
Nanoparticle Mediated Plant Tolerance to Heavy Metal Stress: What We Know?

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
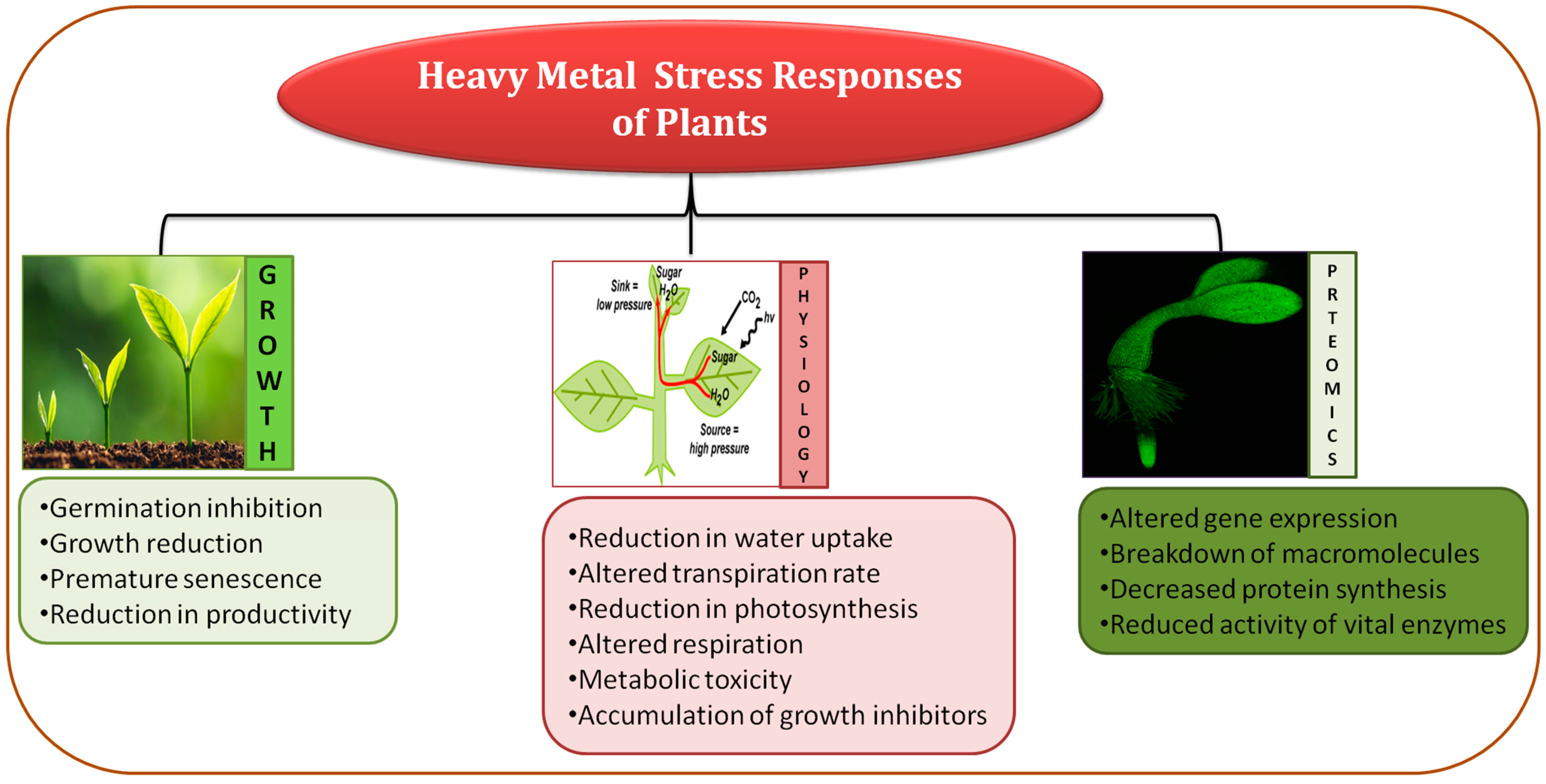
2. Mechanism of Plant Growth Inhibition Induced by Heavy Metals
2.1. Cadmium (Cd)
2.2. Lead (Pb)
2.3. Chromium (Cr)
2.4. Iron (Fe)
2.5. Manganese (Mn)
3. Role of Nanoparticles on Heavy Metal Stress Tolerance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kou, T.J.; Yu, W.W.; Lam, S.K.; Chen, D.L.; Hou, Y.P.; Li, Z.Y. Differential root responses in two cultivars of winter wheat (Triticum aestivum L.) to elevated ozone concentration under fully open-air field conditions. J. Agron. Crop Sci. 2018, 204, 325–332. [Google Scholar]
- Fortina, P.; Kricka, L.J.; Surrey, S.; Grodzinski, P. Nanotechnology, the promise and reality of new approaches to molecular recognition. Trends Biotechnol. 2005, 23, 168. [Google Scholar] [CrossRef] [PubMed]
- Raje, K.; Ojha, S.; Mishra, A.; Munde, V.K.; Rawat, C.; Chaudhary, S.K. Impact of supplementation of mineral nano particles on growth performance and health status of animals, A review. J. Ento. Zool. Stud. 2018, 6, 1690–1694. [Google Scholar]
- Menazea, A.A.; Abdelghany, A.M.; Hakeem, N.A.; Osman, W.H.; Abd El-kader, F.H. Nd:YAG Nanosecond Laser Pulses for Precipitation Silver Nanoparticles in Silicate Glasses: AC Conductivity and Dielectric Studies. Silicon 2018, 12, 13–20. [Google Scholar] [CrossRef]
- Afzal, S.; Singh, N.K. Effect of zinc and iron oxide nanoparticles on plant physiology, seed quality and microbial community structure in a rice-soil-microbial ecosystem. Environ. Pollut. 2022, 314, 120224. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lopez, J.I.; Nino-Medina, G.; Olivares-Saenz, E.; Lira-Saldivar, R.; Barriga-Castro, E.D.; Vazquez-Alvardo, R.; Rodriguez-Salinas, P.A.; Zavala-Garcia, F. Foliar Application of Zinc Oxide Nanoparticles and Zinc Sulfate Boosts the Content of Bioactive Compounds in Habanero Peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, E.S.; Colman, B.P.; Hochella, M.F.; Cardinale, B.J.; Nisbet, M.; Richardson, C.J.; Yin, L. An ecological perspective on nanomaterial impacts in the environment. J. Environ. Qual. 2010, 39, 1954–1965. [Google Scholar] [CrossRef]
- Migowski, P.; Dupont, J. Catalytic applications of metal nanoparticles in imidazoliumionic liquids. Chemistry 2007, 13, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.; Aftab, T.; Singh, N.K. Impact of Zinc Oxide and Iron Oxide Nanoparticles on Uptake, Translocation, and Physiological Effects in Oryza sativa L. J. Plant Growth Regul. 2022, 41, 1445–1461. [Google Scholar] [CrossRef]
- Cheng, Y.; Morshed, R.A.; Auffinger, B.; Tobias, A.L.; Lesniak, M.S. Multifunctional nanoparticles for brain tumor imaging and therapy. Adv. Drug Deliv. Rev. 2014, 66, 42–57. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.; Gusain, D.; Sharma, Y.C. A critical review on the toxicity of some widely engineered nanoparticles. Ind. Eng. Chem. Res. 2015, 54, 6209–6233. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorous-mobilizing enzyme secretion and gum contents in Clusterbean (Cyamopsis tetragonoloba L.). Agric. Res. 2013, 2, 48–57. [Google Scholar] [CrossRef]
- Zafar, H.; Ali, A.; Ali, J.S.; Haq, I.U.; Zia, M. Effect of ZnO nanoparticles on Brassica nigra seedlings and stem explants growth dynamics and antioxidative response. Front. Plant Sci. 2016, 7, 535. [Google Scholar] [CrossRef] [Green Version]
- Prasad, T.N.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination.; growth and yield of peanut. J. Plant Nut. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Furlani, P.R.; Clark, R.B. Screening sorghum for aluminium tolerance in nutrient solution. Agron. J. 1981, 73, 587–594. [Google Scholar] [CrossRef]
- Lopez-Moreno, M.; de la Rosa, G.; Hernandez-Viezcas, J.; Castillo-Michel, H.; Botez, C.; Peralta-Videa, J.; Gardea-Torresdey, J. Evidence of the differential biotransformation and genotoxicity of ZnO and CeO nanoparticles on soybean (Glycine max) plants. Environ. Sci. Technol. 2010, 44, 7315–7320. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.; Liu, L.; Ibrahim, W.; Cai, Y. Alleviating effects of exogenous glutathione, glycinebetaine, brassinosteroids and salicylic acid on cadmium toxicity in rice seedlings (Oryza sativa). Agrotech 2013, 2, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Aziz, M.E.; Morsi, S.M.M.; Salama, D.M.; Abdel-Aziz, M.S.; Abd Elwahed, M.S.; Shaaban, E.A.; Youssef, A.M. Preparation and characterization of chitosan/polyacrylicacid/copper nanocomposites and their impact on onion production. Int. J. Biol. Macromol. 2019, 123, 856–865. [Google Scholar] [CrossRef]
- Vannini, C.; Domingo, G.; Onelli, E.; Prinsi, B.; Marsoni, M.; Espen, L.; Bracale, M. Morphological and proteomic responses of Eruca sativa exposed to silver nanoparticles or silver nitrate. PLoS ONE 2013, 8, 68752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokhrel, L.R.; Dubey, B. Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 2013, 45, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Diao, Y.; Abayneh, B. Removal of Hg(0) from simulated flue gas over silver-loaded rice husk gasification char. R. Soc. Open Sci. 2018, 5, 180248. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 2012, 6, 87–90. [Google Scholar]
- Riahi-Madvar, A.; Rezaee, F.; Jalali, V. Effects of alumina nanoparticles on morphological properties and antioxidant system of Triticum aestivum. Iran. J. Plant Physiol. 2012, 3, 595–603. [Google Scholar]
- Yasmeen, F.; Raja, N.I.; Razzaq, A.; Komatsu, S. Gel-free/label-free proteomic analysis of wheat shoot in stress tolerant varieties under iron nanoparticles exposure. Biochim. Biophys. Acta 2016, 1864, 1586–1598. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles, Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef]
- Mustafa, G.; Komatsu, S. Insights into the response of soybean mitochondrial proteins to various sizes of aluminum oxide nanoparticles under flooding stress. J. Proteome Res. 2016, 15, 4464–4475. [Google Scholar] [CrossRef]
- Larue, C.; Laurette, J.; Herlin-Boime, N.; Khodja, H.; Fayard, B.; Flank, A.; Brisset, F.; Brisset, M. Accumulation translocation and impact of TiO2 nanoparticles in wheat (Triticum aestivum spp.), Influence of diameter and crystal phase. Sci. Total Environ. 2012, 43, 197–208. [Google Scholar] [CrossRef]
- Hao, Y.; Fang, P.; Ma, C.; White, J.C.; Xiang, Z.; Wang, H.; Zhang, Z.; Rui, Y.; Xing, B. Engineered nanomaterials inhibit Podosphaera pannosa infection on rose leaves by regulating phytohormones. Environ. Res. 2018, 170, 1–6. [Google Scholar] [CrossRef]
- Hong, F.H.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Gao, Y.; Wu, H.; Hou, W.; Zhang, C.; Ma, H. Physiological effect of anatase TiO2 nanoparticles on Lemn. Minor. Environ. Toxicol. Chem. 2012, 31, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Maali-Amiri, R.; Abbasi, A. Effect of TiO2 nanoparticles on chickpea response to cold stress. Biol. Trace Elem. Res. 2013, 152, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Hong, F.; Lu, S.; Liu, C. Effect of nano-TiO2 on strength of naturally aged seeds and growth of spinach. Biol. Trace Elem. Res. 2005, 104, 83–91. [Google Scholar] [CrossRef]
- Linglan, M.; Chao, L.; Chunxiang, Q.; Sitao, Y.; Jie, L.; Fengqing, G.; Fashui, H. Rubisco activase mRNA expression in spinach, Modulation by nanoanatase treatment. Biol. Trace Elem. Res. 2008, 122, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T.; et al. Iron Oxide Nanoparticles as a Potential Iron Fertilizer for Peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef] [Green Version]
- Feizi, H.; Moghaddam, P.R.; Shahtahmassebi, N.; Fotovot, A. Assessment of concentration of nano and bulk iron oxide particles on early growth of wheat (Triticum aestivum L.). Annu. Rev. Res. Biol. 2013, 3, 752–761. [Google Scholar]
- Ojuederie, O.B.; Olanrewaju, O.S.; Babalola, O.O. Plant Growth Promoting Rhizobacterial Mitigation of Drought Stress in Crop Plants, Implications for Sustainable Agriculture. Agronomy 2019, 9, 712. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.J.; Wang, X.P.; Gong, P.; Tian, L.D.; Yao, T.D. Heavy metals of the Tibetan top soils Level source spatial distribution temporal variation and risk assessment. Environ. Sci. Pollut. Res. 2012, 19, 3362–3370. [Google Scholar] [CrossRef]
- Shahid, M.; Khalid, S.; Abbas, G.; Shahid, N.; Nadeem, M.; Sabir, M. Heavy metal stress and crop productivity. In Crop Production and Global Environmental Issues; Hakeem, K.R., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–25. [Google Scholar]
- Hossain, M.B.; Jahiruddin, M.; Panaullah, G.M.; Loeppert, R.H.; Islam, M.R.; Duxbury, J.M. Spatial variability of arsenic concentration in soils and plants and its relationship with iron manganese and phosphorus. Environ. Pollut. 2008, 156, 739–744. [Google Scholar] [CrossRef]
- Hassan, T.U.; Bano, A.; Naz, I. Alleviation of heavy metals toxicity by the application of plant growth promoting rhizobacteria and effects on wheat grown in saline sodic field. Int. J. Phytoremediation 2017, 19, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, A.L.; Muneer, S.; Kim, Y.H.; Al-Rawahi, A.; Al-Harrasi, A. Silicon and Salinity, Crosstalk in crop-mediated stress tolerance mechanisms. Front. Plant Sci. 2019, 10, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahv, F.; Wu, W. Soil and crop management strategies to ensure higher crop productivity within sustainable environments. Sustainability 2019, 11, 1485. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chen, L.; Chen, S.; Ma, Y. Alleviation of cadmium-induced root growth Farinhibition in crop seedlings by nanoparticles. Ecotoxicol. Environ. Saf. 2012, 79, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Sehar, S.; Rajput, V.D.; Faraz, A.; Afzal, S.; Minkina, T.; Sushkova, S.; Adil, M.F.; Yu, F.; Alatar, A.A.; et al. Modulation of cellular redox status and antioxidant defense system after synergistic application of zinc oxide nanoparticles and salicylic acid in rice (Oryza sativa) plant under arsenic stress. Plants 2021, 10, 2254. [Google Scholar] [CrossRef]
- Shen, Y.; Tang, J.; Nie, Z.; Wang, Y.; Ren, Y.; Zuo, L. Preparation and application of magnetic Fe3O4 nanoparticles for wastewater purification. Sep. Purif. Technol. 2009, 68, 312–319. [Google Scholar] [CrossRef]
- Skiba, E.; Wolf, W.M. Cerium Oxide Nanoparticles Affect Heavy Metals Uptake by Pea in a Divergent Way than Their Ionic and Bulk Counterparts. Water Air Soil Pollut. 2019, 230, 248. [Google Scholar] [CrossRef] [Green Version]
- Zadeh, R.R.; Arvin, S.M.J.; Jamei, R.; Mozaffari, H.; Nejhad, F.R. Response of tomato plants to interaction effects of magnetic (Fe3O4) nanoparticles and cadmium stress. J. Plant Interact. 2019, 14, 474–481. [Google Scholar]
- Faraji, J.; Sepehri, A.; Salcedo-Reyes, J.C. Titanium dioxide nanoparticles and sodium nitroprusside alleviate the adverse effects of cadmium stress on germination and seedling growth of wheat (Triticum aestivum L.). Univ. Sci. 2018, 23, 61–87. [Google Scholar] [CrossRef]
- Ragab, G.A.; Saad-Allah, K.M. Green synthesis of sulfur nanoparticles using Ocimum basilicum leaves and its prospective effect on manganese-stressed Helianthus annuus (L.) seedlings. Ecotoxicol. Environ. Saf. 2020, 191, 110242. [Google Scholar] [CrossRef]
- Cai, F.; Wu, X.; Zhang, H.; Shen, X.; Zhang, M.; Chen, W.; Gao, Q.; White, J.C.; Tao, S.; Wang, X. Impact of TiO2 nanoparticles on lead uptake and bioaccumulation in rice (Oryza sativa L.). Nano Impact 2017, 5, 101–108. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Nano-silicon alters antioxidant activities of soybean seedlings under salt toxicity. Protoplasma 2018, 255, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Kabir, E.; Kumar, V.; Kim, K.H.; Yip, A.C.K.; Sohn, J.R. Environmental impacts of nanomaterials. J. Environ. Manag. 2018, 225, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.F.; Sehar, S.; Chen, G.; Chen, Z.H.; Jilani, G.; Chaudhry, A.N.; Shamsi, I.H. Cadmium-zinc cross-talk delineates toxicity tolerance in rice via differential genes expression and physiological/ultrastructural adjustments. Ecotoxicol. Environ. Saf. 2020, 190, 110076. [Google Scholar] [CrossRef]
- Ahmad, I.; Akhtar, M.J.; Zahir, Z.A.; Naveed, M.; Mitter, B.; Sessitsch, A. Cadmium-tolerant bacteria induce metal stress tolerance in cereals. Environ. Sci. Pollut. Res. Int. 2014, 21, 11054–11065. [Google Scholar] [CrossRef]
- Adil, M.F.; Sehar, S.; Han, Z.; Lwalaba, J.L.W.; Jilani, G.; Zeng, F.; Chen, Z.H.; Shamsi, I.H. Zinc alleviates cadmium toxicity by modulating photosynthesis ROS homeostasis and ion flux kinetics in rice. Environ. Pollut. 2020, 265, 114979. [Google Scholar] [CrossRef]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediation 2016, 19, 133–141. [Google Scholar] [CrossRef]
- Baycu, G.; Gevrek-Kürüm, N.; Moustaka, J.; Csatári, I.; Rognes, S.E.; Moustakas, M. Cadmium-zinc accumulation and photosystem II responses of Noccaea caerulescens to Cd and Zn exposure. Environ. Sci. Pollut. Res. 2017, 24, 2840–2850. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Rehman, M.Z.; Ali, S.; Rizwan, M.; Naeem, A.; Maqsood, M.A.; Khalid, H.; Rinklebe, J.; Ok, Y.S. Residual effects of monoammonium phosphate.; gypsum and elemental sulfur on cadmium phytoavailability and translocation from soil to wheat in an effluent irrigated field. Chemosphere 2017, 174, 515–523. [Google Scholar] [CrossRef]
- Saifullah, S.N.; Bibi, S.; Ahmad, M.; Ok, Y.S. Effectiveness of zinc application to minimize cadmium toxicity and accumulation in wheat (Triticum aestivum L.). Environ. Earth Sci. 2014, 71, 1663–1672. [Google Scholar] [CrossRef]
- Vasiljeva, S.; Smirnova, G.; Basova, N.; Babarykin, D. Cadmium-Induced Oxidative Damage and Protective Action of Fractioned Red Beet (Beta vulgaris) Root Juice in Chickens. Agron. Res. 2018, 16, 1517–1526. [Google Scholar]
- Li, X.M.; Bu, N.; Li, Y.; Ma, L.; Xin, S.; Zhang, L. Growth photosynthesis and antioxidant responses of endophyte infected and non-infected rice under lead stress conditions. J. Hazard Mater. 2012, 213–214, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Chen, W.; Chang, A.C.; Page, A.L. Environmental risks of trace elements associated with long-term phosphate fertilizers applications, a review. Environ. Pollut. 2012, 168, 44–53. [Google Scholar] [CrossRef]
- Maestri, E.; Marmiroli, M.; Visioli, G.; Marmiroli, N. Metal tolerance and hyperaccumulation, costs and trade-offs between traits and environment. Environ. Exp. Bot. 2010, 68, 1–13. [Google Scholar] [CrossRef]
- Sengar, R.S.; Gautam, M.; Sengar, R.S.; Sengar, R.S.; Garg, S.K.; Sengar, K.; Chaudhary, R. Lead stress effects on physiobiochemical activities of higher plants. Rev. Environ. Contam. Toxicol. 2009, 196, 73–93. [Google Scholar]
- Jiang, W.; Liu, D. Pb-induced cellular defense system in the root meristematic cells of Allium sativum L. BMC Plant Biol. 2010, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Islam, E.; Liu, D.; Li, T.; Yang, X.; Jin, X.; Mahmood, Q.; Tian, S.; Li, J. Effect of Pb toxicity on leaf growth physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. J. Hazard. Mater. 2008, 154, 914–926. [Google Scholar] [CrossRef]
- Mohan, D.; Pittman, C.U. Activated carbons and low cost adsorbents for remediation of tri-and hexavalent chromium from water. J. Hazard. Mater. 2006, 137, 762–811. [Google Scholar] [CrossRef]
- Diwan, H.; Khan, I.; Ahmad, A.; Iqbal, M. Induction of phytochelatins and antioxidant defence system in Brassica juncea and Vigna radiata in response to chromium treatments. Plant Growth Regul. 2010, 61, 97–107. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Augustynowicz, J.; Grosicki, M.; Hanus-Fajerska, E.; Lekka, M.; Waloszek, A.; Koloczek, H. Chromium (VI) bioremediation by aquatic macrophyte Callitriche cophocarpa Sendtn. Chemosphere 2010, 79, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Becquer, T.; Quantin, C.; Sicot, M.; Boudot, J.P. Chromium availability in ultramafic soils from New Caledonia. Sci. Total Environ. 2003, 301, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Zeid, I.M. Response of Phaseolus vulgaris to chromium and cobalt treatments. Biol. Plant 2001, 44, 111–115. [Google Scholar] [CrossRef]
- Barton, L.L.; Johnson, G.V.; O’Nan, A.G.; Wagener, B.M. Inhibition of ferric chelate reductase in alfalfa roots by cobalt nickel chromium and copper. J. Plant Nutr. 2000, 23, 1833–1845. [Google Scholar] [CrossRef]
- Sing, A.K. Effect of trivalent and hexavalent chromium on spinach (Spinacea oleracea L). Environ. Ecol. 2001, 19, 807–810. [Google Scholar]
- Vajpayee, P.; Rai, U.N.; Ali, M.B.; Tripathi, R.D.; Yadav, V.; Sinha, S. Chromium induced physiological changes in Vallisneria spiralis L and its role in phytoremediation of tannery effluent. Bull. Environ. Contam. Toxicol. 2001, 67, 246–256. [Google Scholar] [PubMed]
- Zurayk, R.; Sukkariyah, B.; Baalbaki, R. Common hydrophytes as bioindicators of nickel chromium and cadmium pollution. Water Air Soil Pollut. 2001, 127, 373–388. [Google Scholar] [CrossRef]
- Celletti, S.; Lanz, M.; Bergamo, A.; Benedetti, V.; Basso, D.; Baratieri, M.; Cesco, S.; Mimmo, T. Evaluating the Aqueous Phase from Hydrothermal Carbonization of Cow Manure Digestate as Possible Fertilizer Solution for Plant Growth. Front. Plant Sci. 2022, 12, 1317. [Google Scholar] [CrossRef]
- Muller, C.; Silveira, S.; Daloso, D.M.; Mendes, G.C.; Merchant, A.; Kuki, K.N.; Oliva, M.A.; Loureiro, M.E.; Almeida, A.M. Ecophysiological responses to excess iron in lowland and upland rice cultivars. Chemosphere 2017, 189, 123–133. [Google Scholar]
- Li, W.; Lan, P. The Understanding of the Plant Iron Deficiency Responses in Strategy I Plants and the Role of Ethylene in This Process by Omic Approaches. Front. Plant Sci. 2017, 8, 40. [Google Scholar] [CrossRef]
- Bashir, K.; Rasheed, S.; Kobayashi, T.; Seki, M.; Nishizawa, N.K. Regulating Subcellular Metal Homeostasis, The Key to Crop Improvement. Front. Plant Sci. 2016, 7, 1192. [Google Scholar]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar]
- Araújo, I.P.S.; Costa, D.B.; De Moraes, R.J.B. Identification and Characterization of Particulate Matter Concentrations at Construction Jobsites. Sustainability 2014, 6, 7666–7688. [Google Scholar] [CrossRef] [Green Version]
- Siqueira-Silva, A.I.; Rios, C.O.; Pereira, E.G. Iron toxicity resistance strategies in tropical grasses, The role of apoplastic radicular barriers. J. Environ. Sci. (China) 2019, 78, 257–266. [Google Scholar]
- Santos, E.F.; Kondo Santini, J.M.; Paixão, A.P.; Júnior, E.F.; Lavres, J.; Campos, M.; dos Reis, A.R. Physiological highlights of manganese toxicity symptoms in soybean plants, Mn toxicity responses. Plant Physiol. Biochem. 2017, 113, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Alejandro, S.; Holler, S.; Meier, B.; Peiter, E. Manganese in Plants: From Acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Wu, S. Effect of manganese excess on the soybean plant cultivated under various growth conditions. J. Plant Nutr. 1994, 17, 991–1003. [Google Scholar] [CrossRef]
- Singh, J.; Lee, B.K. Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max), a possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag. 2016, 170, 88–96. [Google Scholar] [CrossRef]
- Liang, S.-X.; Jin, Y.; Liu, W.; Li, X.; Shen, S.-G.; Ding, L. Feasibility of Pb phytoextraction using nano-materials assisted ryegrass, results of a one-year field-scale experiment. J. Environ. Manag. 2017, 190, 170–175. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar]
- Lobzenko, I.; Burachevskaya, M.; Zamulina, I.; Barakhov, A.; Bauer, T.; Mandzhieva, S.; Sushkova, S.; Minkina, T.; Tereschenko, A.; Kalinichenko, V.; et al. Development of a Unique Technology for the Pyrolysis of Rice Husk Biochar for Promising Heavy Metal Remediation. Agriculture 2022, 12, 1689. [Google Scholar] [CrossRef]
- Konate, A.; He, X.; Zhang, Z.; Ma, Y.; Zhang, P.; Alugongo, G.M.; Rui, Y. Magnetic (Fe3O4) nanoparticles reduce heavy metals uptake and mitigate their toxicity in wheat seedling. Sustainability 2017, 9, 790. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Ur Rehman, M.Z.; Malik, S.; Adrees, M.; Qayyum, M.F.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Effect of foliar applications of silicon and titanium dioxide nanoparticles on growth oxidative stress and cadmium accumulation by rice (Oryza sativa). Acta Physiol. Plant 2019, 41, 35. [Google Scholar] [CrossRef]
- Malar, S.; Manikandan, R.; Favas, P.J.; Sahi, S.V.; Venkatachalam, P. Effect of lead on phytotoxicity growth biochemical alterations and its role on genomic template stability in Sesbania grandiflora, a potential plant for phytoremediation. Ecotoxicol. Environ. Saf. 2014, 108, 249–257. [Google Scholar] [CrossRef]
- Marzec, Z.; Schlegel-Zawadzka, M. Exposure to cadmium lead and mercury in the adult population from Eastern Poland 1990–2002. Food Addit. Contam. 2004, 21, 963–970. [Google Scholar] [CrossRef]
- Kikuchi, T.; Okazaki, M.; Kimura, S.D.; Motobayashi, T.; Baasansuren, J.; Hattori, T.; Abe, T. Suppressive effects of magnesium oxide materials on cadmium uptake and accumulation into rice grains, II, Suppression of cadmium uptake and accumulation into rice grains due to application of magnesium oxide materials. J. Hazard. Mater. 2008, 154, 294–299. [Google Scholar] [CrossRef]
- Wang, H.; Zhong, G.; Shi, G.; Pan, F. Toxicity of Cu, Pb, and Zn on Seed Germination and Young Seedlings of Wheat (Triticum aestivum L.). In Proceedings of the International Conference on Computer and Computing Technologies in Agriculture, Nanchang, China, 22–25 October 2010; Li, D.L., Liu, Y., Chen, Y.Y., Eds.; Springer: Cham, Switzerland, 2010; pp. 231–240. [Google Scholar]
- Shaikh, I.R.; Shaikh, P.R.; Shaikh, R.A.; Shaikh, A.A. Phytotoxic effects of heavy metals (Cr, Cd, Mn and Zn) on wheat (Triticum aestivum L.) seed germination and seedlings growth in black cotton soil of Nanded, India. Res. J. Chem. Sci. 2013, 3, 14–23. [Google Scholar]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Hayat, S. Effects of Copper Oxide Nanoparticles on then Photosynthesis and Antioxidant levels of Mustard Plants (Brassica juncea). J. Biol. Chem. Res. 2018, 35, 418–426. [Google Scholar]
- Yasmeen, F.; Razzaq, A.; Iqbal, M.N.; Jhanzab, H.M. Effect of silver copper and iron nanoparticles on wheat germination. Int. J. Biosci. 2015, 6, 112–117. [Google Scholar]
- Jaberzadeh, A.; Moaveni, P.; Moghadam, H.R.T.; Zahedi, H. Influence of bulk and nanoparticles titanium foliar application on some agronomic traits.; seed gluten and starch contents of wheat subjected to water deficit stress. Not. Bot. Horti. Agrobo. 2013, 41, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Long, J.; Geng, J.; Li, J.; Wei, Z. Impact of titanium dioxide nanoparticles on Cd phytotoxicity and bioaccumulation in rice (Oryza sativa L.). Int. J. Environ. Res. Public Health 2020, 17, 2979. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Zhou, Y.; Ma, C.; Feng, Y.; Hao, Y.; Rui, Y.; Wu, W.; Gui, X.; Han, Y.; Wang, Y.; et al. Jointed toxicity of TiO2 NPs and Cd to rice seedlings, NPs alleviated Cd toxicity and Cd promoted NPs uptake. Plant Physiol. Biochem. 2017, 110, 82–93. [Google Scholar] [CrossRef]
- Aghdam, M.T.B.; Mohammadi, H.; Ghorbanpour, M. Effects of nanoparticulate anatase titanium dioxide on physiological and biochemical performance of Linum usitatissimum (Linaceae) under well-watered and drought stress conditions. Braz. J. Bot. 2016, 39, 139–146. [Google Scholar] [CrossRef]
- Kiapour, H.; Moaveni, P.; Habibi, D.; Sani, B. Evaluation of the application of gibbrellic acid and titanium dioxide nanoparticles under drought stress on some traits of basil (Ocimum basilicum L.). Int. J. Agron. Agric. Res. 2015, 6, 138–150. [Google Scholar]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica Nanoparticles on Basil (Ocimum basilicum) Under Salinity Stress. J. Chem. Health Risks 2014, 4, 49–55. [Google Scholar]
- Sabaghnia, N.; Janmohammadi, M. Effect of nano-silicon particles application on salinity tolerance in early growth of some lentil genotypes/Wpływ nanocząstek krzemionki na tolerancję zasolenia we wczesnym rozwoju niektórych genotypów soczewicy. Ann. UMCS Biol. 2015, 69, 39. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef]
- Soliman, A.S.; El-feky, S.A.; Darwish, E. Alleviation of salt stress on Moringa peregrina using foliar application of nanofertilizers. J. Hort. Forest. 2015, 7, 36–47. [Google Scholar]
- Torabian, S.; Zahedi, M.; Khoshgoftarmanesh, A. Effect of Foliar Spray of Zinc Oxide on Some Antioxidant Enzymes Activity of Sunflower under Salt Stress. J. Agric. Sci. Technol. 2016, 18, 1013–1025. [Google Scholar]
- Hussein, M.M.; Abou-Baker, N.H. The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 2018, 5, 171809. [Google Scholar] [CrossRef] [Green Version]
- Mahmoodzadeh, H.; Nabavi, M.; Kashefi, H. Effect of nanoscale titanium dioxide particles on the germination and growth of canola (Brassica napus). J. Ornament. Hortic. Plants 2013, 3, 25–32. [Google Scholar]
- Dimpka, C.O.; Singh, U.; Bindraban, P.S.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Zinc oxide nanoparticles alleviate drought-induced alterations in sorghum performance nutrient acquisition and grain fortification. Sci. Tot. Environ. 2019, 20, 926–934. [Google Scholar]
- Nedelkoska, T.V.; Doran, P.M. Characteristics of metal uptake by plants species with potential for phytoremediation and phytomining. Miner. Eng. 2000, 13, 549–561. [Google Scholar] [CrossRef]
- Ngomsik, A.F.; Bee, A.; Draye, M.; Cote, G.; Cabuil, V. Magnetic nano- and microparticles for metal removal and environmental applications, a review. C. R. Chim. 2005, 8, 963–970. [Google Scholar] [CrossRef]
- Dickinson, M.; Scott, T.B. The application of zero-valent iron nanoparticles for the remediation of a uranium-contaminated waste effluent. J Hazard. Mater. 2010, 178, 171–179. [Google Scholar] [CrossRef]
- Cui, J.; Liu, T.; Li, F.; Yi, J.; Liu, C.; Yu, H. Silica nanoparticles alleviate cadmium toxicity in rice cells, mechanisms and size effects. Environ. Pollut. 2017, 228, 363–369. [Google Scholar]
- Wang, F.; Wang, M.; Liu, Z.; Han, T.; Ye, Y.; Gong, N.; Sun, J.; Zhu, C. Different responses of low grain-Cd-accumulating and high grain-Cd-accumulating rice cultivars to Cd stress. Plant Physiol. Biochem. 2015, 96, 261–269. [Google Scholar] [CrossRef]
- Liu, C.; Li, F.; Luo, C.; Liu, X.; Wang, S.; Liu, T.; Li, X. Foliar application of two silica sols reduced cadmium accumulation in rice grains. J. Hazard. Mat. 2009, 161, 1466–1472. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnO NPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings, a physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Faraz, A.; Mir, A.R.; Hayat, S. Role of Zinc Oxide Nanoparticles in Countering Negative Effects Generated by Cadmium in Lycopersicon esculentum. J. Plant Growth Regul. 2020, 40, 101–115. [Google Scholar] [CrossRef]
- Li, Z.; Huang, J. Effects of nanoparticle hydroxyapatite on growth and antioxidant system in pakchoi (Brassica chinensis L.) from cadmium-contaminated soil. J. Nanomater. 2014, 2014, 470962. [Google Scholar] [CrossRef] [Green Version]
- Park, C.M.; Wang, D.; Han, J.; Heo, J.; Su, C. Evaluation of the colloidal stability and adsorption performance of reduced graphene oxide–elemental silver/magnetite nanohybrids for selected toxic heavy metals in aqueous solutions. Appl. Surf. Sci. 2019, 471, 8–17. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Nanoparticles | Test Crop | Effect of Nanoparticles | Reference |
|---|---|---|---|
| ZnO | Cyamopsis tetragonoloba (Cluster bean) | Significant increase in chlorophyll content, leaf protein, and alkaline phosphate | [12] |
| Vigna radiata | Enhanced germination and growth | [13] | |
| Arachis hypogaea | Increased seed germination rate | [14] | |
| Cicer arientum | Enhanced weight of the plant | [15] | |
| Glycine max | Increased root growth and development | [16] | |
| CeO2 | Glycine max | Enhancement in plant growth and development | [17] |
| Triticum aestivum | Enhanced shoot growth, biomass, grain yield | [18] | |
| Zea mays | Increased stem and root growth | [16] | |
| Sorghum | Enhanced assimilation rates of carbon in leaf and seed yield | [19] | |
| Allium cepa | Improved growth, yield, and nutrient content | [20] | |
| Ag | Lolium multifolium | Enhanced plant growth and development | [21] |
| Eruca sativa | Increased root length | [21] | |
| Zea mays | Increased root length | [22] | |
| Oryza sativa | Enhanced root length | [23] | |
| SiO2 | Lycopersicum esculentum | Increased seed germination | [24] |
| Al2O3 | Triticum aestivum | Enhanced root growth | [25] |
| Glycine max | Improved survival and growth | [26] | |
| Zea mays | Root length augmented | [27] | |
| Glycine max | Significant enhancement in root length | [28] | |
| Raphanus sativus | Improved root growth | [27] | |
| TiO2 | Triticum aestivum | Increased root length | [29] |
| Rosa | Enhanced plant resistance to infection of fungi by changing endogenous hormonal content | [30] | |
| Spinacia oleracea | Enhanced growth | [31] | |
| Lemna minor | Improved the activities of CAT, SOD, and POX by eliminating accumulated ROS in plant | [32] | |
| Cicer arietinum | Altered redox status | [33] | |
| Spinacia oleracea | Dry weight and chlorophyll level increased | [34] | |
| Spinacia oleracea | Increased fresh and dry biomass | [31] | |
| Spinacia oleracea | Improved light absorbance and carbon dioxide assimilation | [35] | |
| Fe/Fe2O3 | Citrullus lanatus | Enhanced the activity of root, POX, SOD | [36] |
| Triticum aestivum | Enhanced seed germination rate and plant growth | [37] | |
| Triticum aestivum | Increased shoot and root biomass | [26] |
| Nanoparticles | Test Crop | Effect of Nanoparticles | Reference |
|---|---|---|---|
| TiO2 | Triticum aestivum L. | Altered growth, yield, and starch level in drought stress | [103] |
| Oryza sativa L | Altered Cd migration in the soil–rice system in Cd toxicity | [104] | |
| Oryza sativa L. | NPs decreased the Cd uptake in Cd toxicity | [105] | |
| Linum usitatissimum L. | Altered length, biomass, yield, and carotenoid level in drought stress | [106] | |
| Oryza sativa L. | Mitigated Cd toxicity by improving antioxidant enzymes activity | [94] | |
| Ocimum basilicum L. | Reduced toxicity caused by drought stress | [107] | |
| SiO2 | Lycopersicum esculentum L. | Reduced salinity stress and enhanced seed germination potential, root length, and dry weight | [24] |
| Solanum lycopersicum L. | Altered weight and photosynthetic attributes under salinity stress | [108] | |
| Ocimum basilicum L. | Ameliorated salt stress and increased fresh and dry weight, chlorophyll content, and proline content | [109] | |
| Lens culinaris Medik | Altered seed germination rate and plant growth under salinity stress | [110] | |
| Cucurbita pepo L. | Increased activities of CAT, POX, and SOD | [111] | |
| ZnO | Moringa peregrina | Decreased salt stress and reduction in Na+ and Cl− contents | [112] |
| Helianthus annuus L. | Reduced salinity stress, increasing net CO2 assimilation rate | [113] | |
| Gossypium barbadense L. | Increased growth, yield, and mineral contents in salinity stress | [114] | |
| Helianthus annuus L. | Reduced salinity stress, increased plant growth, net CO2 assimilation | [113] | |
| Glycine max | Mitigate drought stress and increasing seed germination rate | [115] | |
| Sorghum bicolor | Enhanced drought tolerance, accelerated plant development and promoted yield | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faizan, M.; Alam, P.; Rajput, V.D.; Faraz, A.; Afzal, S.; Ahmed, S.M.; Yu, F.-Y.; Minkina, T.; Hayat, S. Nanoparticle Mediated Plant Tolerance to Heavy Metal Stress: What We Know? Sustainability 2023, 15, 1446. https://doi.org/10.3390/su15021446
Faizan M, Alam P, Rajput VD, Faraz A, Afzal S, Ahmed SM, Yu F-Y, Minkina T, Hayat S. Nanoparticle Mediated Plant Tolerance to Heavy Metal Stress: What We Know? Sustainability. 2023; 15(2):1446. https://doi.org/10.3390/su15021446
Chicago/Turabian StyleFaizan, Mohammad, Pravej Alam, Vishnu D. Rajput, Ahmad Faraz, Shadma Afzal, S. Maqbool Ahmed, Fang-Yuan Yu, Tatiana Minkina, and Shamsul Hayat. 2023. "Nanoparticle Mediated Plant Tolerance to Heavy Metal Stress: What We Know?" Sustainability 15, no. 2: 1446. https://doi.org/10.3390/su15021446
APA StyleFaizan, M., Alam, P., Rajput, V. D., Faraz, A., Afzal, S., Ahmed, S. M., Yu, F. -Y., Minkina, T., & Hayat, S. (2023). Nanoparticle Mediated Plant Tolerance to Heavy Metal Stress: What We Know? Sustainability, 15(2), 1446. https://doi.org/10.3390/su15021446