
Food and Feeding Biology of Nile Tilapia (Oreochromis niloticus) in Lake Langeno, Ethiopia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Study Design, Sample Site Selection and Fish Sampling Method
2.3. Fish Stomach Collection Method
2.4. Stomach Content Analysis
2.5. Statistical Analysis
3. Results
3.1. The Status of Collected Stomach Samples
3.2. Diet Composition
3.3. Variation of Food Composition with the Study Sites
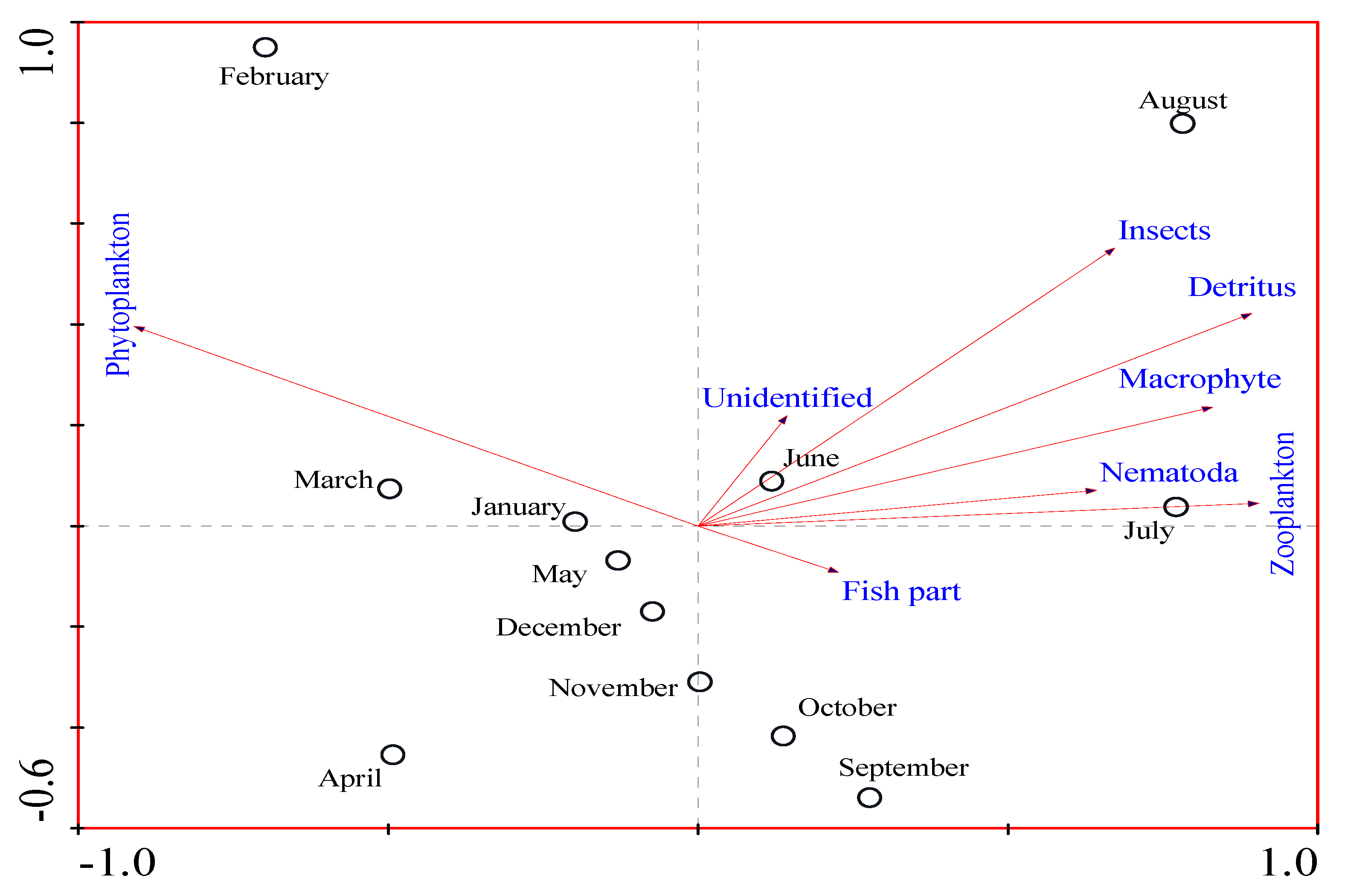
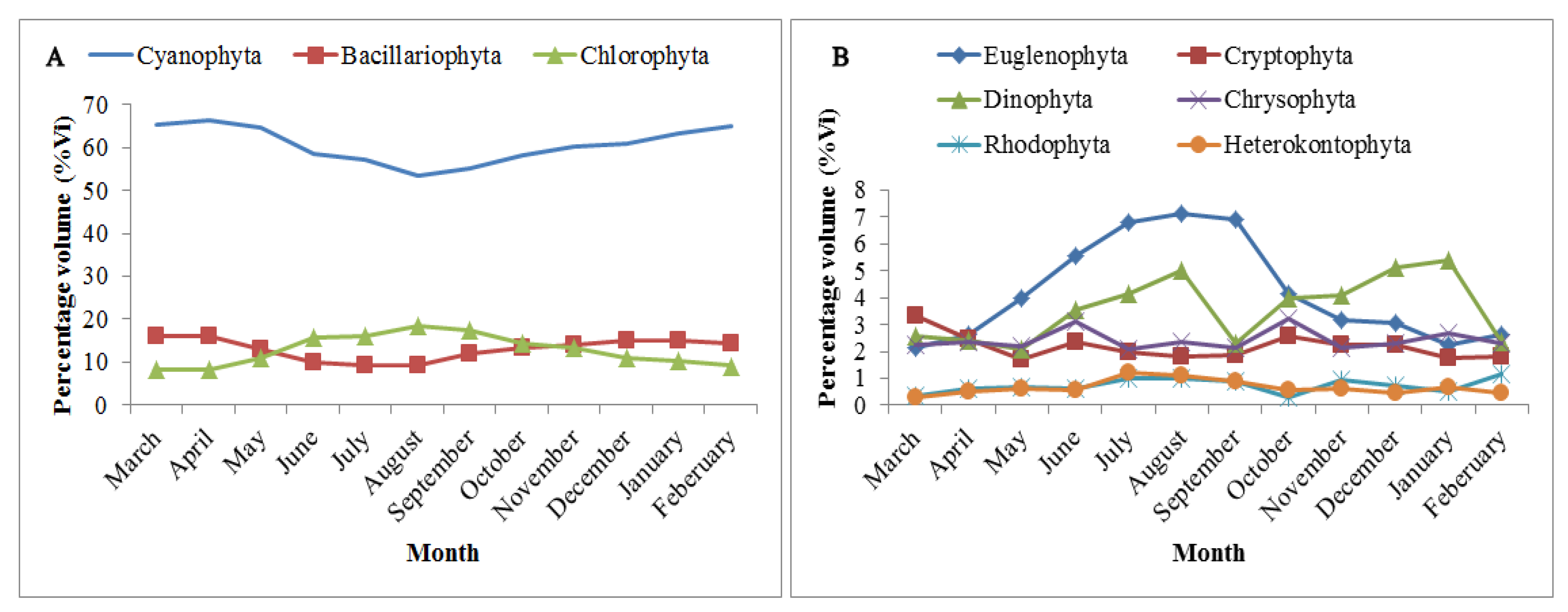
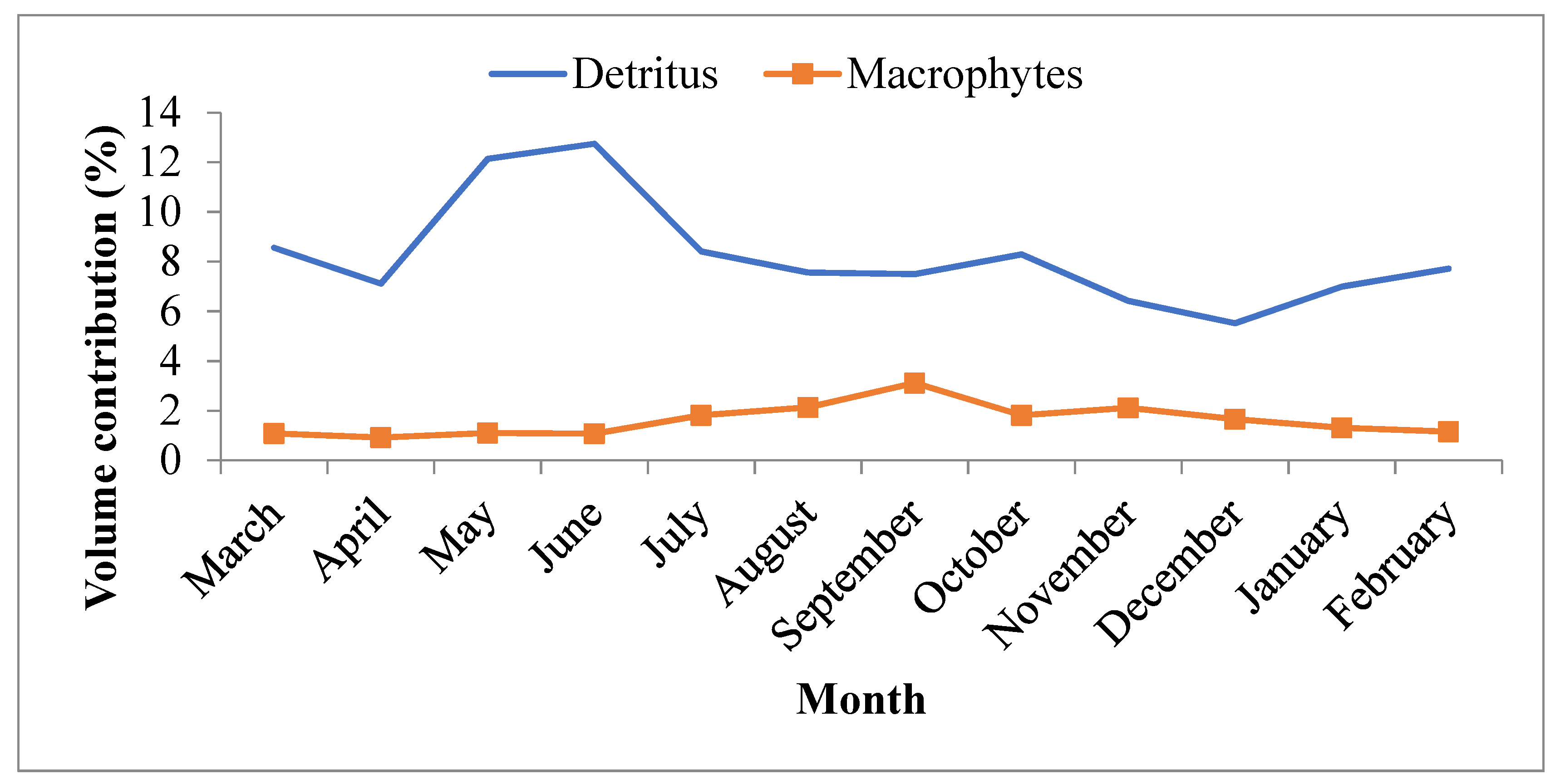
3.4. Seasonal Variation in the Food Composition of O. niloticus in Lake Langeno
3.5. Variation of Food Composition with Fish Size
4. Discussion
4.1. Diet Composition
4.2. Seasonal Variation in the Diet of O. niloticus in Lake Langeno
4.3. Variation of Food Compositions with Fish Sizes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wakil, U.; Haruna, A.; Mohammed, G.; Ndirmbita, W.; Yachilla, B.; Kumai, M. Examinations of the stomach contents of two fish species (C. gariepinus and O. niloticus) in Lake Alau, North-Eastern Nigeria. Agric. For. Fish. 2014, 3, 405–409. [Google Scholar]
- Keyombe, J.L.; Waithaka, E.; Obegi, B. Length–weight relationship and condition factor of Clarias gariepinus in Lake Naivasha, Kenya. Int. J. Fish. Aquat. Stud. 2015, 2, 382–385. [Google Scholar]
- Bolarinwa, J.; Popoola, B. Length-Weight Relationships of Some Economic Fishes of Ibeshe Waterside, Lagos Lagoon, Nigeria. Aquat. Res. Dev. 2015, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Houlihan, D.; Boujard, T.; Jobling, M. Food Intake in Fish; Blackwell Science: Oxford, UK, 2001. [Google Scholar]
- Otieno, O.N.; Kitaka, N.; Njiru, J.M. Length-weight relationship, condition factor, length at first maturity and sex ratio of Nile tilapia, O. niloticus in Lake Naivasha, Kenya. Int. J. Fish. Aquat. Stud. 2014, 2, 67–72. [Google Scholar]
- Shalloof, K.A.S.; Khalifa, N. Stomach Contents and Feeding Habits of Oreochromis niloticus (L.) From Abu-Zabal Lakes, Egypt. World Appl. Sci. J. 2009, 6, 1–5. [Google Scholar]
- Amal, M.N.A.; Zamri-Saad, M. Streptococcosis in Tilapia (Oreochromis niloticus): A Review. Pertanika J. Trop. Agric. Sci. 2011, 34, 195–206. [Google Scholar]
- Mohammed, E.Y.; Uraguchi, Z.B. Impacts of climate change on fisheries: Implications for food security in Sub-Saharan Africa. In Global Food Security; Hanjra, M.A., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2013; pp. 114–135. [Google Scholar]
- Adeyemi, S.O.; Bankole, N.O.; Adikwu, I.A.; Akombu, P.M. Food and feeding habit Habits of some commercially important fish species in Gbedikere Lake, BassaKogi State, Nigeria. Int. J. Lakes Rivers 2009, 2, 31–36. [Google Scholar]
- Canonico, G.C.; Arthington, A.; Thieme, M.L. The effects of introduced tilapias on native biodiversity. Aquat. Conserv. Mar. Freshw. Ecol. 2005, 15, 463–483. [Google Scholar] [CrossRef]
- Mitike, A. Fish Production, Consumption and Management in Ethiopia. Res. J. Agric. Environ. Manag. 2014, 3, 460–466. [Google Scholar]
- Tesfaye, G.; Wolff, M. The state of inland fisheries in Ethiopia: A synopsis with updated estimates of potential yield. Ecohydrol. Hydrobiol. 2014, 14, 200–219. [Google Scholar] [CrossRef]
- Tadesse, Z. The nutritional status and digestibility of Oreochromis niloticus L. diet in Lake Langeno, Ethiopia. Hydrobiologia 1999, 416, 97–106. [Google Scholar] [CrossRef]
- Engdaw, F.; Dadebo, E.; Nagappan, R. Morphometric relationships and feeding habits of Nile Tilapia Oreochromis niloticus (L.) (Pisces: Cichlidae) from Lake Koka, Ethiopia. Int. J. Fish. Aquat. Sci. 2013, 2, 65–71. [Google Scholar]
- Worie, W.; Getahun, A. The food and feeding ecology of Nile tilapia, O. niloticus, in Lake Hayq, Ethiopia. J. Fish. Aquat. Stud. 2015, 2, 176–185. [Google Scholar]
- Temesgen, M.; Getahun, A.; Lemma, B. Diversity, distribution and abundance of fish species in Lake Langeno, Ethiopia. Ethiop. J. Biol. Sci. 2016, 15, 1009–1140. [Google Scholar]
- Temesgen, M. Fish Biology and Fishery Management of Commercial Stocks in a Tropical Rift Valley Lake, Lake Langeno, Ethiopia. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2018. [Google Scholar]
- Abera, L. Current Status and Trends of Fishes and Fishery of a Shallow Rift Valley Lake, Lake Zeway, Ethiopia. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2016. [Google Scholar]
- Tesfaye, G.; Tadesse, Z. Length-weight relationship, Fulton’s condition factor and size at first maturity of Tilapia, Oreochromis niloticus in Lakes Koka, Zeway and Langeno (Ethiopian rift valley). Ethiop. J. Biol. Sci. 2008, 2, 139–157. [Google Scholar]
- Abera, L. Breeding season and condition factor of Oreochromis niloticus (Pisces: Cichlidae) in Lake Babogaya, Ethiopia. Int. J. Agric. Sci. 2012, 7, 116–120. [Google Scholar]
- Teferi, Y.; Admasu, D.; Mengistou, S. The food and feeding habit of O. niloticus L. in Lake Chamo, Ethiopia. SENIT Ethiop. J. Sci. 2000, 23, 1–12. [Google Scholar]
- Ayenew, T. Environmental implications of changes in the levels of lakes in the Ethiopian Rift since 1970. Reg. Environ. Chang. 2004, 4, 192–204. [Google Scholar] [CrossRef]
- Kebede, E. Phytoplankton in a Salinity-Alkalinity Series of Lakes in the Ethiopian Rift Valley. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 1996. [Google Scholar]
- Wodajo, K.; Belay, A. Species composition and seasonal abundance of zooplankton in two Ethiopian Rift Valley Lakes-Lake Abijata and Langeno. Hydrobiology 1984, 113, 129–136. [Google Scholar] [CrossRef]
- Imam, T.S.; Bala, U.; Balarabe, M.L.; Oyeyi, T.I. Length-weight relationship and condition factor of four fish species from Wasai Reservoir in Kano, Nigeria. Afr. J. General Agric. 2010, 6, 125–130. [Google Scholar]
- Gill, K. Identification Guide to Freshwater Macro-Invertebrates; Stroud Water Research Center Macro-Invertebrate Images Prepared for Stroud Water Research Center: Avondale, PA, USA, 1998. [Google Scholar]
- Harding, J.P.; Smith, W.A. A Key to the British Freshwater Cyclopid and Calanoid Copepods; Freshwater Biological Association Scientific Publication No. 18; Titus Wilson and Son Ltd.: London, UK, 1974. [Google Scholar]
- Baker, P.D.; Fabbro, L.D. A Guide to the Identification of Common Blue-Green Algae in Australian Freshwaters. Cooperative Research Center for Freshwater Ecology Identification Guide, 2nd ed.; National Library of Australia Cataloguing in Publication: Perth, Australia, 2002. [Google Scholar]
- Carling, K.J.; Ater, I.M.; Pellam, M.R.; Bouchard, A.M.; Mihuc, T.B. A Guide to the Zooplankton of Lake Champlain. Plattsburgh State Univ. N. Y. 2004, 1, 33–66. [Google Scholar]
- Vuuren, S.J.; Taylor, J.; Ginkel, C.; Gerber, A. Easy Identification of the Most Common FRESHWATER ALGAE; A Guide for the Identification of Microscopic Algae in South African Freshwaters; Department of Water and Forestry, North-West University: Pretoria, South Africa, 2006. [Google Scholar]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Windell, J.T.; Bowen, S.H. Methods for study of fish diets based on analysis of stomach contents. In Methods for the Assessment of Fish Production in Freshwaters; Bagenal, T., Ed.; Blackwell: London, UK, 1978; pp. 219–226. [Google Scholar]
- Assis, C.A. A generalized index for stomach contents analysis in fish. Sci. Mar. 1996, 60, 385–389. [Google Scholar]
- Natarajan, A.V.; Jhingran, A.C. Index of preponderance-a method of grading the food elements in the stomach analysis of fishes. Indian J. Fish. 1961, 8, 54–59. [Google Scholar]
- Hurlbert, S.H. The measurement of niche overlap and some relatives. Ecology 1978, 59, 67–77. [Google Scholar] [CrossRef]
- Cortés, E. A critical review of methods of studying fish feeding based on analysis of stomach contents: Application to e1asmobranch fishes. Can. J. Fish. Aquat. Sci. 1997, 54, 726–738. [Google Scholar] [CrossRef]
- Chipps, S.R.; Garvey, J.E. Assessment of diets and feeding patterns. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2007; pp. 473–514. [Google Scholar]
- Aitchison, J. Principal Component analysis of compositional data. Biometrika 1983, 70, 57–65. [Google Scholar] [CrossRef]
- Bozza, A.N.; Hahn, N.S. Uso de recursosalimentares por peixesimaturos e adultos de species piscívorasemumaplanície de in undação neotropical. Biota Neotrop. 2010, 10, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Meurer, S.; Zaniboni-Filho, E. Reproductive and feeding biology of Acestrorhynchus pantaneiro Menezes, 1992 (Osteichthyes: Acestrorhynchidae) in areas under the influence of dams in the upper Uruguay River, Brazil. Neotrop. Ichth. 2012, 10, 159–166. [Google Scholar] [CrossRef]
- Wakjira, M. Feeding Habits and Some Biological Aspects of Fish Species in Gilgel Gibe Reservoir, Ethiopia. Int. J. Curr. Res. 2013, 5, 4124–4132. [Google Scholar]
- Wakjira, M. Fish Diversity, Community Structure, Feeding Ecology, and Fisheries of Lower Omo River and the Ethiopian Part of Lake Turkana, East Africa. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2016. [Google Scholar]
- Bwanika, G.; Murie, D.; Chapman, L. Comparative Age and Growth of Nile tilapia (Oreochromis niloticus L.) in Lakes Wamala, Uganda. Hydrobiology 2007, 589, 287–301. [Google Scholar] [CrossRef]
- Tudorancea, C.; Fernando, F.; Paggi, J. Food and feeding of O. niloticus (Linnaeus, 1758) juveniles in Lake Hawassa, Ethiopia. Arch. Hydrobiol. Suppl. 1988, 79, 267–289. [Google Scholar]
- Tadesse, Z. Food and Feeding Ecology of Tilapia, Oreochromis niloticus L. and Effects of Diet on the Lipid Quality of Fish in Some Lakes in Ethiopia. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 1998. [Google Scholar]
- Tefera, G. The Composition and Nutritional Status of the Diet of O. niloticus L. (Pisces: Cichlidae) in Lake Chamo, Ethiopia. J. Fish Biol. 1993, 42, 865–874. [Google Scholar]
- Kohl, K.D.; Coogan, S.C.P.; Raubenheimer, D. Do wild carnivores forage for prey or for nutrients? Evidence for nutrient-specific foraging. BioEssays 2015, 37, 701–709. [Google Scholar] [CrossRef]
- Battarbee, R.W. Palaeolimnological approaches to climate change, with special regard to the biological record. Quat. Sci. Rev. 2000, 19, 107–124. [Google Scholar] [CrossRef]
- Njiru, M.; Okeyo-Owuor, J.B.; Muchiri, M.; Cowx, I.G. Shifts in the food of Nile tilapia, O. niloticus (L.) in Lake Victoria, Kenya. Afr. J. Ecol. 2004, 42, 163–170. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Takken, W.; Githeko, A.K.; Jacobs, A.F.G. The effect of water turbidity on the near-surface water temperature of larval habitats of the malaria mosquito Anopheles gambiae. Int. J. Biometeorol. 2008, 52, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergeay, J.; Verschuren, D.; Meester, L.D. Invasion of an asexual American water flea clone throughout Africa and rapid displacement of a native sibling species. Proc. Biol. Sci. 2006, 273, 2839–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okogwu, O.I. Seasonal variations of species composition and abundance of zooplankton in Ehoma Lake, a floodplain lake in Nigeria. Int. J. Trop. Biol. 2010, 58, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Ayoade, A.; Fagade, S.; Adebisi, A. Diet and dietary habits of the fish Schilbemystus (Siluriformes: Schilbeidae) in two artificial lakes in Southwestern Nigeria. Int. J. Trop. Biol. 2008, 56, 1847–1855. [Google Scholar]
- Benavides, A.; Cancino, J.; Ojeda, F. Ontogenetic change in stomach dimensions and microalgal digestibility in marine herbivore fish A. punctatus. Funct. Ecol. 1994, 8, 46–55. [Google Scholar] [CrossRef]
- German, D.P. Inside the guts of wood-eating catWshes: Can they digest wood? J. Comput. Phys. Biol. 2009, 179, 1011–1023. [Google Scholar]
- Miranda, L.E.; Killgore, K.J.; Slack, W.T. Spatial organization of fish diversity in a species-rich basin. River Res. Appl. 2018, 35, 188–196. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stomach Volume | No. of Fish | Percent (%) |
|---|---|---|
| Distended | 149 | 9.0 |
| Full | 363 | 21.9 |
| 3/4 full | 345 | 20.8 |
| 1/2 full | 247 | 14.9 |
| 1/4 full | 298 | 18.0 |
| Empty | 256 | 15.4 |
| Total | 1658 | 100 |
| Food Type | %Oi | %Vi | IP | %IP |
|---|---|---|---|---|
| Phytoplankton | 100.0 | 64.3 | 6825.0 | 74.5 |
| Detritus | 99.5 | 14.6 | 1462.0 | 15.9 |
| Zooplankton | 87.1 | 12.8 | 576.7 | 6.3 |
| Macrophytes | 65.8 | 4.7 | 222.7 | 2.4 |
| Insects | 8.6 | 1.6 | 13.8 | 0.2 |
| Fish parts | 15.9 | 1.4 | 22.5 | 0.3 |
| Nematodes | 7.8 | 0.3 | 2.2 | 0.02 |
| Ostracods | 10.1 | 0.6 | 6.5 | 0.1 |
| Unidentified | 6.04 | 1.3 | 35.4 | 0.4 |
| Food Type | Specific Items | %Oi | %Vi | IP | %IP |
|---|---|---|---|---|---|
| Phytoplankton | Cyanophyta (Blue Green algae) | 100.0 | 4.8 | 477.0 | 10.1 |
| Bacillariophyta (Diatom) | 99.5 | 16.9 | 1684.5 | 35.6 | |
| Chlorophyta (Green algae) | 78.0 | 9.9 | 778.4 | 16.5 | |
| Chrysophyta | 32.4 | 2.6 | 84.6 | 1.8 | |
| Cryptophyta | 47.6 | 6.9 | 330.8 | 7.0 | |
| Dinophyta | 72.6 | 6.1 | 440.7 | 9.3 | |
| Rhodophyta | 16.8 | 3.0 | 51.1 | 1.1 | |
| Euglenophyta | 74.8 | 11.3 | 843.7 | 17.9 | |
| Heterokontophyta | 14.4 | 2.6 | 37.3 | 0.8 | |
| Zooplankton | Anomopoda | 6.0 | 0.4 | 2.6 | 0.5 |
| Cladocera | 33.6 | 3.7 | 123.5 | 21.4 | |
| Ctenopoda | 12.3 | 1.0 | 12.5 | 2.2 | |
| Copepoda | 48.3 | 3.8 | 183.1 | 31.8 | |
| Rotifera | 76.9 | 3.2 | 247.9 | 43.0 | |
| Ostracoda | 10.1 | 0.6 | 6.5 | 1.1 | |
| Aquatic insects | Diptera | 4.7 | 10.3 | 49.5 | 10.4 |
| Plecoptera | 4.2 | 12.7 | 52.9 | 11.1 | |
| Trichoptera | 3.7 | 11.8 | 43.3 | 9.1 | |
| Chilopoda | 4.8 | 14.5 | 70.1 | 14.7 | |
| Coleoptera | 3.8 | 6.96 | 26.1 | 5.5 | |
| Odonata | 8.4 | 13.7 | 114.3 | 23.9 | |
| Hemiptera | 5.0 | 8.2 | 41.0 | 8.6 | |
| Ephemeroptera | 4.6 | 11.5 | 52.8 | 11.1 | |
| Hymenoptera | 2.7 | 10.4 | 27.8 | 5.8 |
| Correlation Coefficients | Canonical Coefficient | |
|---|---|---|
| Axis 1 | Axis 2 | |
| Eigenvalue | 9.19 | 0.62 |
| % of variance | 90.37 | 6.10 |
| Horakelo | 0.3474 | −0.0846 |
| Hoitu | 0.5645 | −0.4115 |
| Dole | 0.2709 | 0.5482 |
| Webishebele | 0.5937 | −0.04752 |
| Middle | 0.1505 | 0.7145 |
| Tufa | 0.3348 | 0.1011 |
| Correlation Coefficients | Canonical Coefficient | |
|---|---|---|
| Axis 1 | Axis 2 | |
| Eigenvalue | 18.36 | 1.82 |
| % of variance | 80.45 | 7.97 |
| Phytoplankton | −0.90 | 0.79 |
| Zooplankton | 0.89 | 0.35 |
| Detritus | 0.95 | 0.30 |
| Insects | 0.87 | 0.21 |
| Macrophytes | 0.83 | 0.08 |
| Fish parts | 0.11 | −0.37 |
| Nematode | 0.64 | 0.18 |
| Unidentified | 0.28 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Temesgen, M.; Getahun, A.; Lemma, B.; Janssens, G.P.J. Food and Feeding Biology of Nile Tilapia (Oreochromis niloticus) in Lake Langeno, Ethiopia. Sustainability 2022, 14, 974. https://doi.org/10.3390/su14020974
Temesgen M, Getahun A, Lemma B, Janssens GPJ. Food and Feeding Biology of Nile Tilapia (Oreochromis niloticus) in Lake Langeno, Ethiopia. Sustainability. 2022; 14(2):974. https://doi.org/10.3390/su14020974
Chicago/Turabian StyleTemesgen, Mathewos, Abebe Getahun, Brook Lemma, and Geert P. J. Janssens. 2022. "Food and Feeding Biology of Nile Tilapia (Oreochromis niloticus) in Lake Langeno, Ethiopia" Sustainability 14, no. 2: 974. https://doi.org/10.3390/su14020974
APA StyleTemesgen, M., Getahun, A., Lemma, B., & Janssens, G. P. J. (2022). Food and Feeding Biology of Nile Tilapia (Oreochromis niloticus) in Lake Langeno, Ethiopia. Sustainability, 14(2), 974. https://doi.org/10.3390/su14020974