
Differences in Ecological Traits between Plants Grown In Situ and Ex Situ and Implications for Conservation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Plants
2.2. Gas Exchange and Leaf Photosynthesis
2.3. Relative Chlorophyll Content (SPAD)
2.4. Leaf Water Potential
2.5. Leaf Water Content
2.6. Data Analyses
2.6.1. Light Response Curve Fitting
2.6.2. Light Energy Utilization Efficiency (LUE) and Water Use Efficiency (WUE) Curve Fitting
2.7. Statistical Test
3. Results
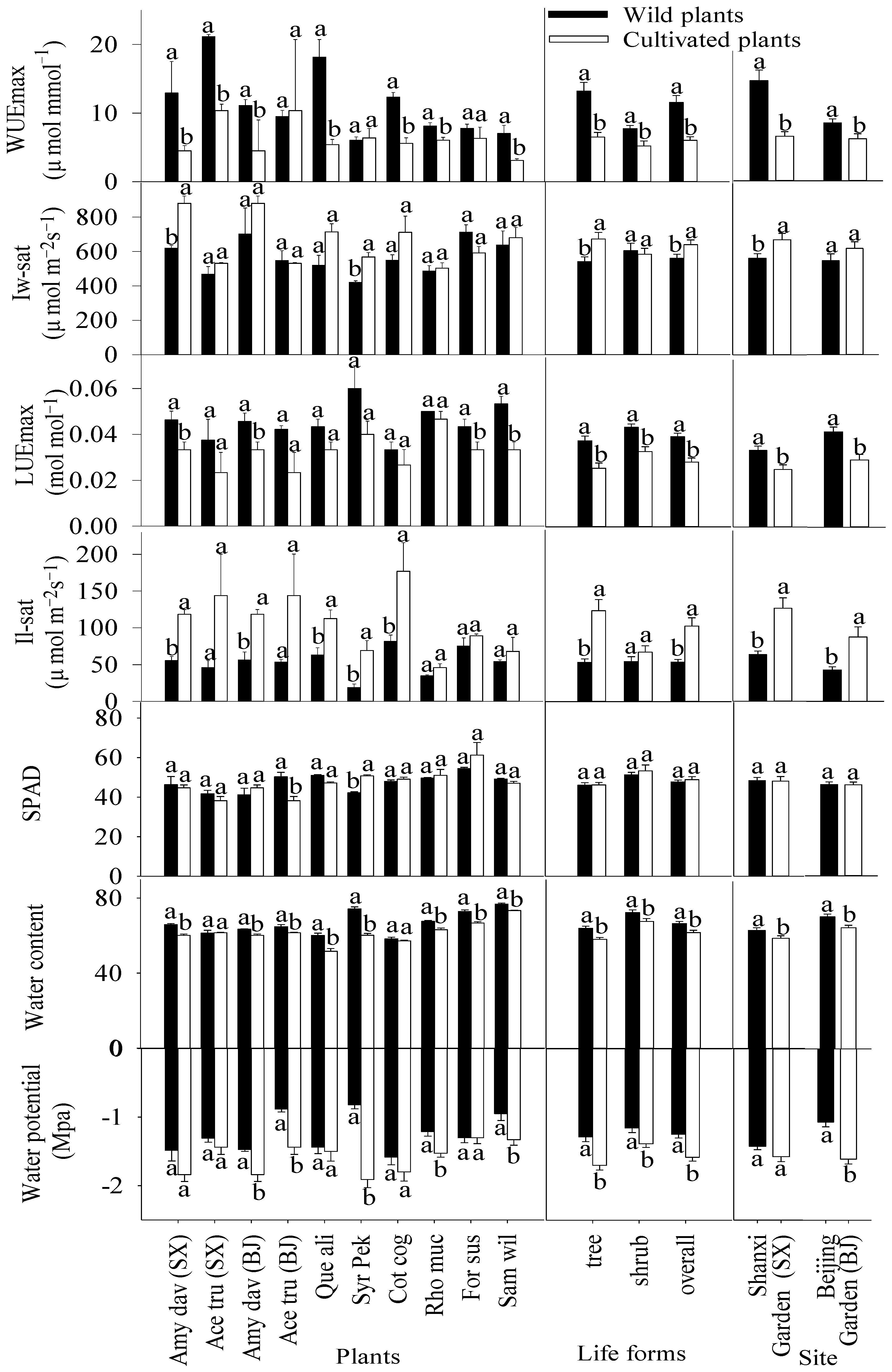
3.1. WUEmax, LUEmax, Relative Chlorophyll Content and Water Physiology of Plants in Natural Sites and Botanical Gardens
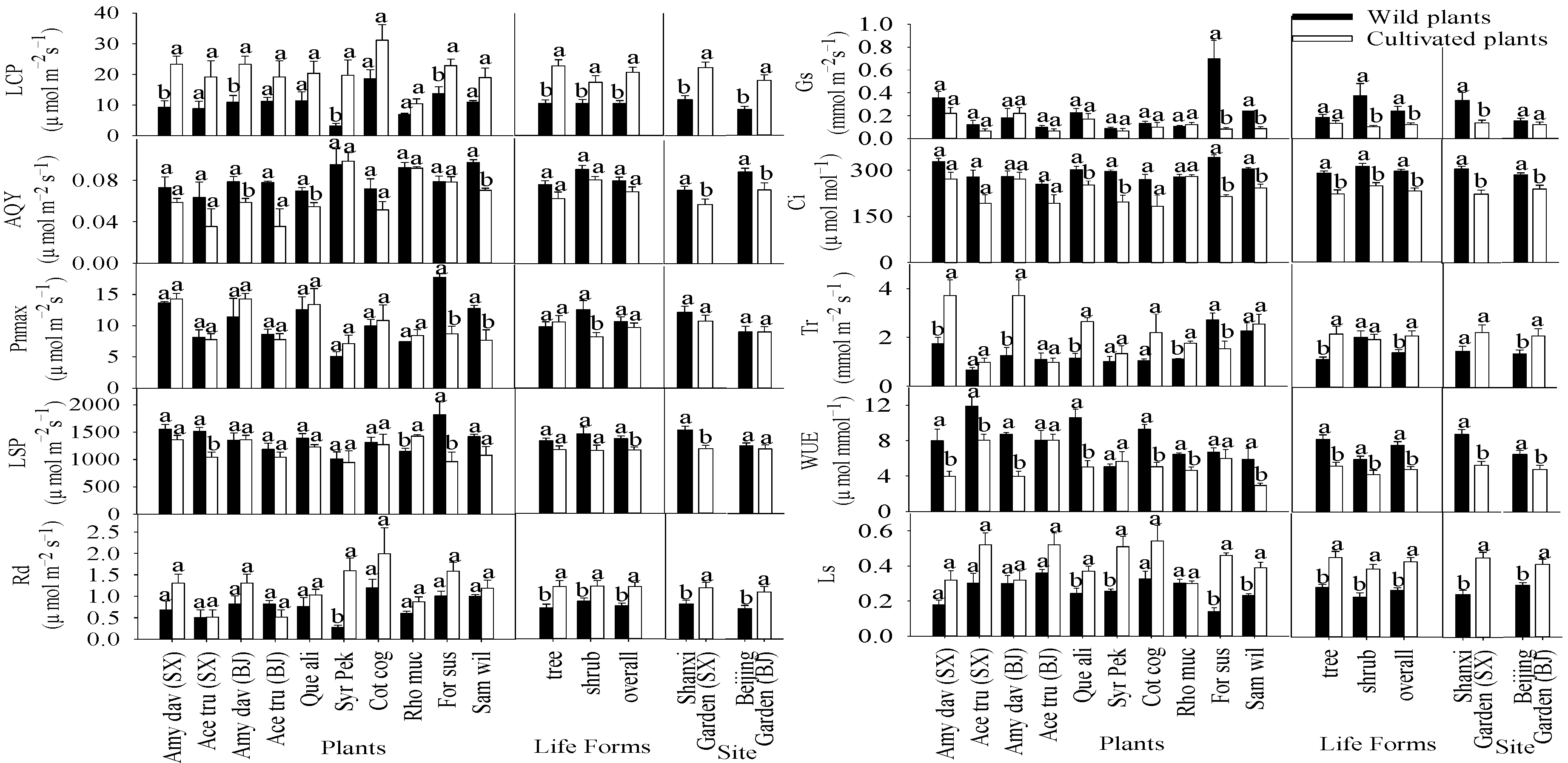
3.2. Light Response, CO2 Response, Light Energy and Water Use Efficiency Curves
3.3. Photosynthetic Parameters of Plants in Natural Sites and Botanical Gardens
4. Discussion
4.1. Ecophysiological Traits of the Same Plants in Different Habitats
4.2. Are the Ranges of Changes in the Ecophysiological Traits of Plants after Ex Situ Conservation in Different Regions Consistent?
4.3. Response of Ecophysiological Traits to Ex Situ Conservation
4.4. Implications for Fitness of Ex Situ Plants Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rivers, M.; Shaw, K.; Beech, E.; Jones, M. Conserving the World’s Most Threatened Trees: A Global Survey of Ex Situ Collections; BGCI: Richmond, UK, 2015; Available online: https://globaltrees.org/wp-content/uploads/2015/10/webLR.pdf (accessed on 10 October 2021).
- Lundholm, J.T. Green roof plant species diversity improves ecosystem multifunctionality. J. Appl. Ecol. 2015, 52, 726–734. [Google Scholar] [CrossRef]
- Li, D.Z.; Pritchard, H.W. The science and economics of ex situ plant conservation. Trends Plant Sci. 2009, 14, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Mounce, R.; Smith, P.; Brockington, S. Ex situ conservation of plant diversity in the world’s botanic gardens. Nat. Plants 2017, 3, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Charles, D. The threat to the world’s plants. Science 2008, 320, 1000. [Google Scholar] [CrossRef] [PubMed]
- Werden, L.K.; Sugii, N.C.; Weisenberger, L.; Keir, M.J.; Koob, G.; Zahawi, R.A. Ex situ conservation of threatened plant species in island biodiversity hotspots: A case study from Hawai’i. Biol. Conserv. 2020, 243, 108435. [Google Scholar] [CrossRef]
- Lambertini, M.; Leape, J.; Marton-Lefevre, J.; Mitter-Meier, R.A.; Rose, M.; Robinson, J.; Stuart, S.; Waldman, B.; Genovesi, P. Invasives: A Major Conservation Threat. Science 2011, 333, 404–405. [Google Scholar] [CrossRef]
- Cohen, J.I.; Williams, J.T.; Plucknett, D.L.; Shands, H. Ex situ conservation of plant genetic resources: Global development and environmental concerns. Science 1991, 253, 866–872. [Google Scholar] [CrossRef]
- McCarthy, J.K.; Wiser, S.K.; Bellingham, P.J.; Beresford, R.M.; Campbell, R.E.; Turner, R.; Richardson, S.J. Using spatial models to identify refugia and guide restoration in response to an invasive plant pathogen. J. Appl. Ecol. 2021, 58, 192–201. [Google Scholar] [CrossRef]
- Hultine, K.R.; Majure, L.C.; Nixon, V.S.; Arias, S.; Búrquez, A.; Goettsch, B.; Puente-Martinez, R.; Zavala-Hurtado, J. The role of botanical gardens in the conservation of cactaceae. Biosience 2016, 66, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Primack, R.B.; Miller-Rushing, A.J. The role of botanical gardens in climate change research. New Phytol. 2009, 182, 303–313. [Google Scholar] [CrossRef]
- Egerer, M.H.; Lin, B.; Threlfall, C.G.; Kendal, D. Temperature variability influences urban garden plant richness and gardener water use behavior, but not planting decisions. Sci. Total Environ. 2018, 646, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U.; Penuelas, J. Gardening and urban landscaping: Significant players in global change. Trends Plant Sci. 2008, 13, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ensslin, A.; Tschope, O.; Burkart, M.; Joshi, J. Fitness decline and adaptation to novel environments in ex situ plant collections: Current knowledge and future perspectives. Biol. Conserv. 2015, 192, 394–401. [Google Scholar] [CrossRef]
- Lou, Y.J.; Kapfer, J.; Smith, P.; Liu, Y.; Jiang, M.; Lu, X.G.; Ashcroft, M. Abundance changes of marsh plant species over 40 years are better explained by niche position water level than functional traits. Ecol. Indic. 2020, 117, 106639. [Google Scholar] [CrossRef]
- Malecore, E.M.; Dawson, W.; Kempel, A.; Muller, G.; van Kleunen, M. Nonlinear effects of phylogenetic distance on early-stage establishment of experimentally introduced plants in grassland communities. J. Ecol. 2019, 107, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Han, T.T.; Ren, H.; Wang, J.; Lu, H.F.; Song, G.M.; Chazdon, R.L. Variations of leaf eco-physiological traits in relation to environmental factors during forest succession. Ecol. Indic. 2020, 117, 106511. [Google Scholar] [CrossRef]
- Onoda, Y.; Hirose, T.; Hikosaka, K. Does leaf photosynthesis adapt to CO2-enriched environments? An experiment on plants originating from three natural CO2 springs. New Phytol. 2009, 182, 698–709. [Google Scholar] [CrossRef]
- Niinemets, U.; Keenan, T.F.; Hallik, L. A worldwide analysis of within-canopy variations in leaf structural, chemical and physiological traits across plant functional types. New Phytol. 2015, 205, 973–993. [Google Scholar] [CrossRef] [Green Version]
- Tsialtas, J.T.; Handley, L.L.; Kassioumi, M.T.; Veresoglou, D.S.; Gagianas, A.A. Interspecific variation in potential water-use efficiency and its relation to plant species abundance in a water-limited grassland. Funct. Ecol. 2001, 15, 605–614. [Google Scholar] [CrossRef]
- Cai, Y.D.; Liu, D.M.; Yao, Y.B.; Li, J.G.; Qiu, Y.K. Geological controls on prediction of coalbed methane of No. 3 coal seam in Southern Qinshui Basin, North China. Int. J. Coal Geol. 2011, 88, 101–112. [Google Scholar] [CrossRef]
- Lu, X. The annual cycle of molt in the Cape hare Lepus capensis in northern China: Pattern, timing and duration. Acta Theri. 2003, 48, 373–384. [Google Scholar] [CrossRef]
- Guo, L.D.; Huang, G.R.; Wang, Y. Seasonal and tissue age influences on endophytic fungi of Pinus tabulaeformis (Pinaceae) in the Dongling Mountains, Beijing. J. Integr. Plant Biol. 2008, 50, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Bera, S.; Yao, Y.F.; David, K.F.; Li, C.S. Tree barks as a natural trap for airborne spores and pollen grains from China. Chin. Sci. Bull. 2014, 59, 2331–2339. [Google Scholar] [CrossRef]
- Lobo, F.D.; de Barro, M.P.; Dalmagro, H.J.; Dalmolin, A.C.; Pereira, W.; de Souza, C.; Vourlitis, G.; Ortiz, C. Fitting net photosynthetic light-response curves with Microsoft Excel a critical look at the models. Photosynthetic 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Tarin, T.; Nolan, R.H.; Medlyn, B.E.; Cleverly, J.; Eamus, D. Water-use efficiency in a semi-arid woodland with high rainfall variability. Glob. Change Biol. 2019, 26, 496–508. [Google Scholar] [CrossRef]
- Garnier, E.; Laurent, G. Leaf anatomy, specific mass and water content of common annual and perennial grasses. New Phytol. 1994, 128, 725–736. [Google Scholar] [CrossRef]
- Freschet, G.H.; Swart, E.M.; Cornelissen, J.H.C. Integrated plant phenotypic responses to contrasting above-and below-ground resources: Key roles of specific leaf area and root mass fraction. New Phytol. 2015, 206, 1247–1260. [Google Scholar] [CrossRef]
- Ye, Z.P.; Suggett, D.J.; Robakowski, P.; Kang, H.J. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Still, C.J.; Randerson, J.T.; Fung, I.L. Large-scale plant light-use efficiency inferred from the seasonal cycle of atmospheric CO(2). Glob. Change Biol. 2004, 10, 1240–1252. [Google Scholar] [CrossRef] [Green Version]
- Loi, F.; Cordova, L.A.; Zhang, R.; Pajarinen, J.; Lin, T.H.; Goodman, S.B.; Yao, Z.Y. The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem Cell Res. Ther. 2016, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.E.; Mickelbart, M.V.; Jacobs, D.F. Leaf physiology and biomass allocation of backcross hybrid American chestnut (Castanea dentata) seedlings in response to light and water availability. Tree Physiol. 2014, 34, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Sim, D.A.; Rahman, A.F.; Cordova, V.D.; Baldocchi, D.D.; Flanagan, B.; Goldstein, A.; Hollinger, D.; Mission, M.; Monson, R.; Schmid, H.; et al. Midday values of gross CO2 flux and light use efficiency during satellite overpasses can be used to directly estimate eight-day mean flux. Agric. For. Meteorol. 2005, 131, 1–12. [Google Scholar] [CrossRef]
- Xu, H.; Xiao, J.F.; Zhang, Z.Q.; Ollinger, S.V.; Hollinger, D.Y.; Pan, Y.D.; Wan, J.M. Canopy photosynthetic capacity drives contrasting age dynamics of resource use efficiencies between mature temperate evergreen and deciduous forests. Glob. Change Biol. 2020, 26, 6156–6167. [Google Scholar] [CrossRef] [PubMed]
- Peters, W.; van der Velde, I.R.; van Schaik, E.; Miller, J.B.; Ciais, P.; Duarte, H.; van der Laan-Luijkx, I.; van der Molen, M.; Scholze, M.; Schaeferet, K.; et al. Increased water-use efficiency and reduced CO2 uptake by plants during droughts at a continental scale. Nat. Geosci. 2018, 11, 774. [Google Scholar] [CrossRef]
- Takanashi, S.; Kosugi, Y.; Matsuo, N.; Tani, M.; Ohte, N. Patchy stomatal behavior in broad-leaved trees grown in different habitats. Tree Physiol. 2006, 26, 1565–1578. [Google Scholar] [CrossRef] [Green Version]
- Badr, A.; Bruggemann, W. Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica 2020, 58, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.L.; Nie, Y.P.; Chen, H.S.; Wang, K.L.; Querejeta, J.I. Water uptake depth is coordinated with leaf water potential, water-use efficiency and drought vulnerability in karst vegetation. New Phytol. 2020, 229, 1339–1353. [Google Scholar] [CrossRef]
- Li, Y.; Song, X.; Li, S.; Salter, W.T.; Barbour, M.M. The role of leaf water potential in the temperature response of mesophyll conductance. New Phytol. 2020, 225, 1193–1205. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–371. [Google Scholar] [CrossRef]
- Rucinska, A.; Puchalski, J. Comparative molecular studies on the genetic diversity of an ex situ garden collection and its source population of the critically endangered polish endemic plant Cochlearia polonica E. Frohlich. Biodivers. Conserv. 2011, 20, 401–413. [Google Scholar] [CrossRef]
- Griffith, M.P.; Calonje, M.; Meerow, A.W.; Tut, F.; Kramer, A.T.; Hird, A.; Magellan, T.M.; Husby, C.E. Can botanic garden cycad collection capture the genetic diversity in a wild population? Int. J. Plant Sci. 2015, 176, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M.; Dayrell, R.L.C.; Wardil, M.V.; Damasceno, A.; Fernandes, T.; Castilho, A.; Santos, F.M.G.; Silveira, F.A.O. Seed functional traits provide support for ecological restoration and ex situ conservation in the threatened amazon ironstone outcrop flora. Front. Plant Sci. 2020, 11, 599496. [Google Scholar] [CrossRef] [PubMed]
- Gerstenberg, T.; Hofmann, M. Perception and preference of trees: A psychological contribution to tree species selection in urban areas. Urban For. Urban Green 2016, 15, 103–111. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Effect | |
|---|---|---|
| Sites | Plants | |
| AQY | 3.74 ns | 0.27 ns |
| Rd | 7.56 ** | 0.15 ns |
| Pnmax | 4.80 * | 5.40 * |
| LSP | 3.91 * | 0.75 ns |
| LCP | 22.77 *** | 0.00 ns |
| Il-sat | 51.36 *** | 0.45 ns |
| LUEmax | 2.33 ns | 0.33 ns |
| Iw-sat | 11.57 ** | 2.93 ns |
| WUEmax | 19.93 *** | 1.85 ns |
| φ | 27.21 *** | 11.59 ** |
| LWC | 11.77 ** | 1.87 ns |
| SPAD | 0.23 ns | 0.48 ns |
| Pn | 4.79 * | 5.34 * |
| Gs | 2.07 ns | 6.40 * |
| Ci | 2.40 ns | 2.84 ns |
| Tr | 23.09 *** | 1.40 ns |
| Ls | 2.38 ns | 2.85 ns |
| WUE | 25.01 *** | 2.28 ns |
| df | 2 | 1 |
| Parameters | Effect | |||
|---|---|---|---|---|
| Sites | Plants | |||
| BS | BB | BS | BB | |
| AQY | 0.756 ns | 2.19 ns | 2.94 ns | 1.32 ns |
| Rd | 7.61 * | 22.12 *** | 4.07 * | 2.66 ns |
| Pnmax | 2.70 ns | 0.65 ns | 1.47 ns | 8.16 ** |
| LSP | 8.08 * | 0.18 ns | 0.24 ns | 3.70 ns |
| LCP | 13.75 ** | 20.96 *** | 3.82 ns | 3.14 ns |
| Il-sat | 12.76 ** | 9.71 ** | 4.04 * | 2.53 ns |
| LUEmax | 7.11 * | 11.27 ** | 2.78 ns | 0.87 ns |
| Iw-sat | 2.94 ns | 3.28 ns | 0.20 ns | 8.36 ** |
| WUEmax | 39.62 *** | 8.34 * | 6.07 * | 3.20 ns |
| φ | 1.00 ns | 76.12 *** | 6.29 * | 5.07 * |
| LWC | 44.69 *** | 139.20 *** | 124.26 *** | 91.05 *** |
| SPAD | 0.23 ns | 3.11 ns | 5.48* | 2.34 ns |
| Pn | 2.81 ns | 0.66 ns | 1.89 ns | 8.11 ** |
| Gs | 15.34 ** | 17.67 ** | 7.33 ** | 16.30 *** |
| Ci | 36.12 *** | 33.20 *** | 5.50 * | 4.70 * |
| Tr | 2.69 ns | 3.67 ns | 0.99 ns | 11.81 * |
| Ls | 35.71 *** | 31.27 *** | 5.51 * | 4.24 * |
| WUE | 35.92 *** | 5.75 * | 2.13 ns | 1.50 ns |
| df | 1 | 1 | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Lai, L.; Zhou, J.; Yi, S.; Liu, X.; Guo, J.; Zheng, Y. Differences in Ecological Traits between Plants Grown In Situ and Ex Situ and Implications for Conservation. Sustainability 2022, 14, 5199. https://doi.org/10.3390/su14095199
Sun Q, Lai L, Zhou J, Yi S, Liu X, Guo J, Zheng Y. Differences in Ecological Traits between Plants Grown In Situ and Ex Situ and Implications for Conservation. Sustainability. 2022; 14(9):5199. https://doi.org/10.3390/su14095199
Chicago/Turabian StyleSun, Qinglin, Liming Lai, Jihua Zhou, Sangui Yi, Xin Liu, Jiaojiao Guo, and Yuanrun Zheng. 2022. "Differences in Ecological Traits between Plants Grown In Situ and Ex Situ and Implications for Conservation" Sustainability 14, no. 9: 5199. https://doi.org/10.3390/su14095199
APA StyleSun, Q., Lai, L., Zhou, J., Yi, S., Liu, X., Guo, J., & Zheng, Y. (2022). Differences in Ecological Traits between Plants Grown In Situ and Ex Situ and Implications for Conservation. Sustainability, 14(9), 5199. https://doi.org/10.3390/su14095199