
Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Small Mammal Sampling
2.3. Vegetation Structure
2.4. Data Analysis
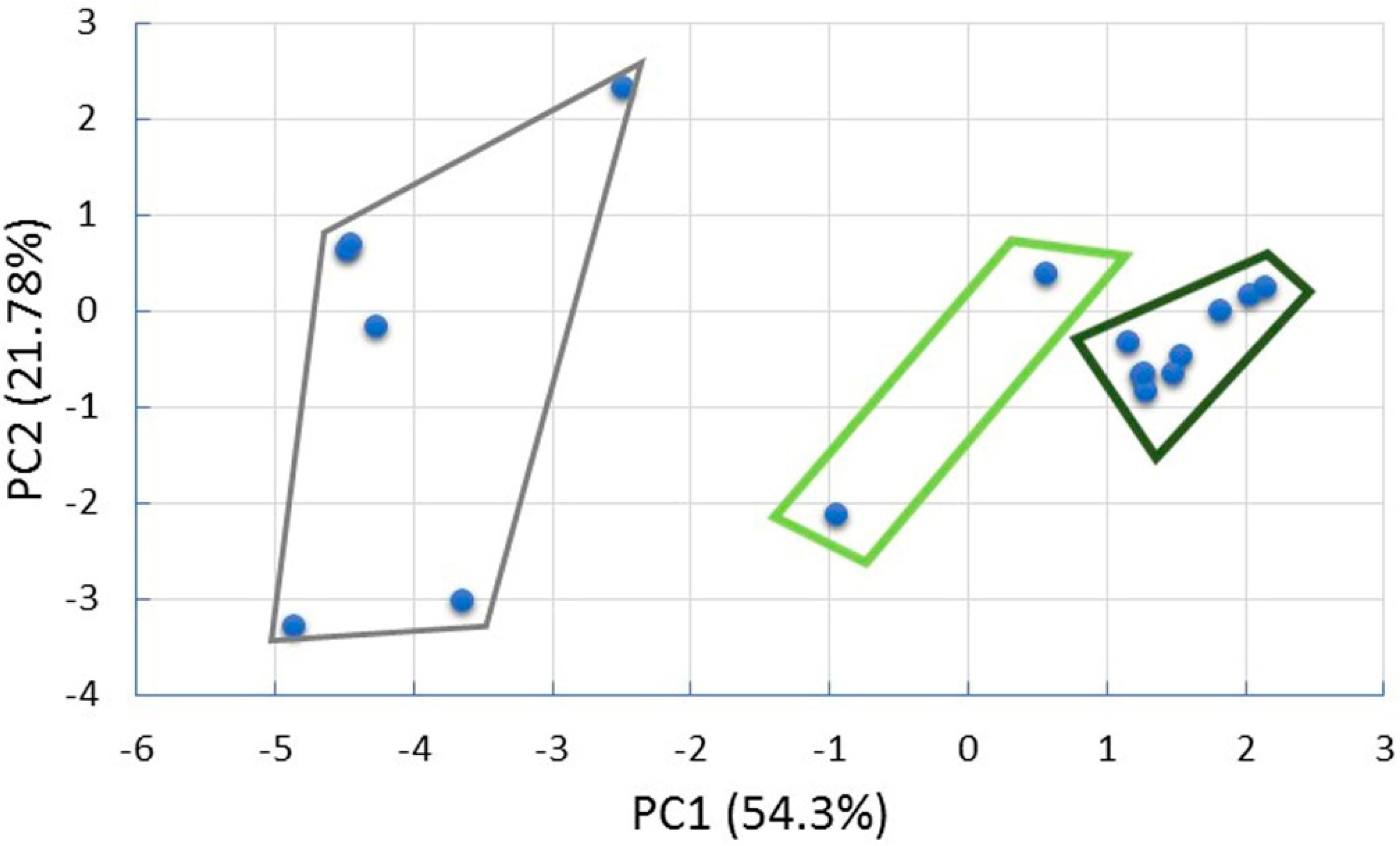
3. Results
3.1. Vegetation Structure
3.2. Small Mammals and Vegetation Structure Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazor, T.; Doropoulos, C.; Schwarzmueller, F.; Gladish, D.W.; Kumaran, N.; Merkel, K.; Di Marco, M.; Gagic, V. Global mismatch of policy and research on drivers of biodiversity loss. Nat. Ecol. Evol. 2018, 2, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G. The Mediterranean Region. Biological Diversity in Space and Time. Available online: https://global.oup.com/academic/product/the-mediterranean-region-9780199557998?cc=es&lang=en& (accessed on 8 December 2021).
- Valladares, F.; Benavides, R.; Rabasa, S.G.; Díaz, M.; Pausas, J.G.; Paula, S.; Simonson, W.D. Global change and Mediterranean forests: Current impacts and potential responses. In Forests and Global Change; Cambridge University Press: Cambridge, UK, 2014; pp. 47–76. [Google Scholar]
- Herrando, S.; Anton, M.; Sardà-Palomera, F.; Bota, G.; Gregory, R.D.; Brotons, L. Indicators of the impact of land use changes using large-scale bird surveys: Land abandonment in a Mediterranean region. Ecol. Indic. 2014, 45, 235–244. [Google Scholar] [CrossRef]
- Regos, A.; D’Amen, M.; Titeux, N.; Herrando, S.; Guisan, A.; Brotons, L. Predicting the future effectiveness of protected areas for bird conservation in Mediterranean ecosystems under climate change and novel fire regime scenarios. Divers. Distrib. 2016, 22, 83–96. [Google Scholar] [CrossRef]
- Brotons, L.; Herrando, S.; Sirami, C.; Kati, V.; Díaz, M. Mediterranean Forest Bird Communities and the Role of Landscape Heterogeneity in Space and Time. Ecol. Conserv. For. Birds 2018, 9, 318–349. [Google Scholar] [CrossRef]
- Moghli, A.; Santana, V.M.; Baeza, M.J.; Pastor, E.; Soliveres, S. Fire Recurrence and Time Since Last Fire Interact to Determine the Supply of Multiple Ecosystem Services by Mediterranean Forests. Ecosystems 2021, 1–13. [Google Scholar] [CrossRef]
- Pausas, J.G.; Fernández-Muñoz, S. Fire regime changes in the Western Mediterranean Basin: From fuel-limited to drought-driven fire regime. Clim. Chang. 2012, 110, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Torre, I.; Díaz, M. Small mammal abundance in Mediterranean post-fire habitats: A role for predators? Acta Oecologica 2004, 25, 137–142. [Google Scholar] [CrossRef]
- Torre, I.; Bastardas-Llabot, J.; Arrizabalaga, A.; Díaz, M. Population dynamics of small endotherms under global change: Greater white-toothed shrews Crocidura russula in Mediterranean habitats. Sci. Total Environ. 2020, 705, 135799. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.; Zellweger, F.; Bollmann, K.; Jones, G.; Obrist, M.K. From field surveys to LiDAR: Shining a light on how bats respond to forest structure. Remote Sens. Environ. 2016, 175, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Vierling, K.T.; Vierling, L.A.; Gould, W.A.; Martinuzzi, S.; Clawges, R.M. Lidar: Shedding new light on habitat characterization and modeling. Front. Ecol. Environ. 2008, 6, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Fotis, A.T.; Patel, S.; Chavez, A.S. Habitat-based isolating barriers are not strong in the speciation of ecologically divergent squirrels (Tamiasciurus douglasii and T. hudsonicus). Behav. Ecol. Sociobiol. 2020, 74, 1–14. [Google Scholar] [CrossRef]
- Schooler, S.L.; Zald, H.S.J. Lidar Prediction of Small Mammal Diversity in Wisconsin, USA. Remote Sens. 2019, 11, 2222. [Google Scholar] [CrossRef] [Green Version]
- Linnell, M.A.; Davis, R.J.; Lesmeister, D.B.; Swingle, J.K. Conservation and relative habitat suitability for an arboreal mammal associated with old forest. For. Ecol. Manag. 2017, 402, 1–11. [Google Scholar] [CrossRef]
- Jaime-González, C.; Acebes, P.; Mateos, A.; Mezquida, E.T. Bridging gaps: On the performance of airborne LiDAR to model wood mouse-habitat structure relationships in pine forests. PLoS ONE 2017, 12, e0182451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doblas-Miranda, E.; Martínez-Vilalta, J.; Lloret, F.; Álvarez, A.; Ávila, A.; Bonet, F.J.; Brotons, L.; Castro, J.; Curiel Yuste, J.; Díaz, M.; et al. Reassessing global change research priorities in mediterranean terrestrial ecosystems: How far have we come and where do we go from here? Glob. Ecol. Biogeogr. 2015, 24, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, A.; Bonal, R.; Díaz, M. Ungulates, rodents, shrubs: Interactions in a diverse Mediterranean ecosystem. Basic Appl. Ecol. 2009, 10, 151–160. [Google Scholar] [CrossRef]
- Carrascal, L.M.; Villén-Pérez, S.; Seoane, J. Thermal, Food and Vegetation Effects on Winter Bird Species Richness of Mediterranean Oakwoods. Ecol. Res. 2012, 27, 293–302. [Google Scholar] [CrossRef]
- Martínez-Jauregui, M.; Díaz, M.; Sánchez de Ron, D.; Soliño, M. Plantation or natural recovery? Relative contribution of planted and natural pine forests to the maintenance of regional bird diversity along ecological gradients in Southern Europe. For. Ecol. Manag. 2016, 376, 183–192. [Google Scholar] [CrossRef]
- Baśnou, C.; Álvarez, E.; Bagaria, G.; Guardiola, M.; Isern, R.; Vicente, P.; Pino, J. Spatial patterns of land use changes across a mediterranean metropolitan landscape: Implications for biodiversity management. Environ. Manag. 2013, 52, 971–980. [Google Scholar] [CrossRef]
- Riera, J.; Castell, C. Efectes dels incendis forestals recurrents sobre la distribució de dues espècies del Parc Natural del Garraf: El pi blanc (Pinus halepensis) i la savina (Juniperus phoenicea). Butlletí Inst. Catalana D’Història Nat. 1997, 65, 105–116. [Google Scholar]
- Puerta-Pinero, C.; Espelta, J.M.; Sanchez-Humanes, B.; Rodrigo, A.; Coll, L.; Brotons, L. History matters: Previous land use changes determine post-fire vegetation recovery in forested Mediterranean landscapes. For. Ecol. Manag. 2012, 279, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Torre, I.; Raspall, A.; Arrizabalaga, A.; Díaz, M. SEMICE: An unbiased and powerful monitoring protocol for small mammals in the Mediterranean Region. Mamm. Biol. 2018, 88, 161–167. [Google Scholar] [CrossRef]
- Torre, I.; López-Baucells, A.; Stefanescu, C.; Freixas, L.; Flaquer, C.; Bartrina, C.; Coronado, A.; López-Bosch, D.; Mas, M.; Míguez, S.; et al. Concurrent Butterfly, Bat and Small Mammal Monitoring Programmes Using Citizen Science in Catalonia (NE Spain): A Historical Review and Future Directions. Diversity 2021, 13, 454. [Google Scholar] [CrossRef]
- Torre, I.; Raspall, A.; Arrizabalaga, A.; Díaz, M. Evaluating trap performance and volunteers’ experience in small mammal monitoring programs based on citizen science: The SEMICE case study. Mamm. Biol. 2019, 95, 26–30. [Google Scholar] [CrossRef]
- Torre, I.; Arrizabalaga, A. Species richness and abundance of small mammals along an elevational gradient of a Mediterranean mountain. Vie Milieu 2009, 59, 199–208. [Google Scholar]
- Carrilho, M.; Teixeira, D.; Santos-Reis, M.; Rosalino, L.M. Small mammal abundance in Mediterranean Eucalyptus plantations: How shrub cover can really make a difference. For. Ecol. Manag. 2017, 391, 256–263. [Google Scholar] [CrossRef]
- Fons, R.; Grabulosa, I.; Saint Girons, M.C.; Galan-Puchades, M.T.; Feliu, C. Incendie et cicatrisation des ecosystemes mediterraneens dynamique du repeuplement en micromammiferes. Vie Milieu 1988, 38, 259–280. [Google Scholar]
- Puig-Gironès, R.; Imbeau, L.; Clavero, M.; Rost, J.; Pons, P. Does post-fire salvage logging affect foraging activity by rodents? Eur. J. For. Res. 2020, 139, 777–790. [Google Scholar] [CrossRef]
- Caceres, N.C.; Napoli, R.P.; Hannibal, W. Differential trapping success for small mammals using pitfall and standard cage traps in a woodland savannah region of southwestern Brazil. Mammalia 2011, 75, 45–52. [Google Scholar] [CrossRef]
- Nicolas, V.; Colyn, M. Relative efficiency of three types of small mammal traps in an African rainforest. Belg. J. Zool. 2006, 136, 107–111. [Google Scholar]
- Torre, I.; Freixas, L.; Arrizabalaga, A.; Díaz, M. The efficiency of two widely used commercial live-traps to develop monitoring protocols for small mammal biodiversity. Ecol. Indic. 2016, 66, 481–487. [Google Scholar] [CrossRef]
- Sikes, R.S. Animal Care and use Committee of the American Society of Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef] [PubMed]
- Gurnell, J.; Flowerdew, J.R. Live Trapping Small Mammals, A Practical Guide; The Mammal Society: London, UK, 2006. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar Remote Sensing for Ecosystem Studies. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Simonson, W.D.; Allen, H.D.; Coomes, D.A. Applications of airborne lidar for the assessment of animal species diversity. Methods Ecol. Evol. 2014, 5, 719–729. [Google Scholar] [CrossRef]
- Vierling, K.T.; Swift, C.E.; Hudak, A.T.; Vogeler, J.C.; Vierling, L.A. How much does the time lag between wildlife field-data collection and LiDAR-data acquisition matter for studies of animal distributions A case study using bird communities. Remote Sens. Lett. 2014, 5, 185–193. [Google Scholar] [CrossRef]
- McGaughey, R.; Carson, W. Fusing LIDAR data, photographs, and other data using 2D and 3D visualization techniques. In Proceedings of the Terrain Data: Applications and Visualization–Making the Connection, Seattle, WA, USA, 28–30 October 2003. [Google Scholar]
- Isenburg, M. LAStools: Converting, Filtering, Viewing, Processing, and Compressing LIDAR Data in LAS Format. Available online: https://www.cs.unc.edu/~isenburg/lastools/ (accessed on 8 December 2021).
- Heurich, M.; Fischer, F.; Knorzer, O.; Krzystek, P. Assessment of Digital Terrain Models (DTM) from Data Gathered with Airborne Laser Scanning in Temperate European Beech (Fagus Sylvatica) and Norway Spruce (Picea Abies) Forests. Photogramm. Fernerkund. Geoinf. 2008, 6, 473–488. [Google Scholar]
- Dytham, C. Choosing and Using Statistics. A Biologist’s Guide; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar]
- Harrell, F.E.; Lee, K.L.; Mark, D.B. Multivariable prognostic models: Issues in developing models, evaluating assumptions and adequacy, and measuring and reducing errors. Stat. Med. 1996, 15, 361–387. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Barton, K. Package “MuMIn” Multi-Model Inference 2020. Available online: www.cran.r-project.org/web/packages (accessed on 1 January 2019).
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples, Sotware and User’s Guide. 2012.
- Kelly, L.T.; Nimmo, D.C.; Spence-Bailey, L.M.; Clarke, M.F.; Bennett, A.F. The short-term responses of small mammals to wildfire in semiarid mallee shrubland, Australia. Wildl. Res. 2010, 37, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Horn, K.J.; McMillan, B.R.; St. Clair, S.B. Expansive fire in Mojave Desert shrubland reduces abundance and species diversity of small mammals. J. Arid Environ. 2012, 77, 54–58. [Google Scholar] [CrossRef]
- Doherty, T.S.; Davis, R.A.; Van Etten, E.J.B. A game of cat-and-mouse: Microhabitat influences rodent foraging in recently burnt but not long unburnt shrublands. J. Mammal. 2015, 96, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.; Torre, I.; Arrizabalaga, A. Relative roles of density and rainfall on the short-term regulation of Mediterranean wood mouse Apodemus sylvaticus populations. Acta Theriol. 2010, 55, 251–260. [Google Scholar] [CrossRef]
- Sunyer, P.; Muñoz, A.; Mazzerole, M.J.; Bonal, R.; Espelta, J.M. Wood mouse population dynamics: Interplay among seed abundance seasonality, shrub cover and wild boar interference. Mamm. Biol. 2016, 81, 372–379. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Pausas, J.G. Does plant richness influence animal richness? the mammals of Catalonia (NE Spain). Divers. Distrib. 2004, 10, 247–252. [Google Scholar] [CrossRef]
- Teixeira, D.; Carrilho, M.; Mexia, T.; Kobel, M.; Santos, M.J.; Santos-Reis, M.; Rosalino, L.M. Management of Eucalyptus plantations influences small mammal density: Evidence from Southern Europe. For. Ecol. Manag. 2017, 385, 25–34. [Google Scholar] [CrossRef]
- Loggins, A.A.; Shrader, A.M.; Monadjem, A.; McCleery, R.A. Shrub cover homogenizes small mammals’ activity and perceived predation risk. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Stefanescu, C.; Soldevila, A.; Gutiérrez, C.; Torre, I.; Ubach, A.; Miralles, M. Explosions demogràfiques de l’eruga peluda del suro, Lymantria dispar (Linnaeus, 1758), als boscos del Montnegre el 2019 i 2020: Possibles causes, impactes i idoneïtat dels tractaments per combatre la plaga. Butlletí Inst. Catalana d’Història Nat. 2020, 84, 267–279. [Google Scholar]
- Lima, M.; Stenseth, N.C.; Leirs, H.; Jaksic, F.M. Population dynamics of small mammals in semi-arid regions: A comparative study of demographic variability in two rodent species. Proc. R. Soc. B Biol. Sci. USA 2003, 270, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, W.I. Population regulation in the wood mouse, Apodemus sylvaticus.I. density dependence in the annual cycle of abundance. J. Anim. Ecol. 1989, 58, 465–475. [Google Scholar] [CrossRef]
- Massoud, D.; Lao-Pérez, M.; Ortega, E.; Burgos, M.; Jiménez, R.; Barrionuevo, F.J. Divergent seasonal reproductive patterns in syntopic populations of two murine species in southern spain, mus spretus and apodemus sylvaticus. Animals 2021, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitatheterogeneity/diversity: The importanceof keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Morales-Diaz, S.P.; Yolotl Alvarez-Anorve, M.; Edith Zamora-Espinoza, M.; Dirzo, R.; Oyama, K.; Daniel Avila-Cabadilla, L. Rodent community responses to vegetation and landscape changes in early successional stages of tropical dry forest. For. Ecol. Manag. 2019, 433, 633–644. [Google Scholar] [CrossRef]
- Williams, S.E.; Marsh, H.; Winter, J. Spatial scale, species diversity, and habitat structure: Small mammals in Australian tropical rain forest. Ecology 2002, 83, 1317–1329. [Google Scholar] [CrossRef]
- de Camargo, N.F.; Sano, N.Y.; Vieira, E.M. Forest vertical complexity affects alpha and beta diversity of small mammals. J. Mammal. 2018, 99, 1444–1454. [Google Scholar] [CrossRef]
- Buesching, C.D.; Newman, C.; Twell, R.; Macdonald, D.W. Reasons for arboreality in wood mice Apodemus sylvaticus and Bank voles Myodes glareolus. Mamm. Biol. 2008, 73, 318–324. [Google Scholar] [CrossRef]
- Redpath, S.M. Impact of habitat fragmentation on activity and hunting behavior in the tawny owl, Strix aluco. Behav. Ecol. 1995, 6, 410–413. [Google Scholar] [CrossRef] [Green Version]
- Vilella, M.; Ferrandiz-Rovira, M.; Sayol, F. Coexistence of predators in time: Effects of season and prey availability on species activity within a Mediterranean carnivore guild. Ecol. Evol. 2020, 10, 11408–11422. [Google Scholar] [CrossRef]
- Díaz, M.; Torre, I.; Peris, A.; Tena, L. Foraging behavior of wood mice as related to presence and activity of genets. J. Mammal. 2005, 86, 1178–1185. [Google Scholar] [CrossRef]
- Zmihorski, M.; Gryz, J.; Krauze-Gryz, D.; Olczyk, A.; Osojca, G. The tawny owl Strix aluco as a material collector in faunistic investigations: The case study of small mammals in NE Poland. Acta Zool. Litu. 2011, 21, 185–191. [Google Scholar] [CrossRef]
- Grajera, J.; Vilella, M.; Torre, I. A pilot study of the use of dry dog food as an alternative attractant in mesocarnivore studies. Mammalia 2021, 85, 422–427. [Google Scholar] [CrossRef]
- Krebs, C.J. Population dynamics of large and small mammals: Graeme Caughley’s grand vision. Wildl. Res. 2009, 36, 1–7. [Google Scholar] [CrossRef]
- Morán-López, T.; Wiegand, T.; Morales, J.M.; Valladares, F.; Díaz, M. Predicting forest management effects on oak–rodent mutualisms. Oikos 2016, 125, 1445–1457. [Google Scholar] [CrossRef] [Green Version]
- Garden, J.G.; Mcalpine, C.A.; Possingham, H.P.; Jones, D.N. Habitat structure is more important than vegetation composition for local-level management of native terrestrial reptile and small mammal species living in urban remnants: A case study from Brisbane, Australia. Austral. Ecol. 2007, 32, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; Sánchez-Mejía, T.; Morán-López, T. Long-Term Tree Regeneration of Fragmented Agroforestry Systems Under Varying Climatic Conditions. Front. Ecol. Evol. 2021, 9, 96. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VARIABLES | C. Russula | A. Sylvaticus | M. Spretus | TOTAL | Richness | CV–TOTAL |
|---|---|---|---|---|---|---|
| (Intercept) | 1.84 ** | 14.13 *** | 1.33 * | 17.91 *** | 1.79 *** | 0.75 *** |
| (0.65) | (1.28) | (0.55) | (1.67) | (0.17) | (0.06) | |
| LiDAR-PC1 | −0.91 *** | −2.21 *** | −0.68 ** | −3.64 *** | −0.18 *** | 0.07 ** |
| (0.21) | (0.46) | (0.20) | (0.60) | (0.04) | (0.03) | |
| LiDAR-PC2 | 1.81 | 1.88 | ||||
| (0.91) | (1.19) | |||||
| Season (Autumn) | 1.35 * | −10.79 *** | −9.79 *** | |||
| (0.64) | (1.81) | (1.99) | ||||
| PC1 X Season | 2.07 ** | 1.82* | ||||
| (0.65) | (0.71) | |||||
| PC2 X Season | −2.26 | −1.86 | ||||
| (1.29) | (1.41) | |||||
| Log Likelihood | −76.93 | −95.36 | −68.31 | −102.10 | −24.70 | −8.95 |
| AICc | 165.99 | 212.48 | 146.01 | 225.95 | 58.78 | 27.56 |
| Delta | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.40 |
| Weight | 0.70 | 0.48 | 1.00 | 0.40 | 1.00 | 1.00 |
| R2 marginal | 0.49 | 0.67 | 0.38 | 0.67 | 0.41 | 0.26 |
| R2 conditional | 0.73 | 0.67 | 0.86 | 0.77 | 0.76 | 0.48 |
| Num. obs. | 34 | 34 | 34 | 34 | 34 | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torre, I.; Jaime-González, C.; Díaz, M. Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter. Sustainability 2022, 14, 1562. https://doi.org/10.3390/su14031562
Torre I, Jaime-González C, Díaz M. Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter. Sustainability. 2022; 14(3):1562. https://doi.org/10.3390/su14031562
Chicago/Turabian StyleTorre, Ignasi, Carlos Jaime-González, and Mario Díaz. 2022. "Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter" Sustainability 14, no. 3: 1562. https://doi.org/10.3390/su14031562
APA StyleTorre, I., Jaime-González, C., & Díaz, M. (2022). Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter. Sustainability, 14(3), 1562. https://doi.org/10.3390/su14031562