, R.; Long, X.-H.; Shen, Q.-R.; Chen, W.; Cai, F. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial community in a tomato cultivated coastal saline soil. Sci. Total Environ. 2019, 687, 601–609. [Google Scholar] [CrossRef]
- Dang, A.T.N.; Nandy, S.; Srinet, R.; Luong, N.V.; Ghosh, S.; Kumar, A.S. Forest aboveground biomass estimation using machine learning regression algorithm in Yok Don National Park, Vietnam. Ecol. Inform. 2019, 50, 24–32. [Google Scholar] [CrossRef]
- Yuan, S.; Zhang, N.; Wu, X.; Qian, Y.; Chen, X.; Raza, W.; Li, X. Effect of pruned material, extracts, and polyphenols of tea on enzyme activities and microbial community structure in soil. Soil Sci. Plant Nutr. 2017, 63, 607–615. [Google Scholar] [CrossRef]
- Elfstrand, S.; Bath, B.; Martensson, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Appl. Soil Ecol. 2007, 36, 70–82. [Google Scholar] [CrossRef]
- Caban, J.R.; Kuppusamy, S.; Kim, J.H.; Yoon, Y.-E.; Kim, S.Y.; Lee, Y.B. Green manure amendment enhances microbial activity and diversity in antibiotic-contaminated soil. Appl. Soil Ecol. 2018, 129, 72–76. [Google Scholar] [CrossRef]
- Liu, Y.P.; Mao, Y.F.; Hu, Y.L.; Zhang, L.L.; Yin, Y.J.; Pang, H.L.; Su, X.F.; Yang, L.; Shen, X. Effects of grass planting in apple orchard on soil microbial diversity, enzyme activities and carbon components. J. Plant Nutr. Fertil. 2021, 10, 1792–1805. [Google Scholar]
- Si, G.H.; Zhao, S.J.; Wang, R.; Xu, D.B.; Qin, X.C.; Tan, J.; Xiang, B.K.; Yuan, J.F. Effects of consecutive overturning of green manure on soil physical and biological characteristics in tobacco-planting fields. J. Plant Nutr. Fertil. 2014, 20, 905–912. [Google Scholar]
- Wei, Q.Q.; Han, M.Y.; Mu, X.Q. Influences of white clover returning to apple orchard on soil microbial community. Pratacultural Sci. 2016, 33, 385–392. [Google Scholar]
- Fu, H.P.; Zhou, P.Q.; Wang, Y.J.; Mo, Z.D.; Li, Z.H.; Ma, Q.P.; Hoang, H.N.; Li, X.H. Effects of intercropping different green manures on fungal community characteristics in rhizosphere soil of tea plant. J. Tea Commun. 2020, 47, 406–415. [Google Scholar]
- Qian, Y.L.; Wang, X.Z.; Lai, X.F. Effects of perennial forage planting on soil fungal community characteristics in apple orchard. J. Grass Ind. 2019, 28, 124. [Google Scholar]
- Glass, N.L.; Schmoll, M.; Cate, J.H.; Coradetti, S. Plant cell wall deconstruction by ascomycete fungi. Annu. Rev. Microbiol. 2013, 67, 477–498. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Chang, S.X.; Liang, X.; Qin, H.; Chen, J.; Xu, Q. Linking soil fungal community structure and function to soil organic carbon chemical composition in intensively managed subtropical bamboo forests. Soil Biol. Biochem. 2017, 107, 19–31. [Google Scholar] [CrossRef]
- Chen, L.L.; Liu, J.; Li, M.D.; Zhang, L.Z.; Zou, Y.B.; Huang, M. Diversity of Filamentous Fungus Community in Paddy Fields with Different Tillage Methods. Acta Laser Biol. Sin. 2018, 27, 60–68. [Google Scholar]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Yan, H.-T.; Yin, Q.-Y.; Ding, S.-S.; Ren, T.-B.; Xu, J.-L.; Zong, H.; Gao, Q.; Liu, G.-S. Effect of biochar amendment on physicochemical properties and fungal community structures of cinnamon soil. Environ. Sci. 2018, 39, 2412–2419. [Google Scholar]
- Jiang, Y.; Arafat, Y.; Letuma, P.; Ali, L.; Tayyab, M.; Waqas, M.; Li, Y.; Lin, W.; Lin, S.; Lin, W. Restoration of long-term monoculture degraded tea orchard by green and goat manures applications system. Sustainability 2019, 11, 1011. [Google Scholar] [CrossRef]
- Wang, J.; Rhodes, G.; Huang, Q.; Shen, Q. Plant growth stages and fertilization regimes drive soil fungal community compositions in a wheat-rice rotation system. Biol. Fertil. Soils 2018, 54, 731–742. [Google Scholar] [CrossRef]
- Anthony, M.A.; Frey, S.D.; Stinson, K.A. Fungal community homogenization, shift in dominant trophic guild, and appearance of novel taxa with biotic invasion. Ecosphere 2017, 8, e01951. [Google Scholar] [CrossRef]
- Sanon, A.; Beguiristain, T.; Cébron, A.; Berthelin, J.; Duponnois, R. Towards the influence of plant-soil-microbes feedbacks on plant biodiversity, grassland variability and productivity. In Grassland Biodiversity: Habitat Types, Ecological Processes and Environmental Impacts; Runas, J., Dahlgren, T., Eds.; Nova Science Publishers: New York, NY, USA, 2011; pp. 267–301. [Google Scholar]
- Li, T.; Hu, Y.; Hao, Z.; Li, H.; Wang, Y.; Chen, B. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef]
- De, L.; Pena, E.; Echeverría, S.R.; Van, D.; Putten, W.H.; Freitas, H.; Moens, M. Mechanism of control of root-feeding nematodes by mycorrhizal fungi in the dune grass Ammophila arenaria. New Phytol. 2006, 169, 829–840. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | QG5 | A5 | QG7 | A7 | QG9 | A9 |
|---|---|---|---|---|---|---|
| TC (gkg−1) | 2.97 ± 0.31 a | 3.1 ± 0.3 a | 2.72 ± 0.4 a | 2.9 ± 0.39 a | 2.78 ± 0.24 a | 2.88 ± 0.25 a |
| TN (gkg−1) | 0.85 ± 0.06 ab | 0.91 ± 0.07 ab | 0.8 ± 0.05 b | 0.91 ± 0.04 a | 0.81 ± 0.06 ab | 0.91 ± 0.07 ab |
| NH4+-N (mgkg−1) | 38.92 ± 9.82 a | 30.33 ± 6.92 a | 4.78 ± 4.77 b | 5.4 ± 2.4 b | 1.15 ± 0.19 b | 1.58 ± 0.7 b |
| AP (mgkg−1) | 9.91 ± 3.97 b | 14.57 ± 2.32 ab | 24.2 ± 7.87 a | 20.65 ± 3.58 ab | 23.72 ± 2.49 a | 21.95 ± 0.59 a |
| AK (mgkg−1) | 136.77 ± 38.5 a | 155.03 ± 42.8 a | 123.67 ± 6.28 a | 99.4 ± 46.35 a | 120.27 ± 45.91 a | 83.7 ± 22.4 a |
| Cu (mgkg−1) | 21.59 ± 0.78 c | 21.47 ± 0.57 c | 23.32 ± 0.99 ab | 23.39 ± 0.93 a | 22.74 ± 0.31 abc | 21.81 ± 1.26 bc |
| Zn (mgkg−1) | 72.36 ± 1.29 a | 69.69 ± 2.05 a | 74.45 ± 4.78 a | 72.34 ± 2.72 a | 74.68 ± 1.06 a | 73.14 ± 3.12 a |
| Fe (mgkg−1) | 25.9 ± 0.56 a | 25.98 ± 0.36 a | 25.29 ± 0.77 a | 25.84 ± 0.95 a | 27.01 ± 0.04 a | 26.74 ± 0.71 a |
| Mn (mgkg−1) | 0.58 ± 0.02 a | 0.58 ± 0.01 a | 0.58 ± 0.02 a | 0.59 ± 0.02 a | 0.61 ± 0.02 a | 0.59 ± 0.02 a |
| Ca (mgkg−1) | 0.56 ± 0.29 b | 2.57 ± 0.56 a | 2.1 ± 0.27 a | 2.61 ± 0.46 a | 1.75 ± 0.46 a | 2.5 ± 1.13 a |
| Mg (mgkg−1) | 0.05 ± 0.02 d | 0.13 ± 0.02 ab | 0.17 ± 0.03 a | 0.14 ± 0.04 ab | 0.12 ± 0.04 bc | 0.07 ± 0.01 cd |
| pH | 8.18 ± 0.08 a | 7.91 ± 0.05 c | 8.03 ± 0.01 b | 7.94 ± 0.05 c | 7.91 ± 0.05 c | 7.81 ± 0.02 d |
| TS(%) | 0.35 ± 0.15 b | 0.86 ± 0.18 a | 0.86 ± 0.18 a | 0.93 ± 0.16 a | 0.67 ± 0.19 ab | 0.86 ± 0.33 a |
| OTUs | Chao1 | ACE | Shannon | Simpson | Coverage | |
|---|---|---|---|---|---|---|
| QG5 | 376 ± 92 a | 391 ± 79 a | 391 ± 79 a | 4.0708 ± 0.2631 a | 0.0534 ± 0.0270 b | 1.00 ± 0.0 a |
| A5 | 367 ± 85 a | 383 ± 64 a | 383 ± 64 a | 3.9907 ± 0.1239 a | 0.0504 ± 0.0078 b | 1.00 ± 0.0 a |
| QG7 | 363 ± 106 a | 370 ± 103 a | 370 ± 103 a | 4.0163 ± 0.3887 a | 0.0544 ± 0.0409 b | 1.00 ± 0.0 a |
| A7 | 390 ± 71 a | 390 ± 71 a | 390 ± 71 a | 3.4343 ± 0.6231 b | 0.1056 ± 0.0636 a | 1.00 ± 0.0 a |
| QG9 | 346 ± 4 a | 359 ± 7 a | 359 ± 7 a | 3.8754 ± 0.3665 a | 0.0478 ± 0.0132 b | 1.00 ± 0.0 a |
| A9 | 420 ± 45 a | 424 ± 45 a | 424 ± 45 a | 3.7333 ± 0.3552 ab | 0.0831 ± 0.0436 ab | 1.00 ± 0.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhao, W.; Xi, L. Alfalfa Cover Crops Influence the Soil Fungal Community and Function in Apple Orchards in Arid Desert Oases in Northwest China. Sustainability 2022, 14, 11816. https://doi.org/10.3390/su141911816
Wang Z, Zhao W, Xi L. Alfalfa Cover Crops Influence the Soil Fungal Community and Function in Apple Orchards in Arid Desert Oases in Northwest China. Sustainability. 2022; 14(19):11816. https://doi.org/10.3390/su141911816
Chicago/Turabian StyleWang, Zhenlei, Weiliang Zhao, and Linqiao Xi. 2022. "Alfalfa Cover Crops Influence the Soil Fungal Community and Function in Apple Orchards in Arid Desert Oases in Northwest China" Sustainability 14, no. 19: 11816. https://doi.org/10.3390/su141911816
APA StyleWang, Z., Zhao, W., & Xi, L. (2022). Alfalfa Cover Crops Influence the Soil Fungal Community and Function in Apple Orchards in Arid Desert Oases in Northwest China. Sustainability, 14(19), 11816. https://doi.org/10.3390/su141911816