



Aquatic Ecological Risk Evaluation of Chiral Triazole Fungicide Prothioconazole and Its Metabolite Prothioconazole-Desthio on Lemna minor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Plant Material and Cultivation
2.3. Acute-Toxicity Experiment
2.4. Physiological and Biochemical Experiment
2.4.1. Determination of Photosynthetic Pigment Content
2.4.2. Determination of Antioxidant Enzymes Activities and Lipid Peroxidation
2.5. Statistical Analysis
3. Results and Discussion
3.1. Enantioselective Acute Toxicity
3.2. Effect of PTZ and PTZ Enantiomer on Dry Weight and Photosynthetic Pigment
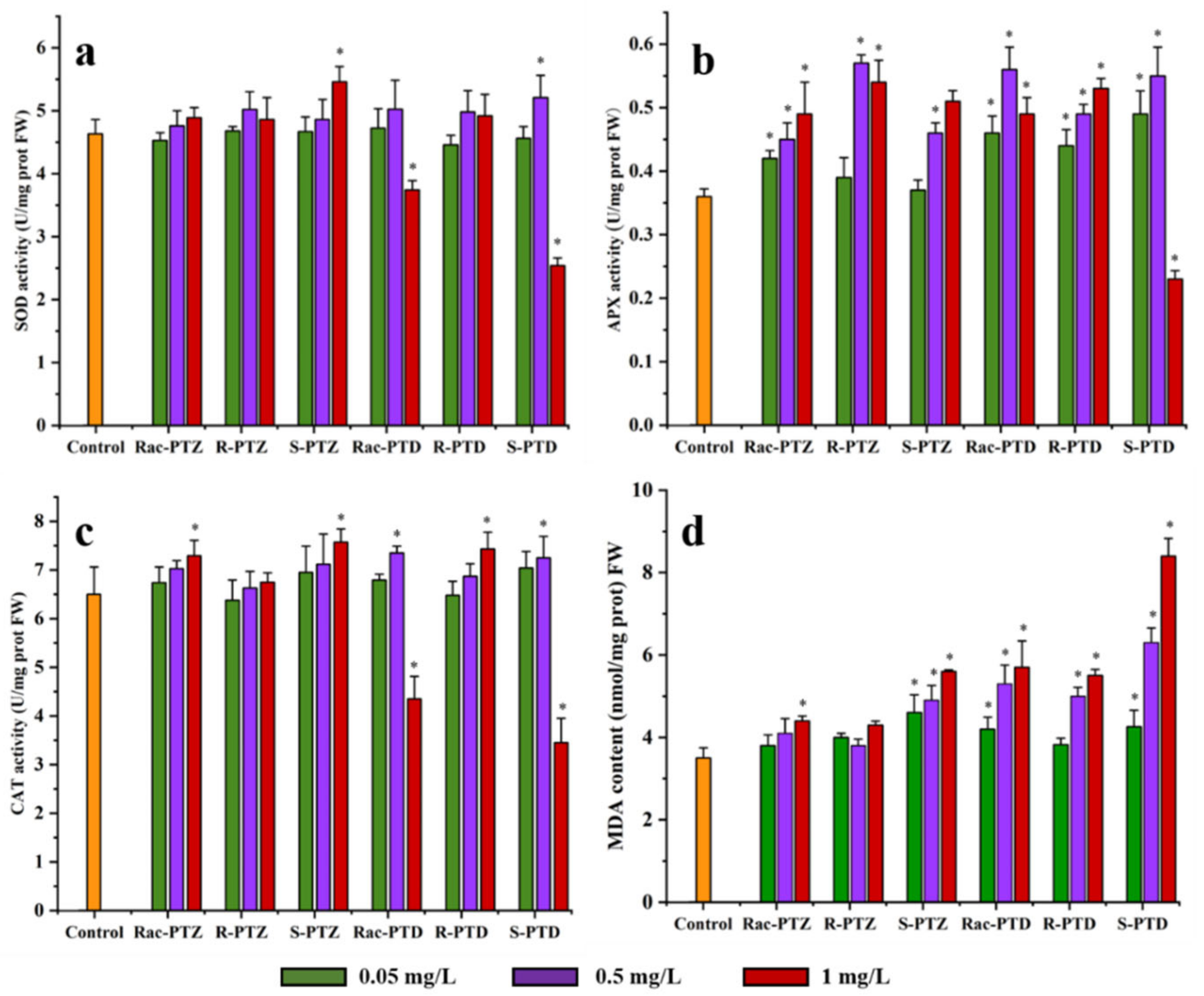
3.3. Antioxidant-Enzyme Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parker, J.E.; Warrilow, A.G.S.; Cools, H.J.; Fraaije, B.A.; Lucas, J.A.; Rigdova, K.; Griffiths, W.J.; Kelly, D.E.; Kelly, S.L. Prothioconazole and Prothioconazole-Desthio Activities against Candida Albicans Sterol 14-α-Demethylase. Appl. Environ. Microbiol. 2013, 79, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, O.; Waliullah, S.; Ji, P.; Ali, M.E. Molecular Characterization of Laboratory Mutants of Fusarium Oxysporum F. Sp. Niveum Resistant to Prothioconazole, a Demethylation Inhibitor (DMI) Fungicide. J. Fungi 2021, 7, 704. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, C. Enantioselective Effect of Chiral Fungicide Prothioconazole on Fusarium Graminearum: Fungicidal Activity and Don Biosynthesis. Environ. Pollut. 2022, 307, 119553. [Google Scholar] [CrossRef] [PubMed]
- Breunig, M.; Chilvers, M.I. Comparison between Prothioconazole and Prothioconazole-Desthio in Poison-Plate Mycelial Growth Assays of Fusarium Graminearum. Plant Health Prog. 2022, 23, 159–161. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, B.; Li, L.; Zhang, Q.; Xia, W.; Wang, M. Enantioselective Degradation and Transformation of the Chiral Fungicide Prothioconazole and Its Chiral Metabolite in Soils. Sci. Total Environ. 2018, 634, 875–883. [Google Scholar] [CrossRef]
- Jing, X.; Huang, X.; Wang, H.; Xue, H.; Wu, B.; Wang, X.; Jia, L. Popping candy-assisted dispersive liquid–liquid microextraction for enantioselective determination of prothioconazole and its chiral metabolite in water, beer, Baijiu, and vinegar samples by HPLC. Food Chem. 2021, 348, 129147. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Gao, B.; Gou, G.; Li, L.; Shi, H.; Wang, M. Simultaneous Enantioselective Determination of the Chiral Fungicide Prothioconazole and Its Major Chiral Metabolite Prothioconazole-Desthio in Food and Environmental Samples by Ultraperformance Liquid Chromatography–Tandem Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 8241–8247. [Google Scholar] [CrossRef]
- Jiang, D.; Dong, F.; Xu, J.; Liu, X.; Wu, X.; Pan, X.; Tao, Y.; Li, R.; Zheng, Y. Enantioselective Separation and Dissipation of Prothioconazole and Its Major Metabolite Prothioconazole-Desthio Enantiomers in Tomato, Cucumber, and Pepper. J. Agric. Food Chem. 2019, 67, 10256–10264. [Google Scholar] [CrossRef]
- Dong, X.; Tong, Z.; Chu, Y.; Sun, M.; Wang, M.; Gao, T.; Duan, J. Dissipation of Prothioconazole and Its Metabolite Prothioconazole-Desthio in Rice Fields and Risk Assessment of Its Dietary Intake. J. Agric. Food Chem. 2019, 67, 6458–6465. [Google Scholar] [CrossRef]
- Lin, H.; Dong, B.; Hu, J. Residue and Intake Risk Assessment of Prothioconazole and Its Metabolite Prothioconazole-Desthio in Wheat Field. Environ. Monit. Assess. 2017, 189, 236. [Google Scholar] [CrossRef]
- Tian, S.; Yan, H.; Meng, Z.; Jia, M.; Sun, W.; Huang, S.; Wang, Y.; Zhou, Z.; Diao, J.; Zhu, W. Prothioconazole and Prothioconazole-Desthio Induced Different Hepatotoxicities Via Interfering with Glycolipid Metabolism in Mice. Pestic. Biochem. Physiol. 2022, 180, 104983. [Google Scholar] [CrossRef]
- Meng, Z.; Tian, S.; Sun, W.; Liu, L.; Yan, S.; Huang, S.; Zhu, W.; Zhou, Z. Effects of Exposure to Prothioconazole and Its Metabolite Prothioconazole-Desthio on Oxidative Stress and Metabolic Profiles of Liver and Kidney Tissues in Male Mice. Environ. Pollut. 2021, 269, 116215. [Google Scholar] [CrossRef]
- Xie, Y.; Jiang, H.; Chang, J.; Wang, Y.; Li, J.; Wang, H. Gonadal Disruption after Single Dose Exposure of Prothioconazole and Prothioconazole-Desthio in Male Lizards (Eremias argus). Environ. Pollut. 2019, 255, 113297. [Google Scholar] [CrossRef]
- Xie, Y.; Li, L.Y.Z.; Hao, W.; Chang, J.; Xu, P.; Guo, B.; Li, J.; Wang, H. Comparative Toxicokinetics and Tissue Distribution of Prothioconazole and Prothioconazole-Desthio in Chinese Lizards (Eremias argus) and Transcriptional Responses of Metabolic-Related Genes. Environ. Pollut. 2019, 247, 524–533. [Google Scholar] [CrossRef]
- Zhang, Z.; Du, G.; Gao, B.; Hu, K.; Kaziem, A.E.; Li, L.; He, Z.; Shi, H.; Wang, M. Stereoselective Endocrine-Disrupting Effects of the Chiral Triazole Fungicide Prothioconazole and Its Chiral Metabolite. Environ. Pollut. 2019, 251, 30–36. [Google Scholar] [CrossRef]
- Butkovskyi, A.; Jing, Y.; Bergheim, H.; Lazar, D.; Gulyaeva, K.; Odenmarck, S.R.; Norli, H.R.; Nowak, K.M.; Miltner, A.; Kästner, M.; et al. Retention and Distribution of Pesticides in Planted Filter Microcosms Designed for Treatment of Agricultural Surface Runoff. Sci. Total Environ. 2021, 778, 146114. [Google Scholar] [CrossRef]
- Sun, Y.; Cao, Y.; Tong, L.; Tao, F.; Wang, X.; Wu, H.; Wang, M. Exposure to Prothioconazole Induces Developmental Toxicity and Cardiovascular Effects on Zebrafish Embryo. Chemosphere 2020, 251, 126418. [Google Scholar] [CrossRef]
- Shen, J.; Liu, P.; Sun, Y.; Xu, X.; Guo, L.; Rao, Q.; Chen, M.; Liu, X. Embryonic Exposure to Prothioconazole Induces Oxidative Stress and Apoptosis in Zebrafish (Danio rerio) Early Life Stage. Sci. Total Environ. 2021, 756, 143859. [Google Scholar] [CrossRef]
- Tian, S.; Teng, M.; Meng, Z.; Yan, S.; Jia, M.; Li, R.; Liu, L.; Yan, J.; Zhou, Z.; Zhu, W. Toxicity Effects in Zebrafish Embryos (Danio Rerio) Induced by Prothioconazole. Environ. Pollut. 2019, 255, 113269. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Zhao, X.; Gao, B.; He, Z.; Li, L.; Shi, H.; Wang, M. Stereoselective Uptake and Metabolism of Prothioconazole Caused Oxidative Stress in Zebrafish (Danio rerio). J. Hazard. Mater. 2020, 396, 122756. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a Model Plant System in the Genomics and Postgenomics Era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Vermaat, J.E.; Hanif, M.K. Performance of Common Duckweed Species (Lemnaceae) and the Waterfern Azolla Filiculoides on Different Types of Waste Water. Water Res. 1998, 32, 2569–2576. [Google Scholar] [CrossRef]
- ISO 20079:2005; Water Quality—Determination of the Toxic Effect of Water Constituents and Waste Water on Duckweed (Lemna minor)—Duckweed Growth Inhibition Test. ISO: Vernier, Switzerland, 2005.
- OECD. Test No. 221: Lemna Sp. Growth Inhibition Test; OECD: Paris, France, 2006. [Google Scholar]
- Wang, F.; Liu, D.; Qu, H.; Chen, L.; Zhou, Z.; Wang, P. A Full Evaluation for the Enantiomeric Impacts of Lactofen and Its Metabolites on Aquatic Macrophyte Lemna Minor. Water Res. 2016, 101, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Luo, C.; Qiu, J.; Li, Y.; Zhang, H.; Tan, H. Metabolomic and Transcriptomic Investigation of the Mechanism Involved in Enantioselective Toxicity of Imazamox in Lemna minor. J. Hazard. Mater. 2022, 425, 127818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xie, Y.; Ye, Y.; Yang, Y.; Hua, R.; Wu, X. Toxification Metabolism and Treatment Strategy of the Chiral Triazole Fungicide Prothioconazole in Water. J. Hazard. Mater. 2022, 432, 128650. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, B.; He, Z.; Li, L.; Zhang, Q.; Kaziem, A.E.; Wang, M. Stereoselective Bioactivity of the Chiral Triazole Fungicide Prothioconazole and Its Metabolite. Pestic. Biochem. Physiol. 2019, 160, 112–118. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Buschmann, C.; Lichtenthaler, H.K. Leaf Chlorophyll Fluorescence Corrected for Re-Absorption by Means of Absorption and Reflectance Measurements. J. Plant Physiol. 1998, 152, 283–296. [Google Scholar] [CrossRef]
- Zhai, W.; Zhang, L.; Cui, J.; Wei, Y.; Wang, P.; Liu, D.; Zhou, Z. The biological activities of prothioconazole enantiomers and their toxicity assessment on aquatic organisms. Chirality 2019, 31, 468–475. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, L.; Zhou, Q. Effects of Bisphenol A on Growth, Photosynthesis and Chlorophyll Fluorescence in Above-Ground Organs of Soybean Seedlings. Chemosphere 2013, 90, 1274–1280. [Google Scholar] [CrossRef]
- Ferreyroa, G.V.; Lagorio, M.G.; Trinelli, M.A.; Lavado, R.S.; Molina, F.V. Lead Effects on Brassica Napus Photosynthetic Organs. Ecotoxicol. Environ. Saf. 2017, 140, 123–130. [Google Scholar] [CrossRef]
- Koschnick, T.J.; Haller, W.T.; Chen, A.W. Carfentrazone-Ethyl Pond Dissipation and Efficacy on Floating Plants. J. Aquat. Plant Manage. 2004, 42, 103–108. [Google Scholar]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The Roles of Environmental Factors in Regulation of Oxidative Stress in Plant. BioMed Res. Int. 2019, 2019, 9732325. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant Enzymes Regulation in Plants in Reference to Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Song, H.; Wang, Y.S.; Sun, C.C.; Wang, Y.T.; Peng, Y.L.; Cheng, H. Effects of Pyrene on Antioxidant Systems and Lipid Peroxidation Level in Mangrove Plants, Bruguiera Gymnorrhiza. Ecotoxicology 2012, 21, 1625–1632. [Google Scholar] [CrossRef]
- Sun, B.Y.; Kan, S.H.; Zhang, Y.Z.; Deng, S.H.; Wu, J.; Yuan, H.; Qi, H.; Yang, G.; Li, L.; Zhang, X.H.; et al. Certain Antioxidant Enzymes and Lipid Peroxidation of Radish (Raphanus sativus L.) as Early Warning Biomarkers of Soil Copper Exposure. J. Hazard. Mater. 2010, 183, 833–838. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Chemical Compound | 3 d | 5 d | 7 d | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | LC50 (mg/L) | 95% CI | R2 | LC50 (mg/L) | 95% CI | R2 | LC50 (mg/L) | 95% CI | |
| Rac-PTZ | 0.99 | 10.51 | 9.23–11.56 | 0.96 | 5.63 | 5.06–6.45 | 0.96 | 2.36 | 2.07–3.15 |
| R-PTZ | 0.98 | 20.36 | 18.26–24.32 | 0.97 | 12.32 | 10.23–13.58 | 0.94 | 5.63 | 4.68–6.38 |
| S-PTZ | 0.95 | 8.69 | 7.21–9.05 | 0.93 | 4.36 | 3.86–5.12 | 0.93 | 2.06 | 1.23–2.98 |
| Rac-PTD | 0.97 | 9.36 | 8.23–10.08 | 0.95 | 3.32 | 3.09–4.87 | 0.93 | 0.98 | 0.76–1.18 |
| R-PTD | 0.93 | 16.59 | 15.69–18.75 | 0.92 | 9.26 | 7.97–10.12 | 0.96 | 2.23 | 2.06–3.56 |
| S-PTD | 0.94 | 7.59 | 6.98–8.45 | 0.94 | 2.57 | 1.89–3.45 | 0.98 | 0.76 | 0.62–0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Li, X.; Tang, Q.; Mei, L.; Cao, J.; Huang, H.; Zhang, Z. Aquatic Ecological Risk Evaluation of Chiral Triazole Fungicide Prothioconazole and Its Metabolite Prothioconazole-Desthio on Lemna minor. Sustainability 2022, 14, 16292. https://doi.org/10.3390/su142316292
Yang Y, Li X, Tang Q, Mei L, Cao J, Huang H, Zhang Z. Aquatic Ecological Risk Evaluation of Chiral Triazole Fungicide Prothioconazole and Its Metabolite Prothioconazole-Desthio on Lemna minor. Sustainability. 2022; 14(23):16292. https://doi.org/10.3390/su142316292
Chicago/Turabian StyleYang, Yaling, Xiaomeng Li, Qiyue Tang, Liangchi Mei, Jian Cao, Haiyun Huang, and Zhaoxian Zhang. 2022. "Aquatic Ecological Risk Evaluation of Chiral Triazole Fungicide Prothioconazole and Its Metabolite Prothioconazole-Desthio on Lemna minor" Sustainability 14, no. 23: 16292. https://doi.org/10.3390/su142316292
APA StyleYang, Y., Li, X., Tang, Q., Mei, L., Cao, J., Huang, H., & Zhang, Z. (2022). Aquatic Ecological Risk Evaluation of Chiral Triazole Fungicide Prothioconazole and Its Metabolite Prothioconazole-Desthio on Lemna minor. Sustainability, 14(23), 16292. https://doi.org/10.3390/su142316292