
Nutrient Remediation Efficiency of the Sedge Plant (Cyperus alopecuroides Rottb.) to Restore Eutrophic Freshwater Ecosystems


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Design and Growth Measurements
2.2. Plant Analysis
2.2.1. Inorganic Nutrients
2.2.2. Organic Nutrients
2.3. Sediments and Water Analysis
2.4. Data Analysis
3. Results
3.1. Sediment and Water Properties
3.2. Growth Properties
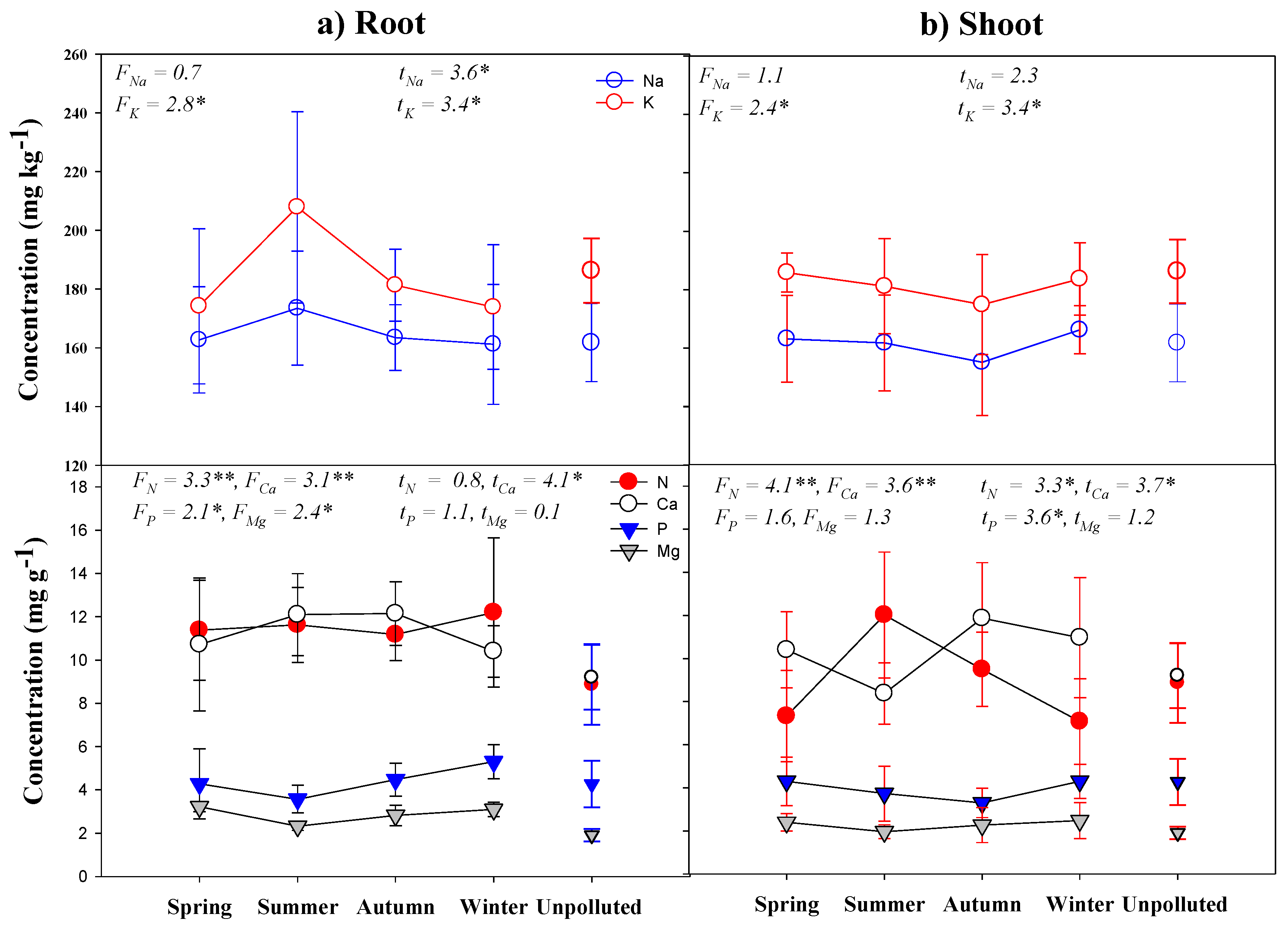
3.3. Inorganic Nutrient Concentrations
3.4. Nutrient’s Removal Efficiency
3.5. Organic Nutrient Content
3.6. Forage Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yaqub, M.; Asif, H.; Kim, S.; Lee, W. Modeling of a full-scale sewage treatment plant to predict the nutrient removal efficiency using a long short-term memory (LSTM) neural n. J. Water Process Eng. 2020, 37, 101388. [Google Scholar] [CrossRef]
- Krimsky, L.S.; Lusk, M.G.; Abeels, H.; Seals, L. Sources and concentrations of nutrients in surface runoff from waterfront homes with different landscape practices. Sci. Total Environ. 2021, 750, 142320. [Google Scholar] [CrossRef] [PubMed]
- Zainol, Z.; Fadzil, M.; Abdullah, S. Hydrodynamics, nutrient concentrations, and phytoplankton biomass in a shallow and restricted coastal lagoon under different tidal and monsoonal environmental drivers. Reg. Stud. Mar. Sci. 2020, 38, 101376. [Google Scholar] [CrossRef]
- Jani, J.; Yang, Y.Y.; Lusk, M.G.; Toor, G.S. Composition of nitrogen in urban residential stormwater runoff: Concentrations, loads, and source characterization of nitrate and organic nitrogen. PLoS ONE 2020, 15, e0229715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, P.; Serra, I.; Townley, B.; Aburto, F.; López, S.; Tapia, J. Biogeochemistry of plant essential mineral nutrients across rock, soil, water and fruits in Vineyards of Central Chile. Catena 2021, 196, 104905. [Google Scholar] [CrossRef]
- Huang, J.; Chen, Q.; Peng, J.; Gao, J. Quantifying the cost-effectiveness of nutrient-removal strategies for a lowland rural watershed: Insights from process-based modeling. Ecol. Model. 2020, 431, 109123. [Google Scholar] [CrossRef]
- Quilliam, R.S.; van Niekerk, M.A.; Chadwick, D.R.; Cross, P.; Hanley, N.; Jones, D.L.; Vinten, A.J.A.; Willby, N.; Oliver, D.M. Can macrophyte harvesting from eutrophic water close the loop on nutrient loss from agricultural land? J. Environ. Manag. 2015, 152, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Peter, T.; Guo, H.; Huang, Z.; Steven, K.; Wang, H.; Gu, M.; Wei, L. Variation of dissolved nutrient exports by surface runoff from sugarcane watershed is controlled by fertilizer application and ground cover. Agric. Ecosyst. Environ. 2020, 303, 107121. [Google Scholar] [CrossRef]
- van der Wiel, B.Z.; Weijma, J.; Van Middelaar, C.E.; Kleinke, M.; Jan, C.; Buisman, N.; Wichern, F. Restoring nutrient circularity: A review of nutrient stock and flow analyses of local agro-food-waste systems. Resour. Conserv. Recycl. X 2020, 160, 104901. [Google Scholar] [CrossRef]
- Fan, H.; Wang, K.; Wang, C.; Yu, F.; He, X. A comparative study on growth characters and nutrients removal from wastewater by two microalgae under optimized light regimes. Environ. Technol. Innov. 2020, 19, 100849. [Google Scholar] [CrossRef]
- Kiani, M.; Tammeorg, P.; Niemistö, J.; Simojoki, A.; Tammeorg, O. Internal phosphorus loading in a small shallow lake: Response after sediment removal. Sci. Total Environ. 2020, 725, 138279. [Google Scholar] [CrossRef] [PubMed]
- Taillardat, P.; Marchand, C.; Friess, D.A.; Widory, D.; David, F.; Ohte, N.; Nakamura, T.; Van Vinh, T.; Thanh-Nho, N.; Ziegler, A.D. Respective contribution of urban wastewater and mangroves on nutrient dynamics in a tropical estuary during the monsoon season. Mar. Pollut. Bull. 2020, 160, 111652. [Google Scholar] [CrossRef] [PubMed]
- Mosley, L.M. Drought impacts on the water quality of freshwater systems; review and integration. Earth Sci. Rev. 2015, 140, 203–214. [Google Scholar] [CrossRef]
- Geurts, J.J.; Oehmke, C.; Lambertini, C.; Eller, F.; Sorrell, B.K.; Mandiola, S.R.; Grootjans, A.P.; Brix, H.; Wichtmann, W.; Lamers, L.P.; et al. Nutrient removal potential and biomass production by Phragmites australis and Typha latifolia on European rewetted peat and mineral soils. Sci. Total Environ. 2020, 747, 141102. [Google Scholar] [CrossRef]
- Kumwimba, M.N.; Zhud, B.; Wang, T.; Dzakpasu, M.; Li, X. Nutrient dynamics and retention in a vegetated drainage ditch receiving nutrient-rich sewage at low temperatures. Sci. Total Environ. 2020, 741, 140268. [Google Scholar] [CrossRef]
- Vymazal, J. Removal of nutrients in constructed wetlands for wastewater treatment through plant harvesting—Biomass and load matter the most. Ecol. Eng. 2020, 155, 105962. [Google Scholar] [CrossRef]
- Sutherland, D.L.; Burke, J.; Ralph, P.J. Increased harvest frequency improves biomass yields and nutrient removal on a filamentous algae nutrient scrubber. Algal Res. 2020, 51, 102073. [Google Scholar] [CrossRef]
- Manolaki, P.; Mouridsen, M.B.; Nielsen, E.; Olesen, A.; Jensen, S.M.; Lauridsen, T.L.; Baattrup-Pedersen, A.; Sorrell, B.K.; Riis, T. A comparison of nutrient uptake efficiency and growth rate between different macrophyte growth forms. J. Environ. Manag. 2020, 274, 111181. [Google Scholar] [CrossRef]
- Meuleman, A.F.M.; Beekman, P.; Verhoeven, J.T.A. Nutrient retention and nutrient-use efficiency in Phragmites australis stands after wastewater application. Wetlands 2002, 22, 712–721. [Google Scholar] [CrossRef]
- Plaimart, J.; Acharya, K.; Mrozik, W.; Davenport, R.J.; Vinitnantharat, S.; Werner, D. Coconut husk biochar amendment enhances nutrient retention by suppressing nitrification in agricultural soil following anaerobic digestate application. Environ. Pollut. 2021, 268, 115684. [Google Scholar] [CrossRef]
- Tanaka, T.S.; Irbis, C.; Kumagai, H.; Wang, P.; Li, K.; Inamura, T. Effect of Phragmites japonicus harvest frequency and timing on dry matter yield and nutritive value. J. Environ. Manag. 2017, 187, 436–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruinsma, J. World Agriculture: Towards 2015/2030, an FAO Perspective; Earthscan, FAO: Rome, Italy, 2003. [Google Scholar]
- Rattanasomboon, T.; Wester, T.G.; Smith, S.L.; Morel, P.C. Nutritive value of plantain and chicory for pigs. Livest. Sci. 2019, 223, 8–15. [Google Scholar] [CrossRef]
- Galal, T.M.; Shedeed, Z.A.; Gharib, F.A.; Al-Yasi, H.M.; Mansour, K.H. The role of Cyperus alopecuroides Rottb. sedge in monitoring water pollution in contaminated wetlands in Egypt: A phytoremediation approach. Environ. Sci. Pollut. Res. 2021, 28, 23005–23016. [Google Scholar] [CrossRef]
- Kantor, H.J. Chapter V: Cyperus alopecuroides Rottb. and the Egyptian volute. In Plant Ornament in the Ancient Near East; Oriental Institute, University of Chicago: Chicago, IL, USA, 1999; pp. 169–198. [Google Scholar]
- Nawaz, T.; Hameed, M.; Ashraf, M.; Sajid, M.; Ahmad, A.; Batool, R.; Fatima, S. Anatomical and physiological adaptations in aquatic ecotypes of Cyperus alopecuroides Rottb. under saline and waterlogged conditions. Aquat. Bot. 2014, 116, 60–68. [Google Scholar] [CrossRef]
- Boulos, L. Medicinal Plants of North Africa; Reference Publications: Algonac, MI, USA, 1983. [Google Scholar]
- Sampat, A.M.; Hicks, A.; Ruiz-mercado, G.J.; Zavala, V.M. Valuing economic impact reductions of nutrient pollution from livestock waste. Resour. Conserv. Recycl. 2021, 164, 105199. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; Al-Sodany, Y.M.; Haroun, S.A.; Galal, T.M.; Ayed, H.; Khedher, K.M.; Jensen, K. Seasonal potential of Phragmites australis in nutrient removal to eliminate the eutrophication in Lake Burullus, Egypt. J. Freshw. Ecol. 2020, 35, 135–155. [Google Scholar] [CrossRef] [Green Version]
- Batool, R.; Hameed, M.; Ashraf, M.; Fatima, S.; Nawaz, T.; Ahmad, M.S.A. Structural and functional response to metal toxicity in aquatic Cyperus alopecuroides Rottb. Limnologica 2014, 48, 46–56. [Google Scholar] [CrossRef]
- Ghazi, S.M.; Galal, T.M.; Hussein, K.H. Monitoring Water Pollution in the Egyptian Watercourses: A Phytoremediation Approach; Lap Lambert Academic Publishing: Saarbrucken, Germany, 2019; p. 153. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: London, UK, 1989. [Google Scholar]
- Adesogon, A.T.; Givens, D.I.; Owen, E. Measuring chemical composition and nutritive value in forages. In Field and Laboratory Methods for Grassland and Animal Production Research; t’Mannetje, L., Jones, R.M., Eds.; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Le Houérou, H.N. Chemical composition and nutritive value of browse in Tropical West Africa. In Browse in Africa: The Current State of Knowledge; Le Houérou, H.N., Ed.; ILCA: Addis Ababa, Ethiopia, 1980; pp. 261–289. [Google Scholar]
- Demarquilly, C.; Weiss, P. Tableau de la Valeur Alimentaire des Fourrages. Et. 42: Versailles INRA-SEI; INRA: Paris, France, 1970. [Google Scholar]
- Naga, M.A.; El-Shazly, K. The prediction of the nutritive value of animal feeds from chemical analysis. J. Agric. Sci. 1971, 77, 25–31. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Domestic Animals: Nutrient Requirement of Beef Cattle, 6th ed.; National Research Council No. 5; National Academic Sciences: Washington, DC, USA, 1984. [Google Scholar]
- Garrett, W.N. Energy utilization of growing cattle as determined in seventy-two comparative laughter experiments. In Energy Metabolism; No. 26; Mount, L.E., Ed.; EAAP Publication: London, UK, 1980. [Google Scholar]
- SPSS. IBM SPSS Statistics Version 21.0. Copyright of IBM and Other(s) 1989–2012; SPSS: Chicago, IL, USA, 2012. [Google Scholar]
- Galal, T.M.; Gharib, F.A.; Ghazi, S.M.; Mansour, K.H. Metal uptake capability of Cyperus articulatus L. and its role in mitigating heavy metals from contaminated wetlands. Environ. Sci. Pollut. Res. 2017, 24, 21636–21648. [Google Scholar] [CrossRef]
- Galal, T.M.; Gharib, F.A.; Al-Yasi, H.M.; Mansour, K.H.; Hassan, M.M. Evaluation of the nutrient status and forage quality of the hippo grass (Vossia cuspidata (Roxb.) Griff.) along Ismailia canal, Egypt. J. Freshw. Ecol. 2021, 36, 63–76. [Google Scholar] [CrossRef]
- Wu, N.; Liu, S.; Zhang, G.; Zhang, H. Anthropogenic impacts on nutrient variability in the lower Yellow River. Sci. Total Environ. 2021, 755, 142488. [Google Scholar] [CrossRef] [PubMed]
- Galal, T.M.; Abu Alhmad, M.F.; Al-Yasi, H.M. Nutrient sequestration potential of water primrose Ludwigia stolinefera (Guill. & Perr.) PH Raven: A strategy for restoring wetland eutrophication. Saudi J. Biol. Sci. 2021, 28, 2438–2446. [Google Scholar] [PubMed]
- Saleh, H.M.; Aglan, R.F.; Mahmoud, H.H. Ludwigia stolonifera for remediation of toxic metals from simulated wastewater. Chem. Ecol. 2019, 35, 164–178. [Google Scholar] [CrossRef]
- Galal, T.M.; Shehata, H.S. Growth and nutrients accumulation potentials of giant reed (Arundo donax L.) in different habitats in Egypt. Int. J. Phytoremediation 2016, 18, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Eid, E.M.; Shaltout, K.H.; El-Sheikh, M.A.; Asaeda, T. Seasonal courses of nutrients and heavy metals in water, sediment and above- and below-ground Typha domingensis biomass in Lake Burullus (Egypt): Perspective for phytoremediation. Flora 2012, 207, 783–794. [Google Scholar] [CrossRef]
- Soda, S.; Hamada, T.; Yamaoka, Y.; Ike, M.; Nakazato, H.; Saeki, Y.; Kasamatsu, T.; Sakurai, Y. Constructed wetlands for advanced treatment of wastewater with a complex matrix from a metal-processing plant: Bioconcentration and translocation factors of various metals in Acorus gramineus and Cyperus alternifolius. Ecol. Eng. 2012, 39, 63–70. [Google Scholar] [CrossRef]
- Caselles-osorio, A.; Vega, H.; Camilo, J.; Casierra-martínez, H.A.; Euliser, J. Horizontal subsurface-flow constructed wetland removal efficiency using Cyperus articulatus L. Ecol. Eng. 2017, 99, 479–485. [Google Scholar] [CrossRef]
- Klaus, V.H.; Kleinebecker, T.; Hölzel, N.; Blüthgen, N.; Boch, S.; Müller, J.; Socherc, S.A.; Prati, D.; Fischer, M. Nutrient concentrations and fiber contents of plant community biomass reflect species richness patterns along a broad range of land-use intensities among agricultural grasslands. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 287–295. [Google Scholar] [CrossRef]
- Vymazal, J.; Richardson, C.J. The relationship between soil, water nutrients and biomass of Cladium jamaicense and Typha domingensis in the northern Everglades. In Wetlands—Nutrients, Metals and Mass Cycling; Vymazal, J., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 107–127. [Google Scholar]
- Zhao, Y.; Xia, X.; Yang, Z. Growth and nutrient accumulation of Phragmites australis in relation to water level variation and nutrient loadings in a shallow lake. J. Environ. Sci. 2013, 25, 16–25. [Google Scholar] [CrossRef]
- Vymazal, J. Nitrogen removal in constructed wetlands with horizontal sub-surface flow—Can we determine the key process? In Nutrient Cycling and Retention in Natural and Constructed Wetlands; Vymazal, J., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1999; pp. 1–17. [Google Scholar]
- Kasak, K.; Valach, A.C.; Rey-Sanchez, C.; Kill, K.; Shortt, R.; Liu, J.; Dronova, I.; Mander, Ü.; Szutu, D.; Verfaillie, J.; et al. Experimental harvesting of wetland plants to evaluate trade-offs between reducing methane emissions and removing nutrients accumulated to the biomass in constructed wetlands. Sci. Total Environ. 2020, 715, 136960. [Google Scholar] [CrossRef]
- Mulderij, G.; Mau, B.; Domis, L.N.; Smolders, A.J.P.; Van Donk, E. Interaction between the macrophyte Stratiotes aloides and filamentous algae: Does it indicate allelopathy? Aquat. Ecol. 2009, 43, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, T.R.; Gill, R.S.; Ichhponani, J.S. Nutritive value of berseem and cluster bean forages. Indian J. Anim. Sci. 1980, 50, 1052–1055. [Google Scholar]
- El-Kady, H. Seasonal variation in phytomass and nutrient status of Phragmites australis along the water courses in the Middle Delta region. Taeckholmia 2002, 20, 123–138. [Google Scholar] [CrossRef]
- Shoukry, M.M. An actual vision about the availability of the utilization of water hyacinth in feeding ruminants. In Proceedings of the National Symposium on Water Hyacinth, Assiut University, Assiut, Egypt, 25–26 February 1992; pp. 75–92. [Google Scholar]
- El-Beheiry, M.A. Nutritive evaluation of two Acacia populations in Southwestern Saudi Arabia. J. Appl. Sci. Res. 2009, 5, 1030–1039. [Google Scholar]
- NRC. Nutrient Requirements of Domestic Animals: Nutrient Requirement of Sheep, 6th ed.; National Research Council Pamphelets No. 5; National Academic Sciences: Washington, DC, USA, 1985. [Google Scholar]
- NRC. Nutrient Requirements of Domestic Animals: Nutrient Requirement of Dairy Cattle, 5th ed.; National Research Council: Washington, DC, USA, 1978. [Google Scholar]
- NRC. Nutrient Requirements of Domestic Animals: Nutrient Requirement of Goats; National Research Council No. 15; National Academy of Science: Washington, DC, USA, 1981. [Google Scholar]
- NRC. Nutrient Requirements of Domestic Animals: Nutrient Requirement of Sheep, 5th ed.; Natlional Research Council: Washington, DC, USA, 1975. [Google Scholar]
- Ray, P.; Chattaraj, S.; Bandyopadhyay, S.; Jena, R.K.; Singh, S.K.; Ray, S.K. Shifting cultivation, soil degradation and agricultural land use planning in the Northeastern hill region of India using geo-spatial techniques. Land Degrad. Dev. 2021, 32, 3856–3869. [Google Scholar] [CrossRef]
- Kumar, A.; Jigyasu, D.K.; Kumar, A.; Subrahmanyam, G.; Mondal, R.; Shabnam, A.A.; Cabral-Pinto, M.M.S.; Malyan, S.K.; Chaturvedi, A.K.; Gupta, D.K.; et al. Nickel in terrestrial biota: Comprehensive review on contamination, toxicity, tolerance and its remediation approaches. Chemosphere 2021, 275, 129996. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Water (mg/L) | t-Value | Sediment (mg/kg) | t-Value | ||
|---|---|---|---|---|---|---|
| Unpolluted | Polluted | Unpolluted | Polluted | |||
| pH | 6.1 ± 0.8 | 7.4 ± 0.8 | 2.7 * | 7.3 ± 0.4 | 5.4 ± 0.8 | 2.6 * |
| EC (µS/cm) | 242.3 ± 10.2 | 456.6 ± 9.6 | 5.8 ** | 372.7 ± 32.7 | 476.4 ± 24.6 | 2.8 * |
| CO3 | 364.3 ± 63.6 | 401.1 ± 54.6 | 1.1 | 254.2 ± 2.2 | 392.0 ± 5.8 | 3.1 * |
| HCO3 | 396.2 ± 18.4 | 408.1 ± 21.4 | 1.2 | 241.9 ± 21.7 | 396.5 ± 13.1 | 3.4 * |
| SO4 | 78.1 ± 9.9 | 106.1 ± 15.6 | 2.5 * | 251.3 ± 9.8 | 350.0 ± 8.1 | 3.6 * |
| N | 31.2 ± 6.4 | 52.2 ± 6.7 | 2.6 * | 132.4 ± 12.3 | 221.3 ± 12.8 | 4.7 ** |
| P | 183.4 ± 32.1 | 424.1 ± 31.5 | 6.2 ** | 81.8 ± 2.1 | 92.9 ± 6.1 | 2.6 * |
| Na | 48.6 ± 7.3 | 94.2 ± 11.2 | 3.4 * | 142.7 ± 6.2 | 213.5 ± 6.7 | 2.8 * |
| K | 211.7 ± 21.4 | 221.3 ± 43.6 | 1.1 | 124.6 ± 6.9 | 143.8 ± 10.8 | 4.2 ** |
| Cl | 14.2 ± 2.4 | 20.2 ± 3.6 | 2.6 * | 156.2 ± 11.2 | 221.3 ± 12.2 | 4.6 ** |
| Season | Organic Nutrient (%) | ||||||
|---|---|---|---|---|---|---|---|
| EE | CF | Ash | TP | NFE | |||
| Polluted locations | Spring | R | 1.1 ± 0.3a | 34.9 ± 5.5bc | 7.7 ± 0.6b | 8.4 ± 0.5a | 48.0 ± 5.4bc |
| S | 0.8 ± 0.1ab | 42.9 ± 8.2b | 7.8 ± 1.2b | 5.7 ± 0.4b | 42.8 ± 7.2bc | ||
| Summer | R | 0.4 ± 0.1d | 7.9 ± 2.8e | 11.2 ± 1.3a | 7.3 ± 1.6ab | 73.1 ± 14.4a | |
| S | 0.7 ± 0.1b | 50.7 ± 2.2ab | 8.7 ± 0.8ab | 5.5 ± 0.3b | 34.4 ± 2.7bc | ||
| Autumn | R | 0.7 ± 0.1b | 16.8 ± 4.5de | 11.3 ± 1.2a | 6.4 ± 0.7ab | 64.8 ± 15.1ab | |
| S | 0.6 ± 0.2bc | 38.6 ± 13.3bc | 9.8 ± 1.1ab | 6.7 ± 0.5ab | 44.3 ± 2.0bc | ||
| Winter | R | 0.5 ± 0.1c | 20.3 ± 7.2d | 9.7 ± 0.6ab | 8.8 ± 2.4a | 60.7 ± 11.3b | |
| S | 0.8 ± 0.3ab | 55.4 ± 7.8a | 9.5 ± 0.8ab | 5.4 ± 0.3b | 28.9 ± 7.8c | ||
| F-value | 3.2 ** | 2.6 * | 2.7 * | 3.3 ** | 2.6 * | ||
| Organic Nutrient | Root System | t-Value | Shoot System | t-Value | ||
|---|---|---|---|---|---|---|
| P | U | P | U | |||
| EE | 0.7 ± 0.3 | 0.5 ± 0.1 | 1.2 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.1 |
| CF | 20.0 ± 11.2 | 34.3 ± 9.9 | 4.1 * | 46.9 ± 7.5 | 40.5 ± 7.1 | 2.4 * |
| Ash | 10.1 ± 1.7 | 10.9 ± 1.6 | 1.1 | 9.2 ± 0.9 | 10.1 ± 1.8 | 0.8 |
| TP | 7.7 ± 1.1 | 4.2 ± 1.1 | 2.6 * | 5.8 ± 0.6 | 6.6 ± 0.8 | 1.4 |
| NFE | 61.6 ± 10.5 | 50.2 ± 11.2 | 2.7 * | 37.6 ± 7.3 | 42.1 ± 2.6 | 3.1 * |
| Season | Nutritive Value | |||||||
|---|---|---|---|---|---|---|---|---|
| DCP | TDN | DE | ME | NE | GE | |||
| % | Mcal/kg | |||||||
| Polluted canals | Spring | R | 4.3 ± 0.5a | 57.1 ± 11.3c | 2.5 ± 0.1b | 2.1 ± 0.1bc | 1.0 ± 0.1bc | 418.7 ± 36.2ab |
| S | 1.8 ± 0.4b | 58.8 ± 13.6ab | 2.2 ± 0.2bc | 1.8 ± 0.2bc | 0.9 ± 0.1bc | 418.1 ± 29.9ab | ||
| Summer | R | 3.3 ± 1.3a | 57.5 ± 11.1c | 3.3 ± 0.9a | 2.7 ± 0.5a | 1.3 ± 0.2a | 379.1 ± 13.7c | |
| S | 1.6 ± 0.3b | 58.8 ± 12.2ab | 2.0 ± 0.c | 1.7 ± 1.1bc | 0.8 ± 0.1bc | 419.5 ± 12.8ab | ||
| Autumn | R | 2.4 ± 0.6ab | 58.3 ± 13.4ab | 2.9 ± 0.6ab | 2.4 ± 0.9ab | 1.2 ± 0.3ab | 384.8 ± 13.8abc | |
| S | 2.7 ± 0.4ab | 58.0 ± 4.7ab | 2.2 ± 0.4bc | 1.8 ± 0.6bc | 0.9 ± 0.1bc | 407.4 ± 17.2ab | ||
| Winter | R | 4.7 ± 2.2a | 56.3 ± 11.8c | 2.8 ± 0.9ab | 2.3 ± 0.4b | 1.1 ± 0.3b | 396.7 ± 15.3ab | |
| S | 1.5 ± 0.2b | 59.2 ± 12.5a | 1.9 ± 0.4cbc | 1.6 ± 0.7c | 0.8 ± 0.1c | 420.0 ± 27.6a | ||
| F-value | 3.3 ** | 3.4 ** | 2.8 ** | 2.8 ** | 2.7 * | 3.1 ** | ||
| Nutritive Value | Root System | t-Value | Shoot System | t-Value | ||
|---|---|---|---|---|---|---|
| P | U | P | U | |||
| DCP (%) | 3.7 ± 1.1 | 0.4 ± 0.1 | 3.4 * | 1.9 ± 0.5 | 2.6 ± 0.8 | 2.6 * |
| TDN (%) | 57.3 ± 8.2 | 59.7 ± 6.3 | 1.4 | 58.6 ± 5.6 | 58.1 ± 5.6 | 0.2 |
| DE (Mcal/kg) | 2.9 ± 0.3 | 2.3 ± 0.3 | 1.8 | 2.1 ± 0.7 | 2.2 ± 0.1 | 0.5 |
| ME (Mcal/kg) | 2.3 ± 0.3 | 1.9 ± 0.3 | 2.5 * | 1.7 ± 0.1 | 1.8 ± 0.1 | 0.3 |
| NE (Mcal/kg) | 1.2 ± 0.3 | 0.9 ± 0.1 | 3.1 * | 0.9 ± 0.2 | 0.9 ± 0.1 | 0.1 |
| GE (Mcal/kg) | 394.8 ± 17.5 | 395.3 ± 99.2 | 1.3 | 416.3 ± 25.9 | 407.8 ± 20.9 | 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galal, T.M.; Gharib, F.A.; Al-Yasi, H.M.; Al-Mutairi, K.A.; Mansour, K.H.; Eid, E.M. Nutrient Remediation Efficiency of the Sedge Plant (Cyperus alopecuroides Rottb.) to Restore Eutrophic Freshwater Ecosystems. Sustainability 2022, 14, 2823. https://doi.org/10.3390/su14052823
Galal TM, Gharib FA, Al-Yasi HM, Al-Mutairi KA, Mansour KH, Eid EM. Nutrient Remediation Efficiency of the Sedge Plant (Cyperus alopecuroides Rottb.) to Restore Eutrophic Freshwater Ecosystems. Sustainability. 2022; 14(5):2823. https://doi.org/10.3390/su14052823
Chicago/Turabian StyleGalal, Tarek M., Fatma A. Gharib, Hatim M. Al-Yasi, Khalid A. Al-Mutairi, Khalid H. Mansour, and Ebrahem M. Eid. 2022. "Nutrient Remediation Efficiency of the Sedge Plant (Cyperus alopecuroides Rottb.) to Restore Eutrophic Freshwater Ecosystems" Sustainability 14, no. 5: 2823. https://doi.org/10.3390/su14052823
APA StyleGalal, T. M., Gharib, F. A., Al-Yasi, H. M., Al-Mutairi, K. A., Mansour, K. H., & Eid, E. M. (2022). Nutrient Remediation Efficiency of the Sedge Plant (Cyperus alopecuroides Rottb.) to Restore Eutrophic Freshwater Ecosystems. Sustainability, 14(5), 2823. https://doi.org/10.3390/su14052823