
Climate Change in Africa and Vegetation Response: A Bibliometric and Spatially Based Information Assessment



Abstract
:1. Introduction
2. Methods
2.1. Space-Based Climate Data Analysis
2.2. Bibliometric Data and Analysis
3. Results
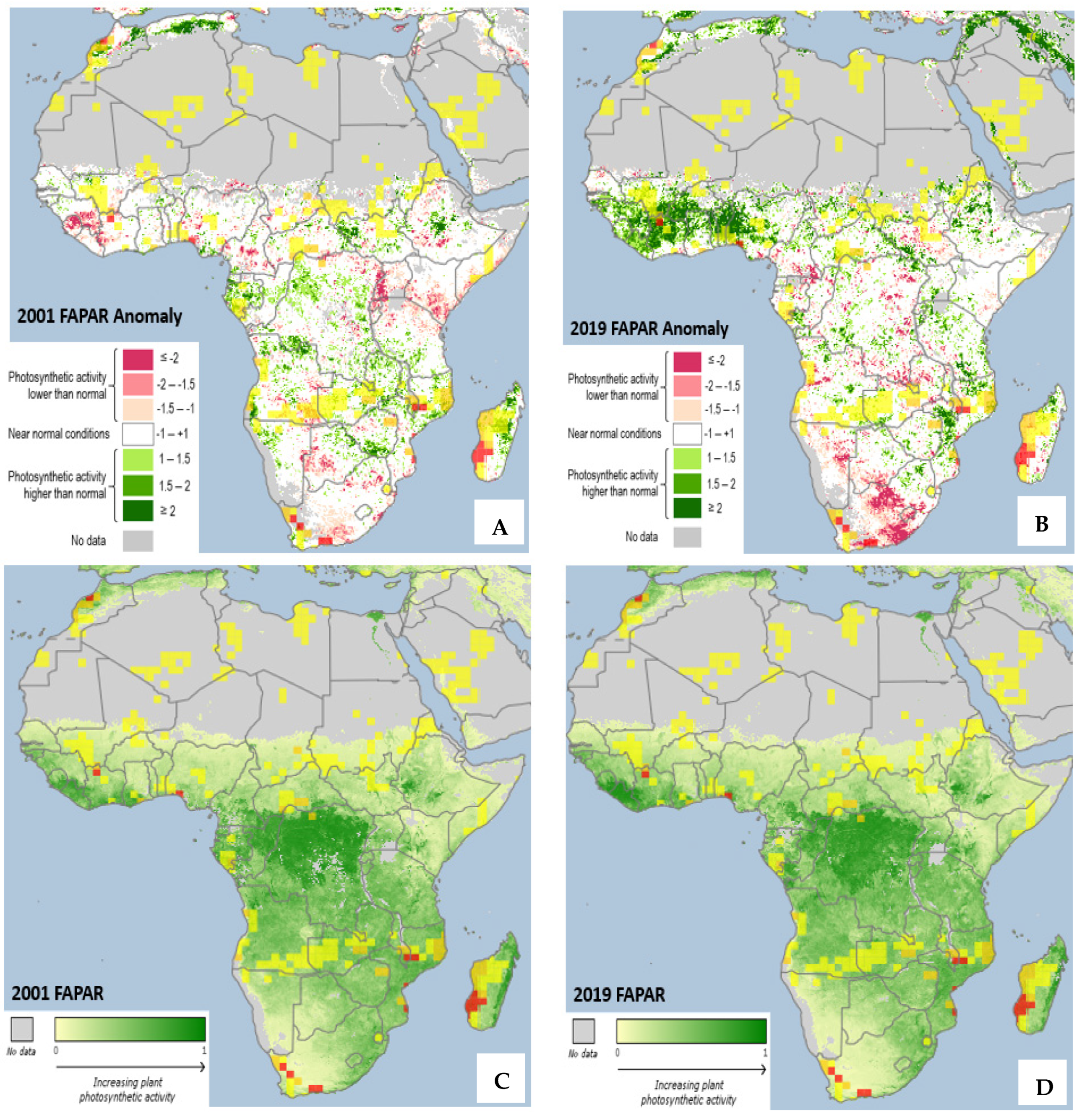
3.1. Vegetation Response to Climate Change
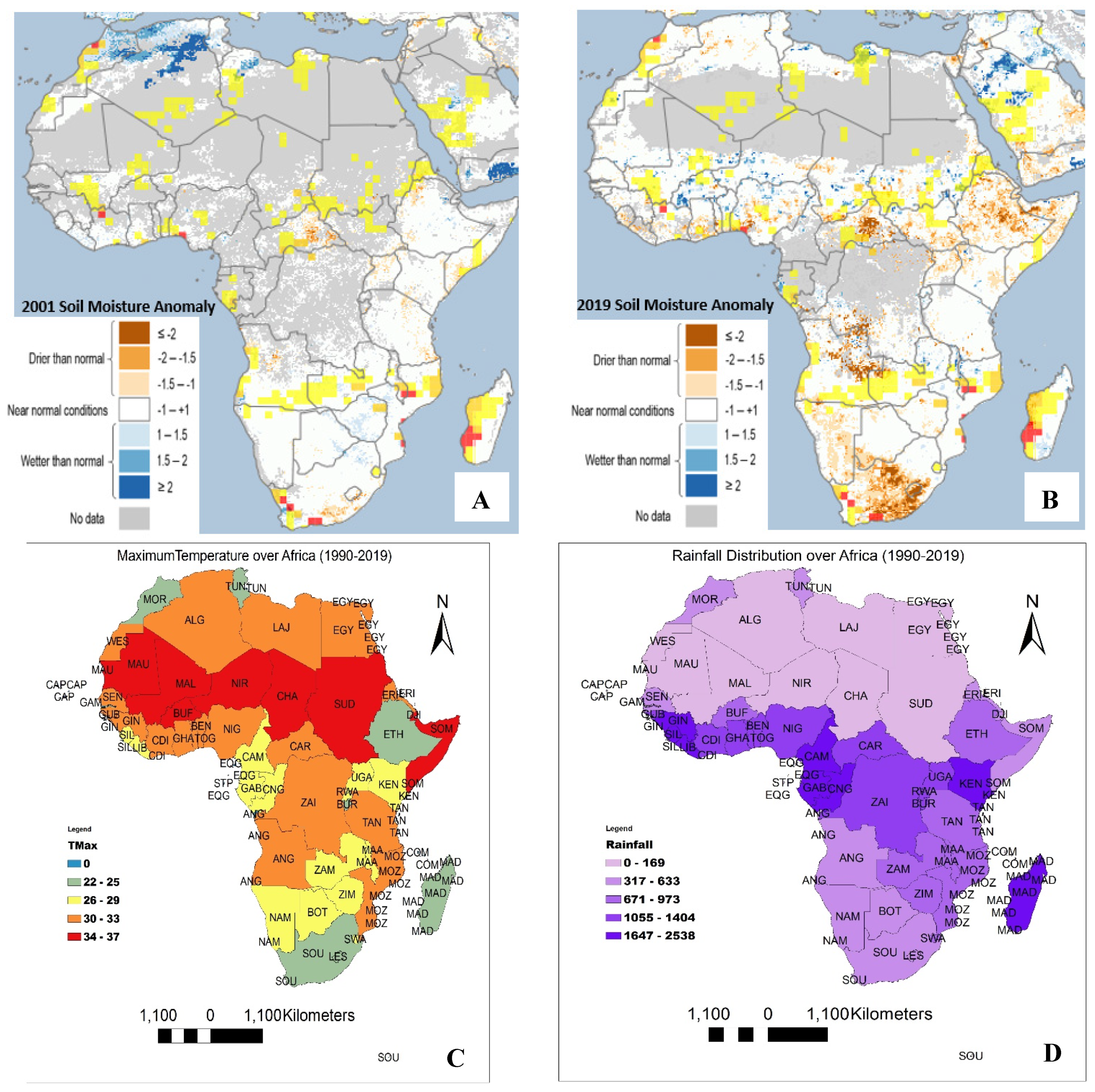
3.2. Soil Moisture Anomaly, Temperature and Rainfall Distribution
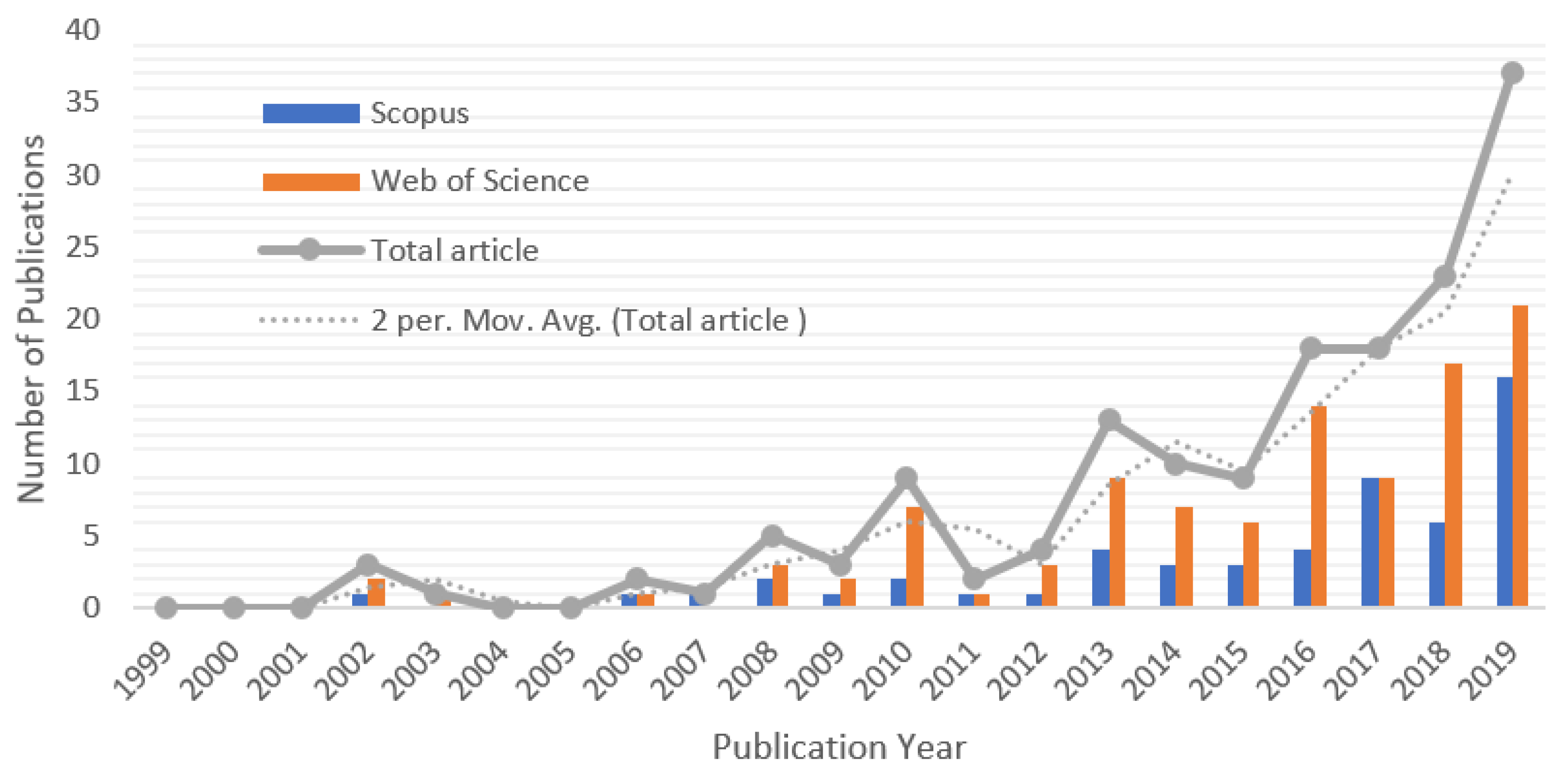
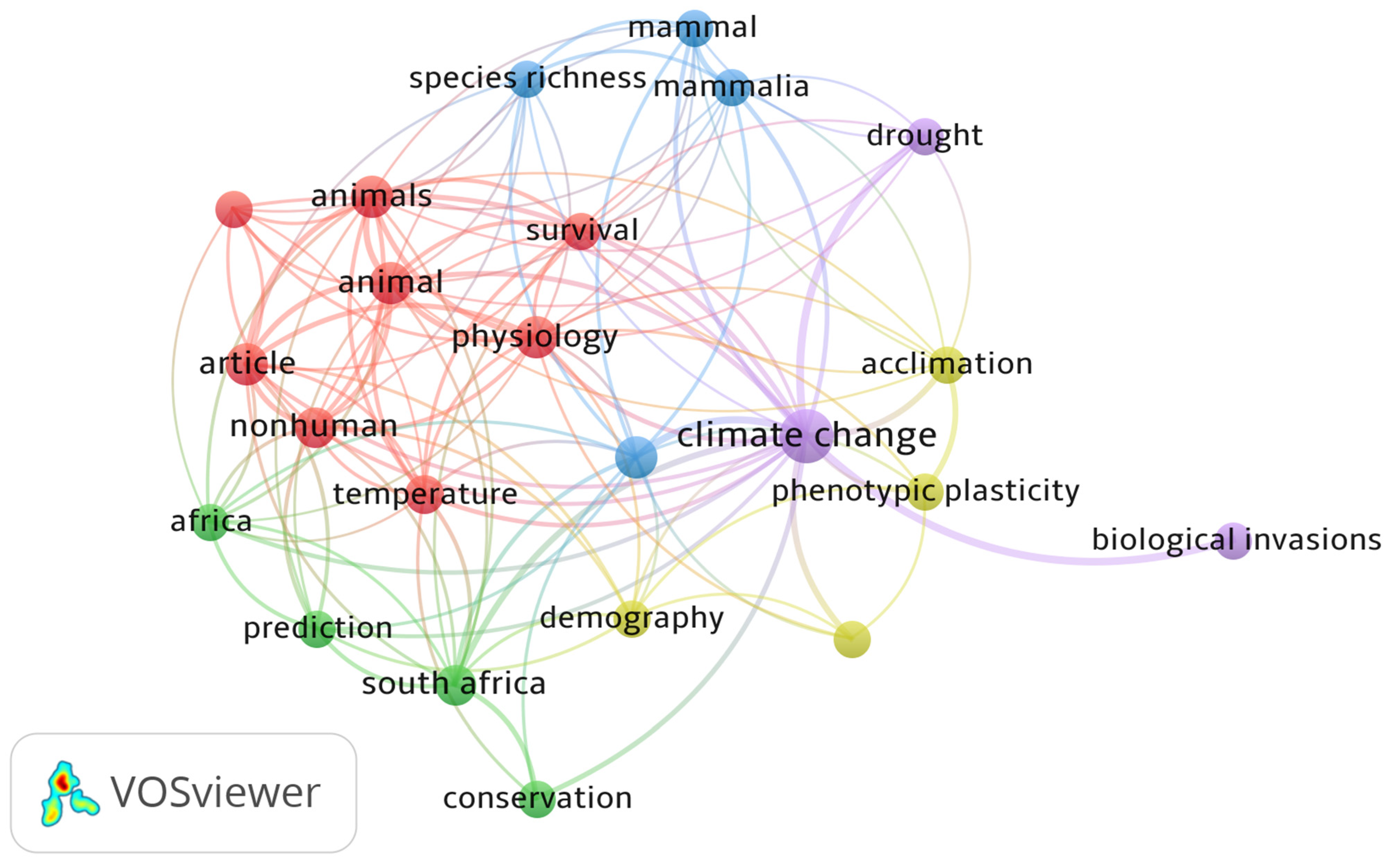
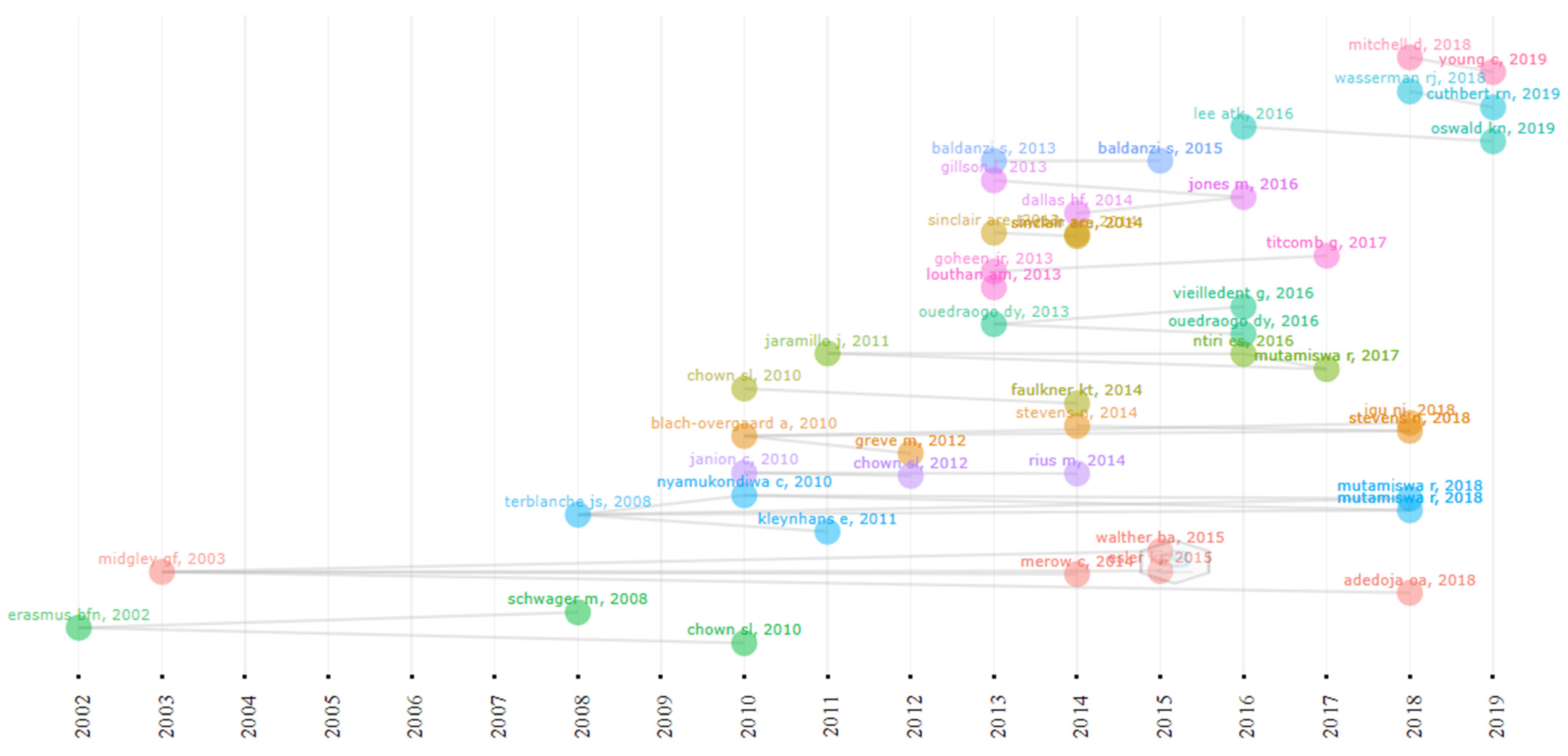
3.3. Progress of Research on African Climate Change from 1999 to 2019
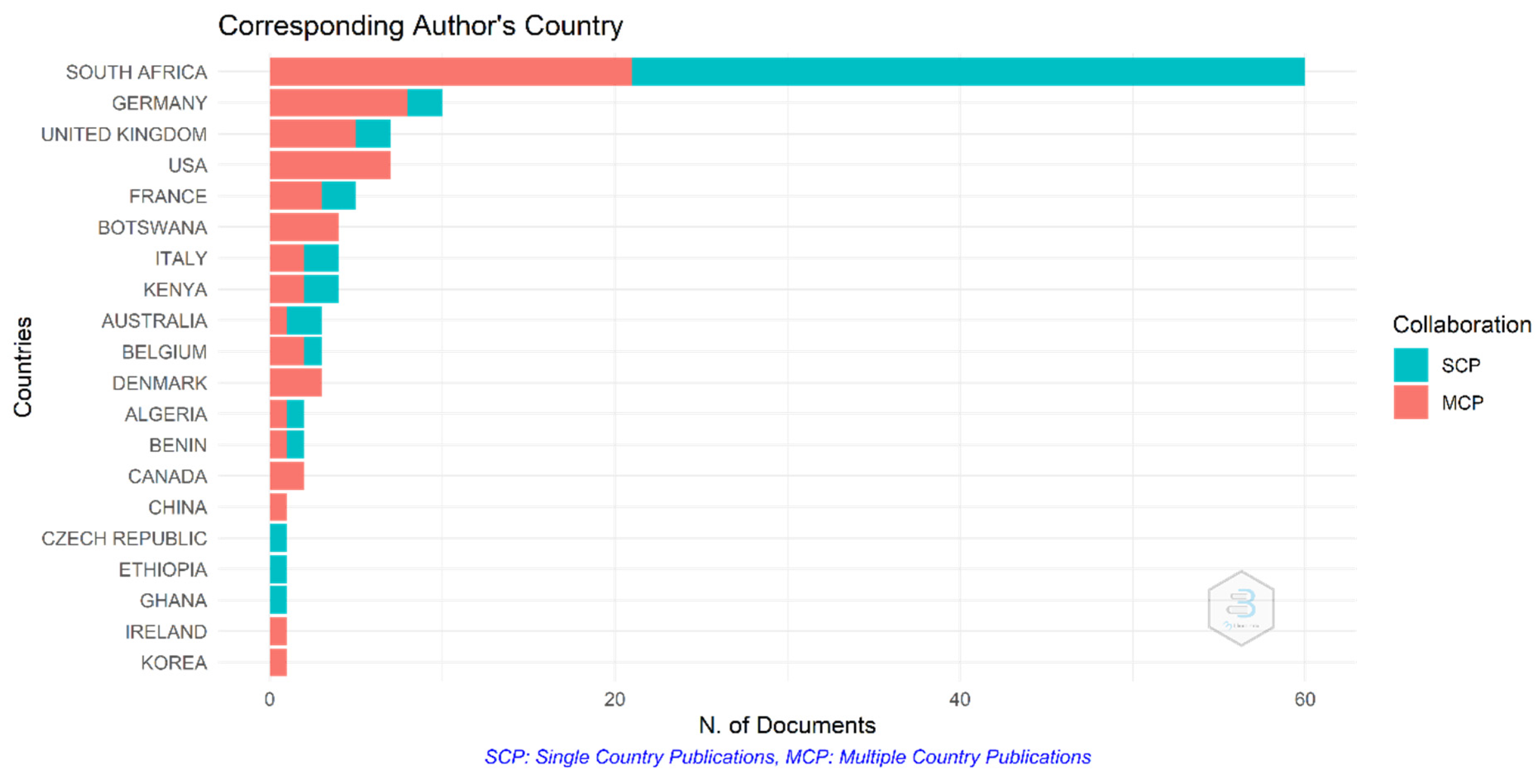
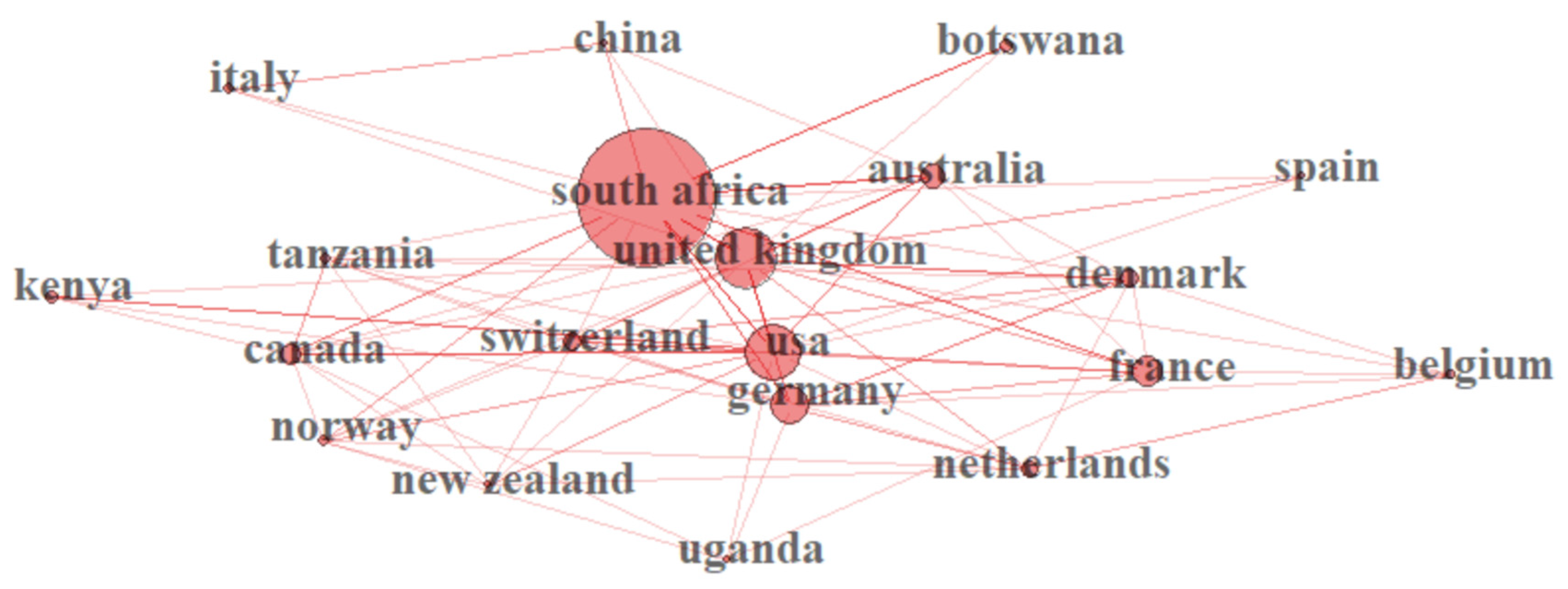
3.4. Most Productive Countries in Africa Regarding Climate Change
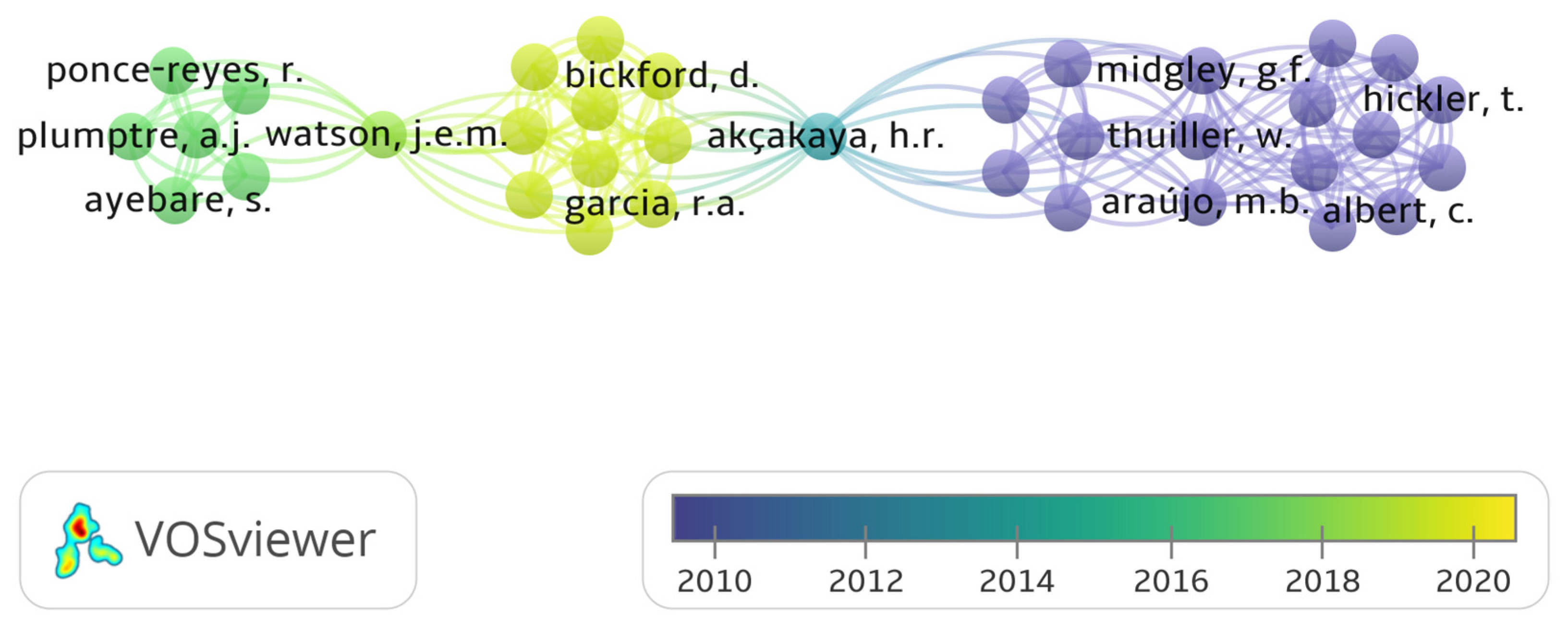
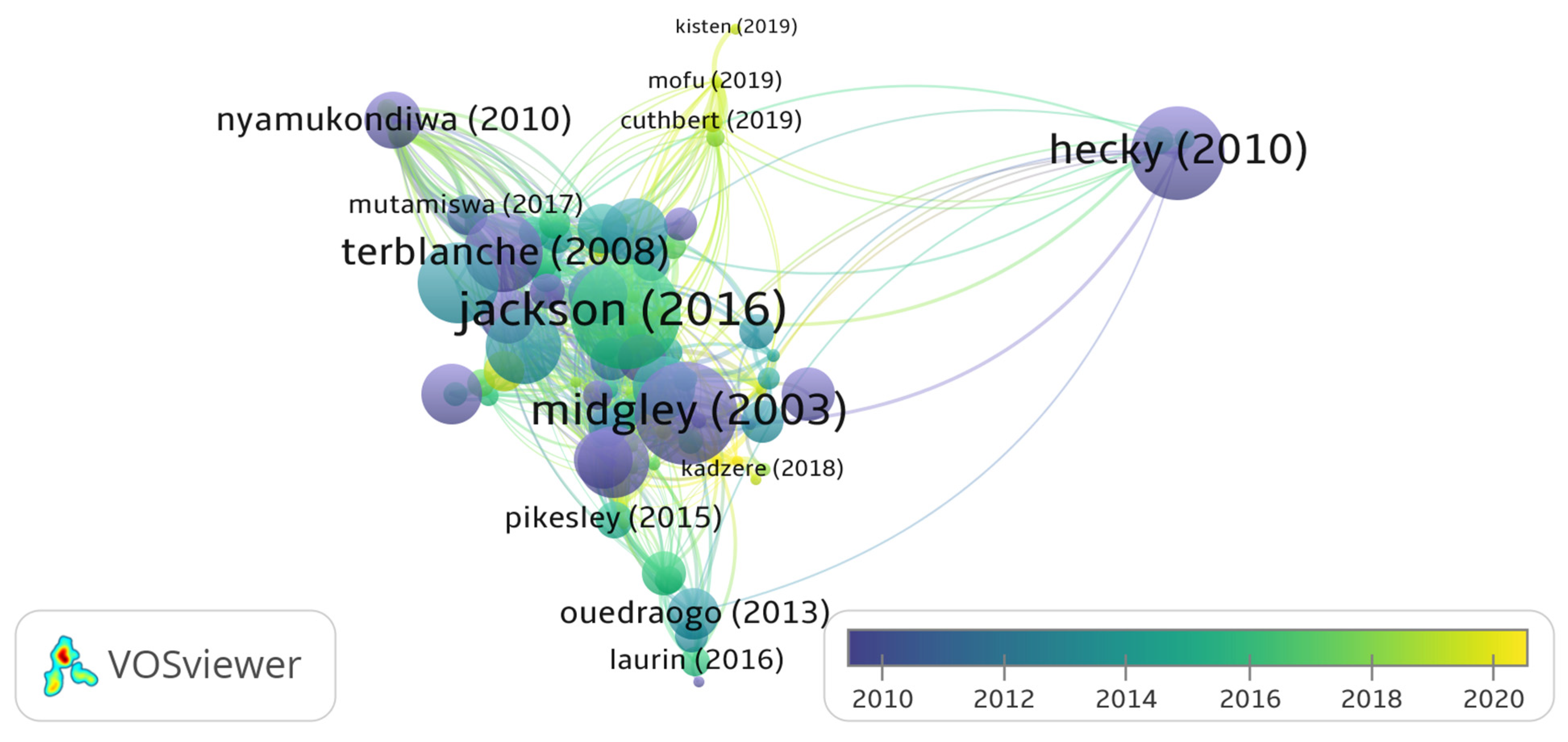
3.5. Distribution of Climate Change Publications and Citation Network
4. Responses of Climate in Fauna and Flora
4.1. Heat Stress Response
4.1.1. Heat Stress Responses at the Molecular Level: Oxidative Stress
4.1.2. Cellular and Metabolic Responses to Heat Stress
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Angert, A.L.; La Deau, S.L.; Ostfeld, R.S. Climate change and species interactions: Ways forward. Ann. N. Y. Acad. Sci. 2013, 1297, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Niles, M.T.; Mueller, N.D. Farmer perceptions of climate change: Associations with observed temperature and precipitation trends, irrigation, and climate beliefs. Glob. Environ. Chang. 2016, 39, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.; Agudo, R. The future of species under climate change: Resilience or decline? Science 2013, 341, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Vedder, O.; Bouwhuis, S.; Sheldon, B.C. Quantitative Assessment of the Importance of Phenotypic Plasticity in Adaptation to Climate Change in Wild Bird Populations. PLoS Biol. 2013, 11, 1001605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, J.S.; Hall, E.K.; Nolan, B.T.; Finlay, J.C.; Bernhardt, E.S.; Harrison, J.A.; Chan, F.; Boyer, E.W. The interactive effects of excess reactive nitrogen and climate change on aquatic ecosystems and water resources of the United States. Biogeochemistry 2013, 114, 71–92. [Google Scholar] [CrossRef]
- Salguero-GóMez, R.; Siewert, W.; Casper, B.B.; Tielbörger, K. A demographic approach to study effects of climate change in desert plants. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3100–3114. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global Change in Forests: Responses of Species, Communities, and Biomes: Interactions between climate change and land use are projected to cause large shifts in biodiversity. Bioscience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Conway, D.; Schipper, E.L.F. Adaptation to climate change in Africa: Challenges and opportunities identified from Ethiopia. Glob. Environ. Chang. 2011, 21, 227–237. [Google Scholar] [CrossRef]
- Roudier, P.; Sultan, B.; Quirion, P.; Berg, A. The impact of future climate change on West African crop yields: What does the recent literature say? Glob. Environ. Chang. 2011, 21, 1073–1083. [Google Scholar] [CrossRef] [Green Version]
- Masipa, T.S. The impact of climate change on food security in South Africa: Current realities and challenges ahead. Jamba J. Disaster Risk Stud. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Baillie, J.E.; Hilton-Taylor, C.; Stuart, S.N. 2004 IUCN Red List of Threatened Species; A Global Species Assessment IUCN: Gland, Switzerland; Cambridge, UK, 2004. [Google Scholar]
- IUCN (International Union for Conservation of Nature). Red List of Threatened Species. Available online: https://www.iucnredlist.org/about/background-history (accessed on 3 August 2020).
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Siebert, J.; Eisenhauer, N.; Schädler, M. Climate change and intensive land use reduce soil animal biomass via dissimilar pathways. Elife 2020, 9, e54749. [Google Scholar] [CrossRef]
- Personnic, S.; Boudouresque, C.F.; Astruch, P.; Ballesteros, E.; Blouet, S.; Bellan-Santini, D.; Bonhomme, P.; Thibault-Botha, D.; Feunteun, E.; Harmelin-Vivien, M.; et al. An ecosystem-based approach to assess the status of a mediterranean ecosystem, the Posidonia oceanica seagrass meadow. PLoS ONE 2014, 9, e98994. [Google Scholar] [CrossRef] [Green Version]
- Thakur, M.P.; Milcu, A.; Manning, P.; Niklaus, P.A.; Roscher, C.; Power, S.; Reich, P.B.; Scheu, S.; Tilman, D.; Ai, F.; et al. Plant diversity drives soil microbial biomass carbon in grasslands irrespective of global environmental change factors. Glob. Chang. Biol. 2015, 21, 4076–4085. [Google Scholar] [CrossRef] [Green Version]
- Batjes, N.H.; Milne, E.; Williams, S. Map-based estimates of present carbon stocks of grazing lands in Sub-Sahara Africa. In Grazing Lands, Livestock and Climate Resilient Mitigation in Sub-Saharan Africa: The State of the Science; United States Agency for International Development (USAID): Washington, DC, USA, 2015; pp. 31–33, 97–100. [Google Scholar]
- Gelaro, R.; McCarty, W.; Suárez, M.J.; Todling, R.; Molod, A.; Takacs, L.; Randles, C.A.; Darmenov, A.; Bosilovich, M.G.; Reichle, R. The Modern Era Retrospective Analysis for Research and Applications, Version 2 (MERRA-2). J. Clim. 2017, 30, 5419–5454. [Google Scholar] [CrossRef]
- Zyoud, S.H.; Waring, W.S.; Al-Jabi, S.W.; Sweileh, W.M. Global cocaine intoxication research trends during 1975-2015: A bibliometric analysis of Web of Science publications. Subst. Abus. Treat. Prev. Policy 2017, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Larayetan, R.; Olisah, C.; Idris, O.A. Research Progression on Studies Related to Green Synthesis Nanoparticles: A Bibliometric Review. In Green Synthesis of Nanoparticles: Applications and Prospects; Springer: Singapore, 2020; pp. 1–22. [Google Scholar]
- Olisah, C.; Okoh, O.O.; Okoh, A.I. Global evolution of organochlorine pesticides research in biological and environmental matrices from 1992 to 2018: A bibliometric approach. Emerg. Contam. 2019, 5, 157–167. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Orimoloye, I.R.; Ololade, O.O. Global trends assessment of environmental health degradation studies from 1990 to 2018. Environ. Dev. Sustain. 2020, 23, 1–14. [Google Scholar] [CrossRef]
- Perianes-Rodriguez, A.; Waltman, L.; van Eck, N.J. Constructing bibliometric networks: A comparison between full and fractional counting. J. Informetr. 2016, 10, 1178–1195. [Google Scholar] [CrossRef] [Green Version]
- Snyder, K.A.; Tartowski, S.L. Multi-scale temporal variation in water availability: Implications for vegetation dynamics in arid and semi-arid ecosystems. J. Arid Environ. 2006, 65, 219–234. [Google Scholar] [CrossRef]
- Orimoloye, I.R.; Ololade, O.O.; Mazinyo, S.P.; Kalumba, A.M.; Ekundayo, O.Y.; Busayo, E.T.; Akinsanola, A.A.; Nel, W. Spatial assessment of drought severity in Cape Town area, South Africa. Heliyon 2019, 5, e02148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, P.; Bali, S.; Sharma, A.; Vig, A.P.; Bhardwaj, R. Effect of earthworms on growth, photosynthetic efficiency and metal uptake in Brassica juncea L. plants grown in cadmium-polluted soils. Environ. Sci. Pollut. Res. 2017, 24, 13452–13465. [Google Scholar] [CrossRef]
- Fouché, T.; Maboeta, M.; Claassens, S. Effect of Biofumigants on Soil Microbial Communities and Ecotoxicology of Earthworms (Eisenia andrei). Water Air Soil Pollut. 2016, 227, 1–11. [Google Scholar] [CrossRef]
- Berry, E.C.; Jordan, D. Temperature and soil moisture content effects on the growth of Lumbricus terrestris (Oligochaeta: Lumbricidae) under laboratory conditions. Soil Biol. Biochem. 2001, 33, 133–136. [Google Scholar] [CrossRef]
- Siebert, J.; Eisenhauer, N.; Poll, C.; Marhan, S.; Bonkowski, M.; Hines, J.; Koller, R.; Ruess, L.; Thakur, M.P. Earthworms modulate the effects of climate warming on the taxon richness of soil meso- and macrofauna in an agricultural system. Agric. Ecosyst. Environ. 2019, 278, 72–80. [Google Scholar] [CrossRef]
- Wever, L.A.; Lysyk, T.J.; Clapperton, M.J. The influence of soil moisture and temperature on the survival, aestivation, growth and development of juvenile Aporrectodea tuberculata (Eisen) (Lumbricidae). Pedobiologia 2001, 45, 121–133. [Google Scholar] [CrossRef]
- Albarrán, P.; Crespo, J.A.; Ortuño, I.; Ruiz-Castillo, J. The skewness of science in 219 sub-fields and a number of aggregates. Scientometrics 2011, 88, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Midgley, G.F.; Hannah, L.; Millar, D.; Thuiller, W.; Booth, A. Developing regional and species-level assessments of climate change impacts on biodiversity in the Cape Floristic Region. Biol. Conserv. 2003, 112, 87–97. [Google Scholar] [CrossRef]
- Travers, M.; Shin, Y.-J.; Jennings, S.; Machu, E.; Huggett, J.A.; Field, J.G.; Cury, P.M. Two-way coupling versus one-way forcing of plankton and fish models to predict ecosystem changes in the Benguela. Ecol. Model. 2009, 220, 3089–3099. [Google Scholar] [CrossRef]
- Hecky, R.E.; Mugidde, R.; Ramlal, P.S.; Talbot, M.R.; Kling, G.W. Multiple stressors cause rapid ecosystem change in Lake Victoria. Freshw. Biol. 2010, 55, 19–42. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.C.; Loewen, C.J.G.; Vinebrooke, R.D.; Chimimba, C.T. Net effects of multiple stressors in freshwater ecosystems: A meta-analysis. Glob. Chang. Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef]
- Erasmus, B.F.N.; Van Jaarsveld, A.S.; Chown, S.L.; Kshatriya, M.; Wessels, K.J. Vulnerability of South African animal taxa to climate change. Glob. Chang. Biol. 2002, 8, 679–693. [Google Scholar] [CrossRef]
- Chown, S.L.; McGeoch, M.A.; Marshall, D.J. Diversity and conservation of invertebrates on the sub-Antarctic Prince Edward Islands. Afr. Entomol. 2002, 10, 67–82. [Google Scholar]
- Cassia, R.; Nocioni, M.; Correa-Aragunde, N.; Lamattina, L. Climate change and the impact of greenhouse gasses: CO2 and NO, friends and foes of plant oxidative stress. Front. Plant Sci. 2018, 9, 273. [Google Scholar] [CrossRef]
- Amador, J.A.; Görres, J.H. Role of the anecic earthworm Lumbricus terrestris L. in the distribution of plant residue nitrogen in a corn (Zea mays)-soil system. Appl. Soil Ecol. 2005, 30, 203–214. [Google Scholar] [CrossRef]
- Carney, K.M.; Hungate, B.A.; Drake, B.G.; Megonigal, J.P. Altered soil microbial community at elevated CO2 leads to loss of soil carbon. Proc. Natl. Acad. Sci. USA 2007, 104, 4990–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis: Summary for Policymakers Technical Summary Frequently Asked Questions, the Intergovernmental Panel on Climate Change. 2013. Available online: https://policycommons.net/artifacts/67690/climate-change-2013/ (accessed on 8 February 2022).
- IPCC. The IPCC Climate Change Report: Why It Matters to Everyone on the Planet. Available online: https://www.nrdc.org/stories/ipcc-climate-change-report-why-it-matters-everyone-planet (accessed on 19 October 2020).
- Brevik, E.C. The Potential Impact of Climate Change on Soil Properties and Processes and Corresponding Influence on Food Security. Agriculture 2013, 3, 398–417. [Google Scholar] [CrossRef] [Green Version]
- Guillot, E.; Hinsinger, P.; Dufour, L.; Roy, J.; Bertrand, I. With or without trees: Resistance and resilience of soil microbial communities to drought and heat stress in a Mediterranean agroforestry system. Soil Biol. Biochem. 2019, 129, 122–135. [Google Scholar] [CrossRef]
- Riah-Anglet, W.; Trinsoutrot-Gattin, I.; Martin-Laurent, F.; Laroche-Ajzenberg, E.; Norini, M.P.; Latour, X.; Laval, K. Soil microbial community structure and function relationships: A heat stress experiment. Appl. Soil Ecol. 2015, 86, 121–130. [Google Scholar] [CrossRef]
- Lacetera, N. Impact of climate change on animal health and welfare. Anim. Front. 2019, 9, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Dowd, W.W.; King, F.A.; Denny, M.W. Thermal variation, thermal extremes and the physiological performance of individuals. J. Exp. Biol. 2015, 218, 1956–1967. [Google Scholar] [CrossRef] [Green Version]
- Osborne, C.P.; Charles-Dominique, T.; Stevens, N.; Bond, W.J.; Midgley, G.; Lehmann, C.E.R. Human impacts in African savannas are mediated by plant functional traits. New Phytol. 2018, 220, 10–24. [Google Scholar] [CrossRef]
- Denny, M.W.; Dowd, W.W. Biophysics, environmental stochasticity, and the evolution of thermal safety margins in intertidal limpets. J. Exp. Biol. 2012, 215, 934–947. [Google Scholar] [CrossRef] [Green Version]
- Lacetera, N.; Segnalini, M.; Bernabucci, U.; Ronchi, B.; Vitali, A.; Tran, A.; Guis, H.; Caminade, C.; Calvete, C.; Morse, A.; et al. Climate Induced Effects on Livestock Population and Productivity in the Mediterranean Area. Adv. Glob. Chang. Res. 2013, 51, 135–156. [Google Scholar]
- Mcnally, A.; Verdin, K.; Harrison, L.; Getirana, A.; Jacob, J.; Shukla, S.; Arsenault, K.; Peters-Lidard, C.; Verdin, J.P. Acute Water-Scarcity Monitoring for Africa. Water 2019, 11, 1968. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Guillén, R.A.; Córdoba-Aguilar, A.; Hansson, B.; Ott, J.; Wellenreuther, M. Evolutionary consequences of climate-induced range shifts in insects. Biol. Rev. 2016, 91, 1050–1064. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, A.H.; Ghasura, R.S.; Mir, N.A.; Bumla, N.A.; Sankar, G.; Wani, S.A. Biochemical and physiological changes during thermal stress in bovines: A review. Iran. J. Appl. Anim. Sci. 2013, 3, 423–430. [Google Scholar]
- Idris, O.A.; Wintola, O.A.; Afolayan, A.J. Phytochemical and antioxidant activities of Rumex crispus L. in treatment of gastrointestinal helminths in Eastern Cape Province, South Africa. Asian Pac. J. Trop. Biomed. 2017, 7, 1071–1078. [Google Scholar] [CrossRef]
- Jimoh, M.A.; Idris, O.A.; Jimoh, M.O. Cytotoxicity, Phytochemical, Antiparasitic Screening, and Antioxidant Activities of Mucuna pruriens (Fabaceae). Plants 2020, 9, 1249. [Google Scholar] [CrossRef]
- Farnese, F.S.; Menezes-Silva, P.E.; Gusman, G.S.; Oliveira, J.A. When bad guys become good ones: The key role of reactive Oxygen species and Nitric Oxide in the plant responses to abiotic stress. Front. Plant Sci. 2016, 7, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.M.; Jess, M.; Williams, S.E. Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.L.; Fujiwara, Y.; Kondo, T. Mechanism of cell death induction by nitroxide and hyperthermia. Free Radic. Biol. Med. 2006, 40, 1131–1143. [Google Scholar] [CrossRef]
- McNutt, J.W.; Groom, R.; Woodroffe, R. Ambient temperature provides an adaptive explanation for seasonal reproduction in a tropical mammal. J. Zool. 2019, 309, 153–160. [Google Scholar] [CrossRef]
- Monaco, C.J.; McQuaid, C.D. Climate warming reduces the reproductive advantage of a globally invasive intertidal mussel. Biol. Invasions 2019, 21, 2503–2516. [Google Scholar] [CrossRef]
- Kurake, N.; Ishikawa, K.; Tanaka, H.; Hashizume, H.; Nakamura, K.; Kajiyama, H.; Toyokuni, S.; Kikkawa, F.; Mizuno, M.; Hori, M. Non-thermal plasma-activated medium modified metabolomic profiles in the glycolysis of U251SP glioblastoma. Arch. Biochem. Biophys. 2019, 662, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kapoor, N.; Mahajan, R. Metabolic responses of medicinal plants to global warming, temperature and heat stress. In Medicinal Plants and Environmental Challenges; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 69–80. ISBN 9783319687179. [Google Scholar]
- Sejian, V.; Bhatta, R.; Gaughan, J.B.; Dunshea, F.R.; Lacetera, N. Review: Adaptation of animals to heat stress. Animal 2018, 12, S431–S444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Singh, J.; Vig, A.P. Effect of abiotic factors on the distribution of earthworms in different land use patterns. J. Basic Appl. Zool. 2016, 74, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Schädler, M.; Demetrio, W.; Brown, G.G.; Eisenhauer, N. Climate change effects on earthworms—A review Europe PMC Funders Group. Soil Org. 2019, 91, 114–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Guo, Y.; Scarlat, M.M.; Lv, G.; Yang, X.G.; Hu, Y.C. Bibliometric study of the orthopaedic publications from China. Int. Orthop. 2018, 42, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Haunschild, R.; Bornmann, L.; Marx, W. Climate Change Research in View of Bibliometrics. PLoS ONE 2016, 11, e0160393. [Google Scholar] [CrossRef] [Green Version]
- Scheiter, S.; Higgins, S.I. Impacts of climate change on the vegetation of Africa: An adaptive dynamic vegetation modelling approach. Glob. Chang. Biol. 2009, 15, 2224–2246. [Google Scholar] [CrossRef]
- Goosse, H.; Kay, J.E.; Armour, K.C.; Bodas-Salcedo, A.; Chepfer, H.; Docquier, D.; Jonko, A.; Kushner, P.J.; Lecomte, O.; Massonnet, F.; et al. Quantifying climate feedbacks in polar regions. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Zyoud, S.H.; Waring, W.S.; Al-Jabi, S.W.; Sweileh, W.M.; Rahhal, B.; Awang, R. Intravenous Lipid Emulsion as an Antidote for the Treatment of Acute Poisoning: A Bibliometric Analysis of Human and Animal Studies. Basic Clin. Pharmacol. Toxicol. 2016, 119, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Dangles, O.; Loirat, J.; Freour, C.; Serre, S.; Vacher, J.; Le Roux, X. Research on Biodiversity and Climate Change at a Distance: Collaboration Networks between Europe and Latin America and the Caribbean. PLoS ONE 2016, 11, e0157441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olisah, C.; Adams, J.B. Systematic mapping of organophosphate contaminant (OPC) research trends between 1990 and 2018. Environ. Geochem. Health 2020, 42, 3481–3505. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Value |
|---|---|
| Article | 127 |
| Book chapter | 5 |
| Review | 18 |
| Total publications | 150 |
| Sources (journals, books, etc.) | 112 |
| Authors | 692 |
| Author appearances | 813 |
| Authors of single-authored documents | 5 |
| Authors of multi-authored documents | 687 |
| Single-authored documents | 6 |
| Authors per document | 4.61 |
| Average citations per document | 27.85 |
| Collaboration index | 4.77 |
| Keywords plus (ID) | 1099 |
| Author’s keywords (DE) | 622 |
| Most relevant keyword | Climate change |
| Source with the highest number of articles | Plos one |
| Most productive country | South Africa |
| Most affiliated institute | Stellenbosch University |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idris, O.A.; Opute, P.; Orimoloye, I.R.; Maboeta, M.S. Climate Change in Africa and Vegetation Response: A Bibliometric and Spatially Based Information Assessment. Sustainability 2022, 14, 4974. https://doi.org/10.3390/su14094974
Idris OA, Opute P, Orimoloye IR, Maboeta MS. Climate Change in Africa and Vegetation Response: A Bibliometric and Spatially Based Information Assessment. Sustainability. 2022; 14(9):4974. https://doi.org/10.3390/su14094974
Chicago/Turabian StyleIdris, Oladayo Amed, Prosper Opute, Israel Ropo Orimoloye, and Mark Steve Maboeta. 2022. "Climate Change in Africa and Vegetation Response: A Bibliometric and Spatially Based Information Assessment" Sustainability 14, no. 9: 4974. https://doi.org/10.3390/su14094974
APA StyleIdris, O. A., Opute, P., Orimoloye, I. R., & Maboeta, M. S. (2022). Climate Change in Africa and Vegetation Response: A Bibliometric and Spatially Based Information Assessment. Sustainability, 14(9), 4974. https://doi.org/10.3390/su14094974