
Different Contribution of Olive Groves and Citrus Orchards to Soil Organic Carbon Sequestration: A Field Study in Four Sites in Crete, Greece


 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
- Farming land use either with olive or citrus trees will increase the soil organic C potential due to litter fall and rhizodeposition (will be indicated as a difference between tree crop species cultivations and non-cultivated fields in four different sites).
- Different tree crop species will have different soil organic C sequestration potentials due to their distinct physiological and morphological characteristics (e.g., litter quantity and quality and root distribution and rhizodeposition) and expected crop-specific soil microbiota communities and activities involved in C and nutrient cycling in the soil.
- Sites will affect the storage of soil organic C due to site-specific adjustment of crops due to existing soil-climatic conditions and site-specific soil microorganisms and their responses to environmental conditions.
- Soil organic C concentration could be correlated with specific soil properties and plant species characteristics (i.e., litter C:N ratio). Both the latter factors could affect organic C retention and availability and shape soil microbiota, consequently impacting the SOM storage in the soil.
2. Materials and Methods
2.1. Site Descriptions
2.2. Field Samplings and Analyses
2.3. Statistical Analysis
3. Results
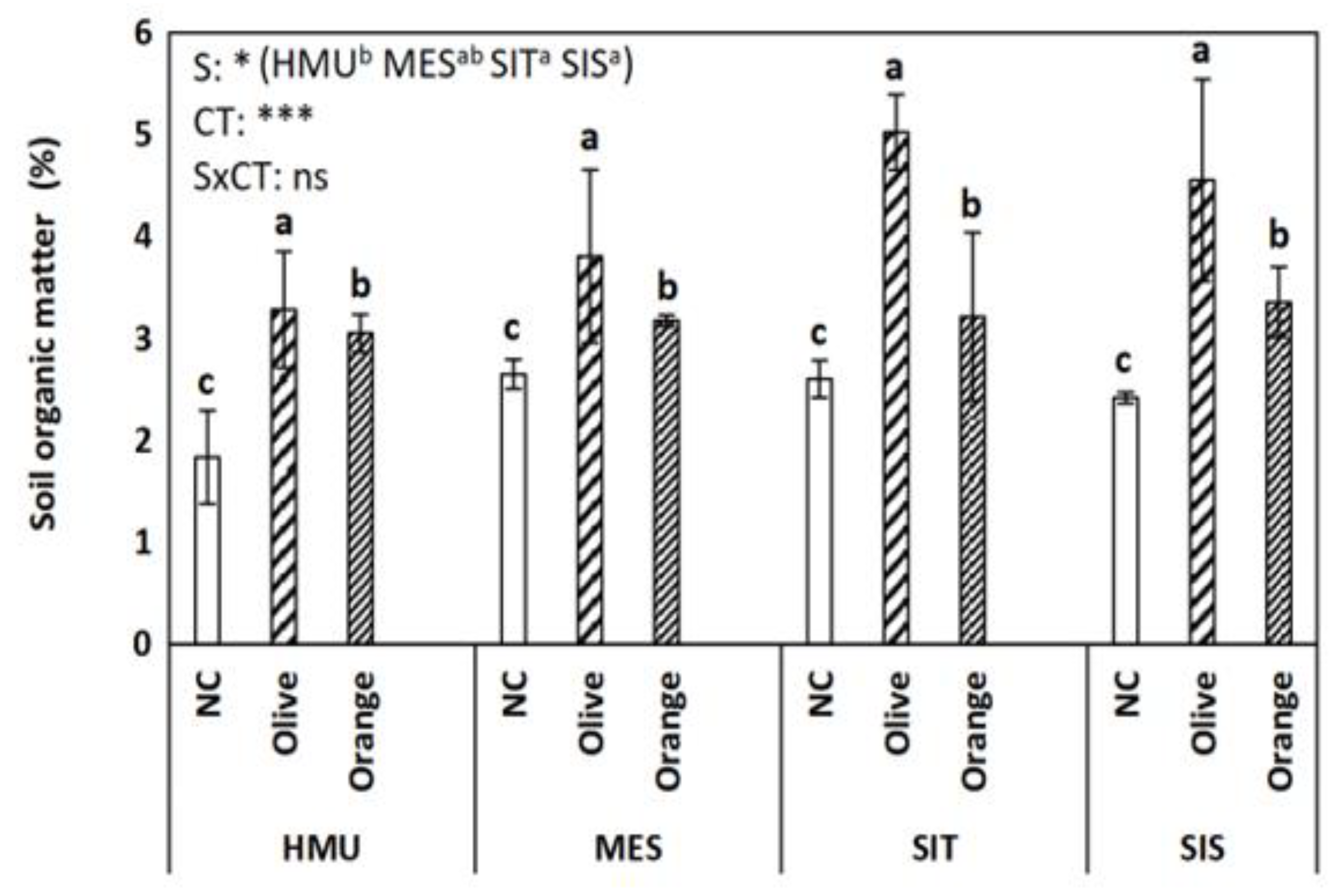
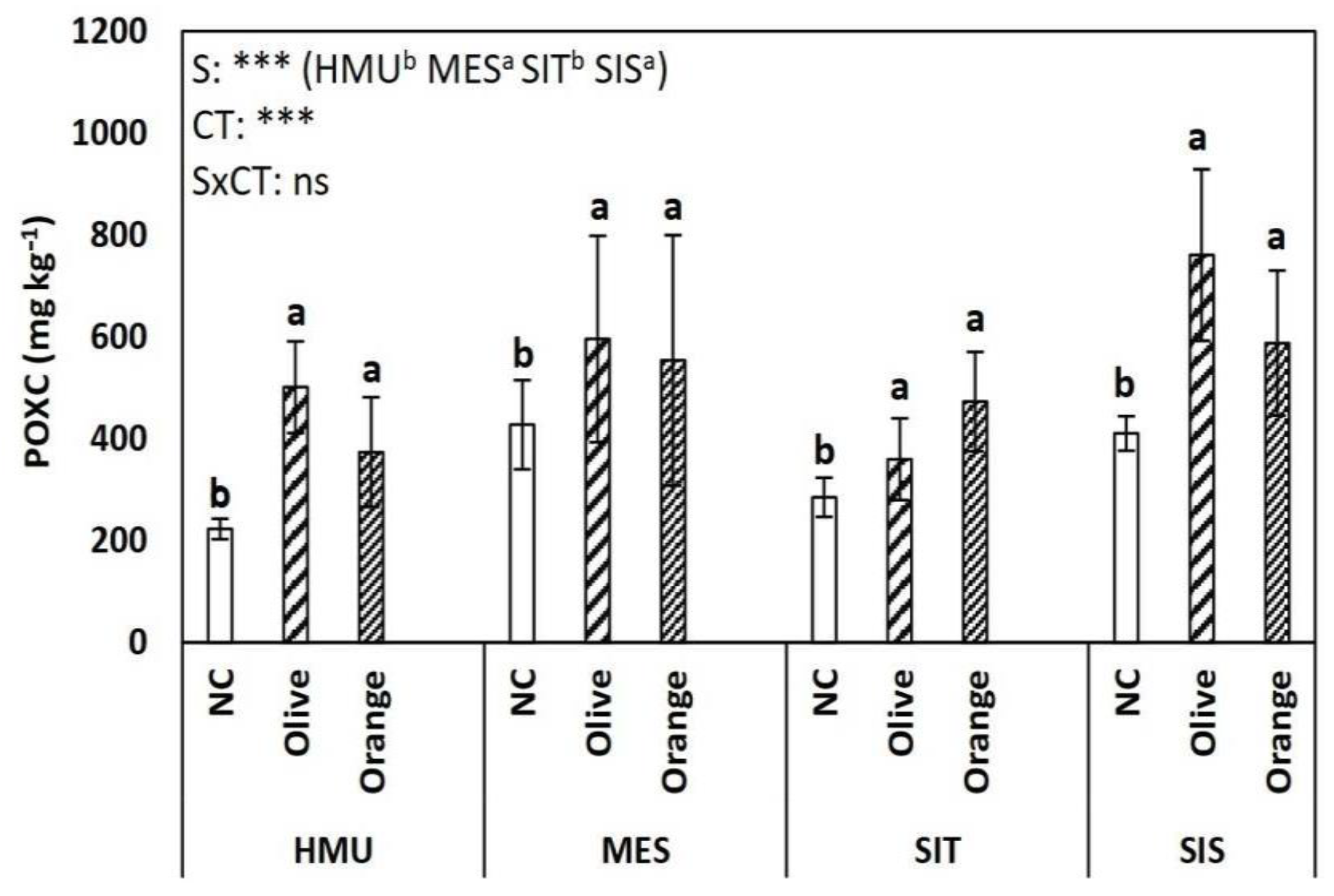
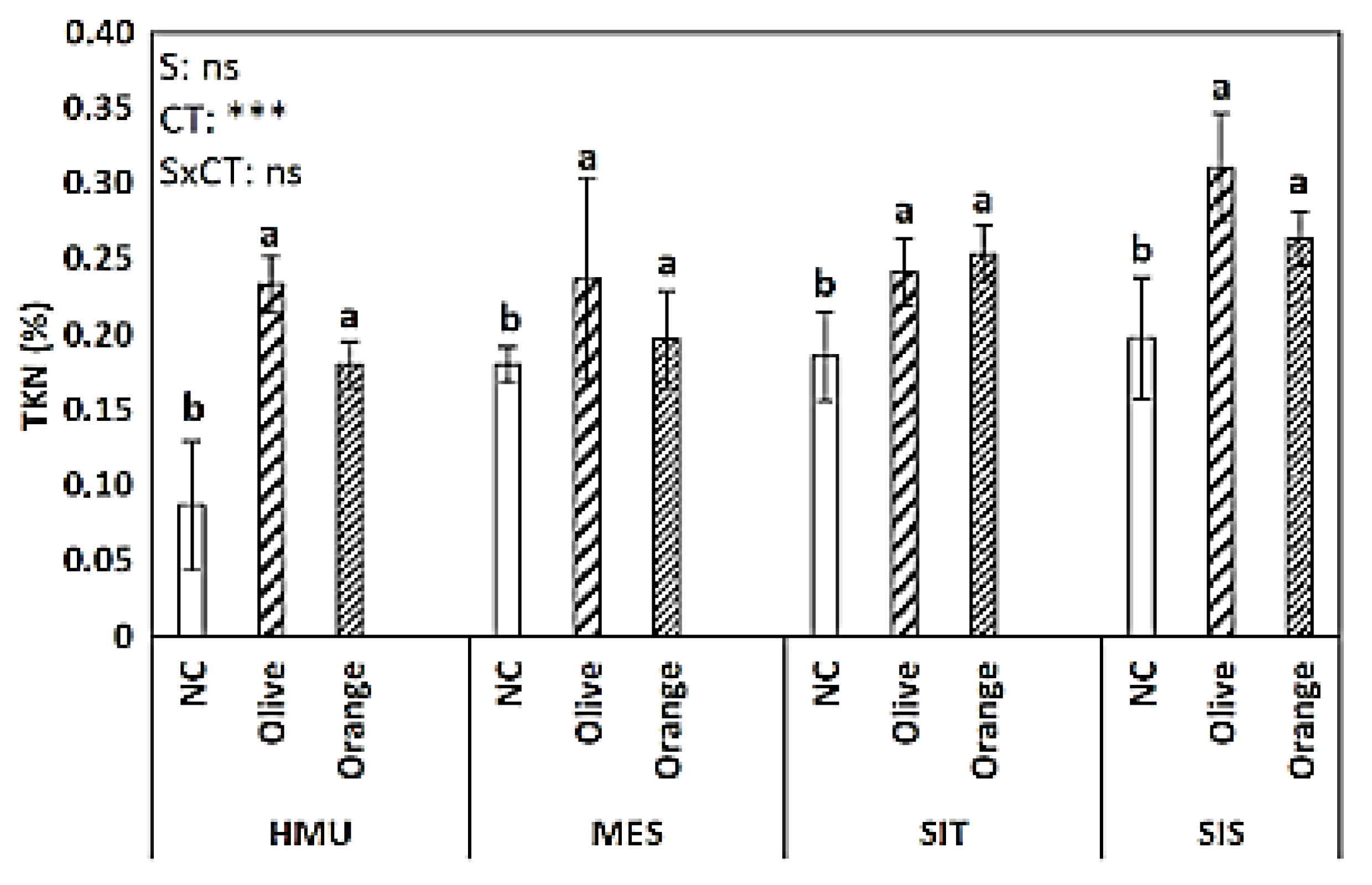
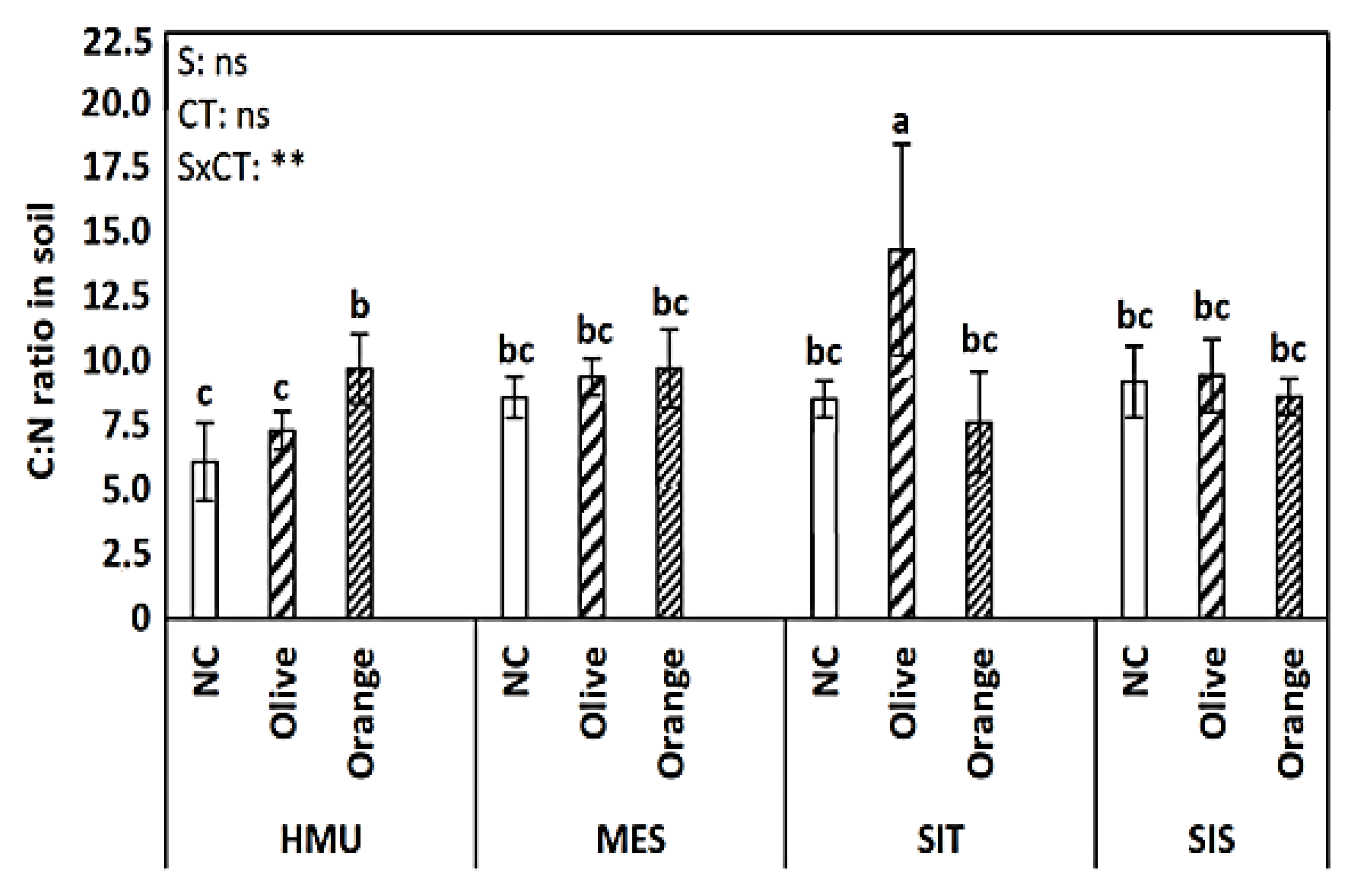
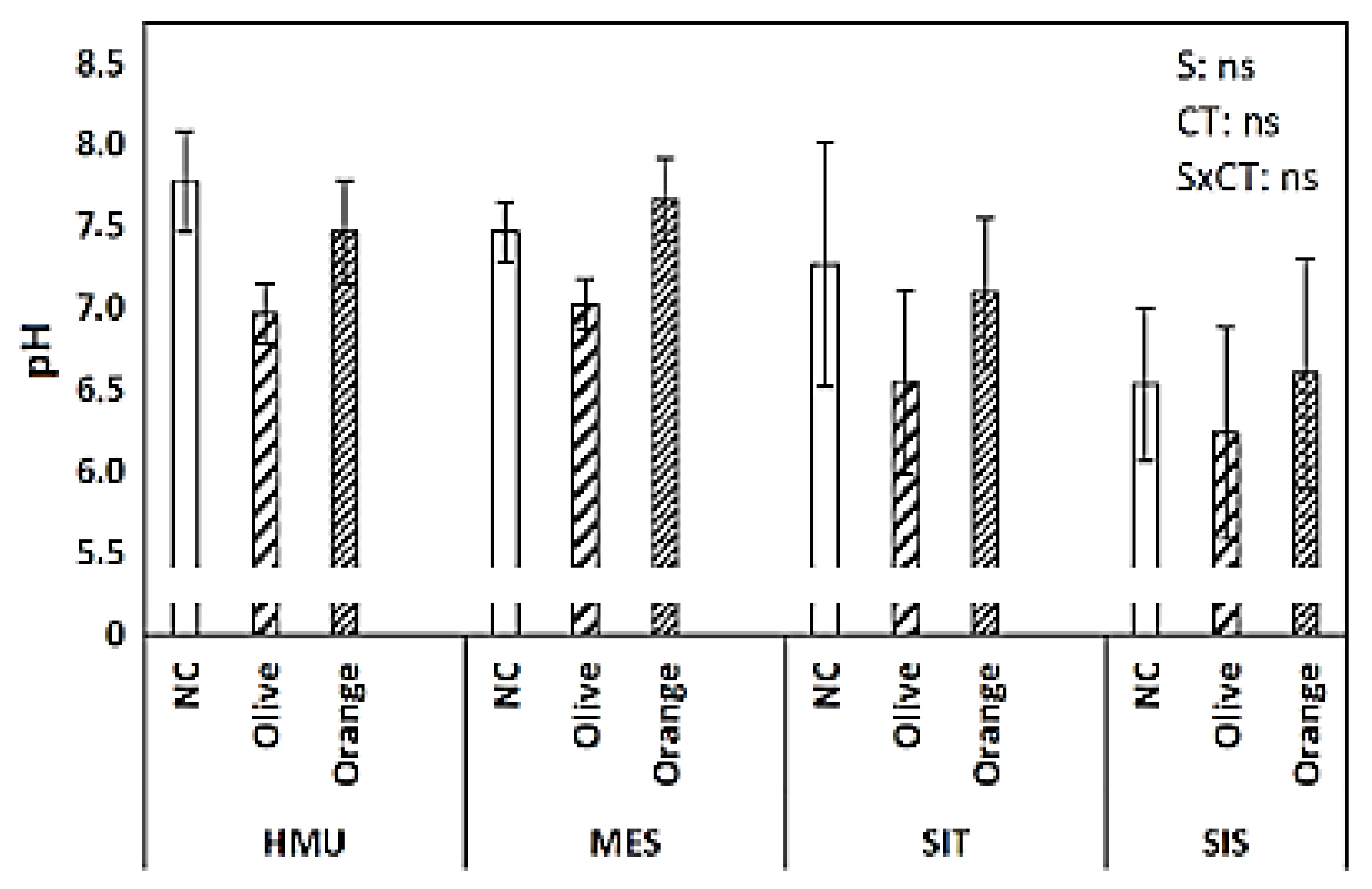
3.1. Soil Organic Matter (SOM), Permanganate Oxidizable Carbon (POXC), Total Kjeldahl Nitrogen (TKN), C:N Ratio, and pH
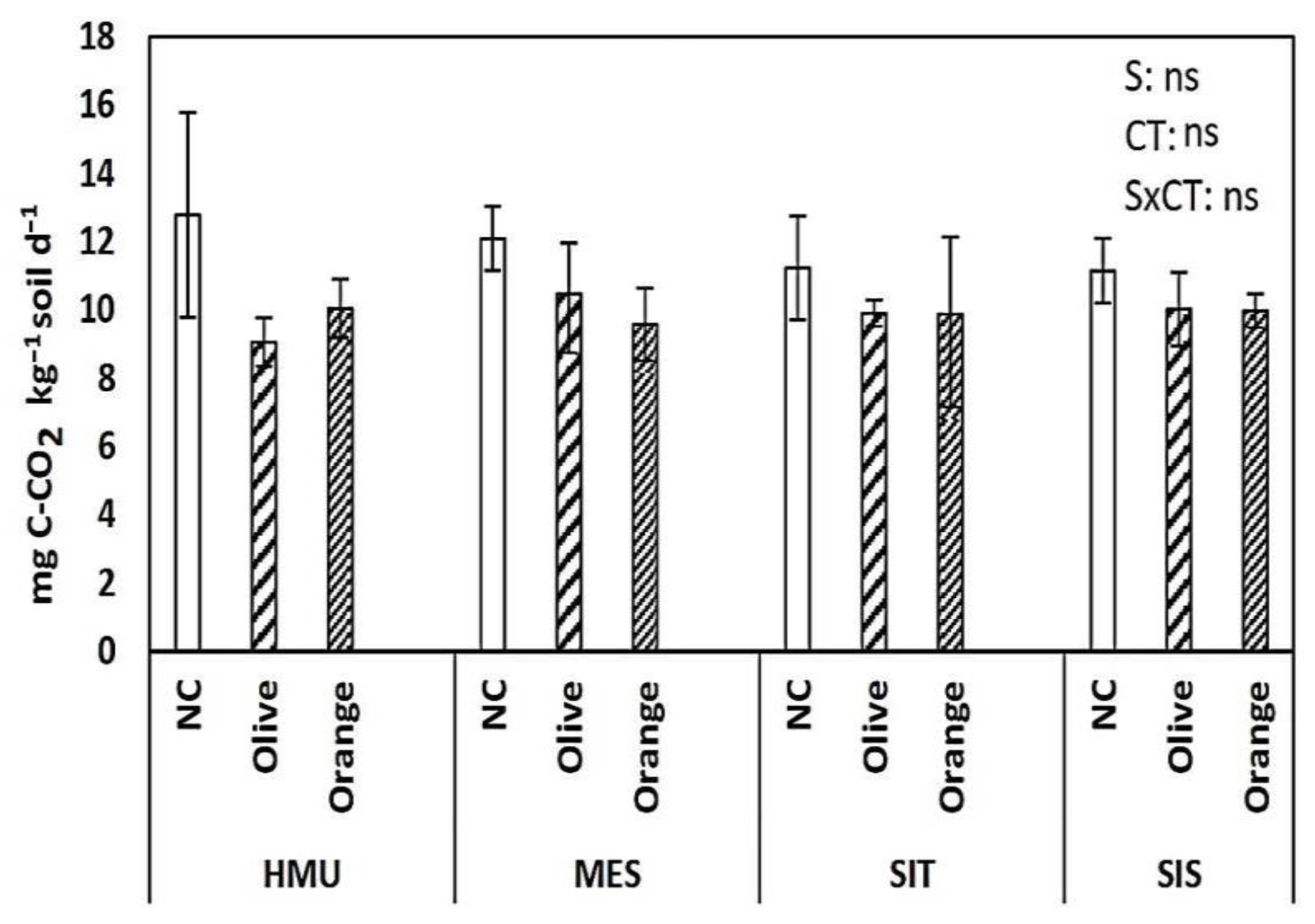
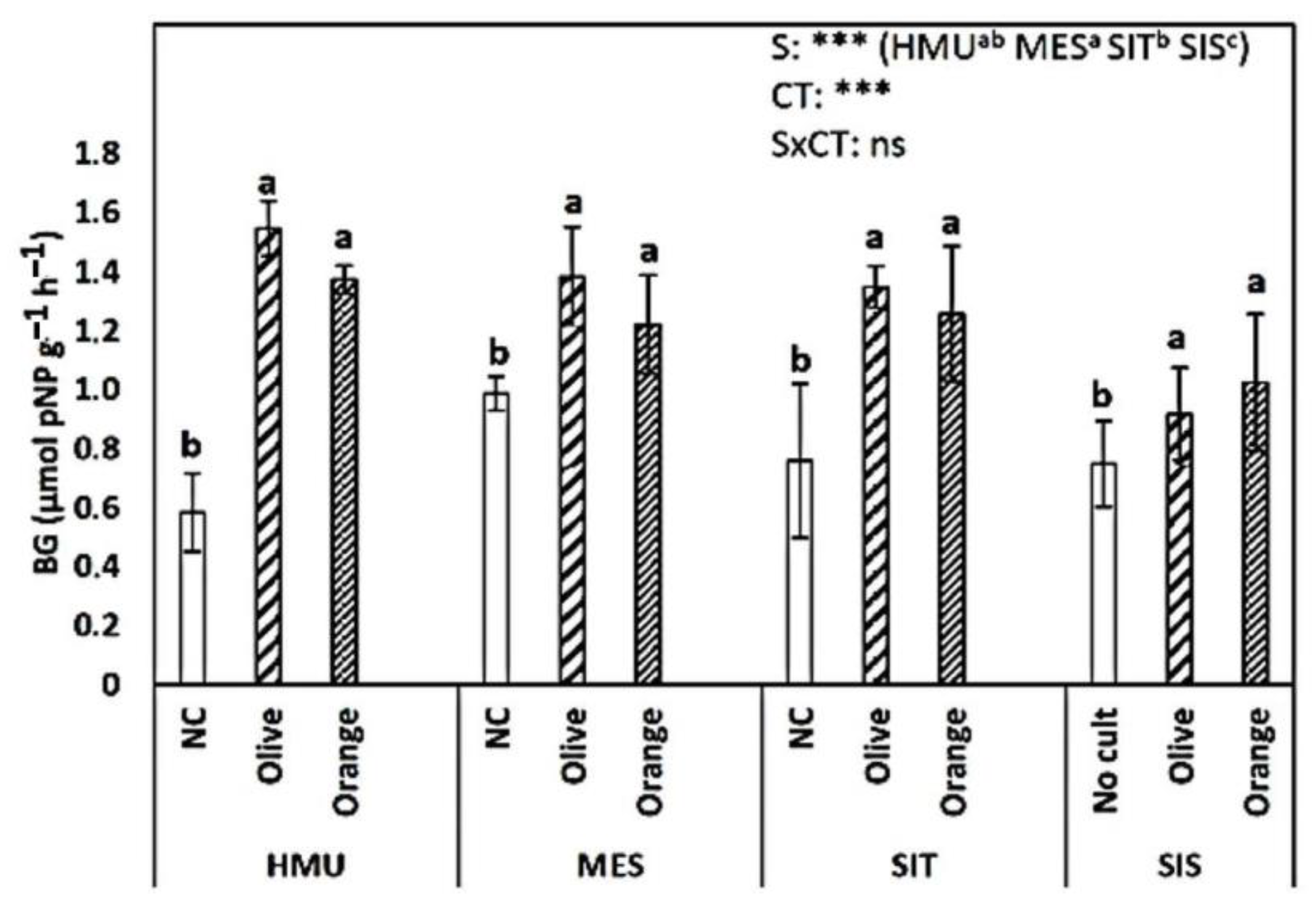
3.2. Respiration rates
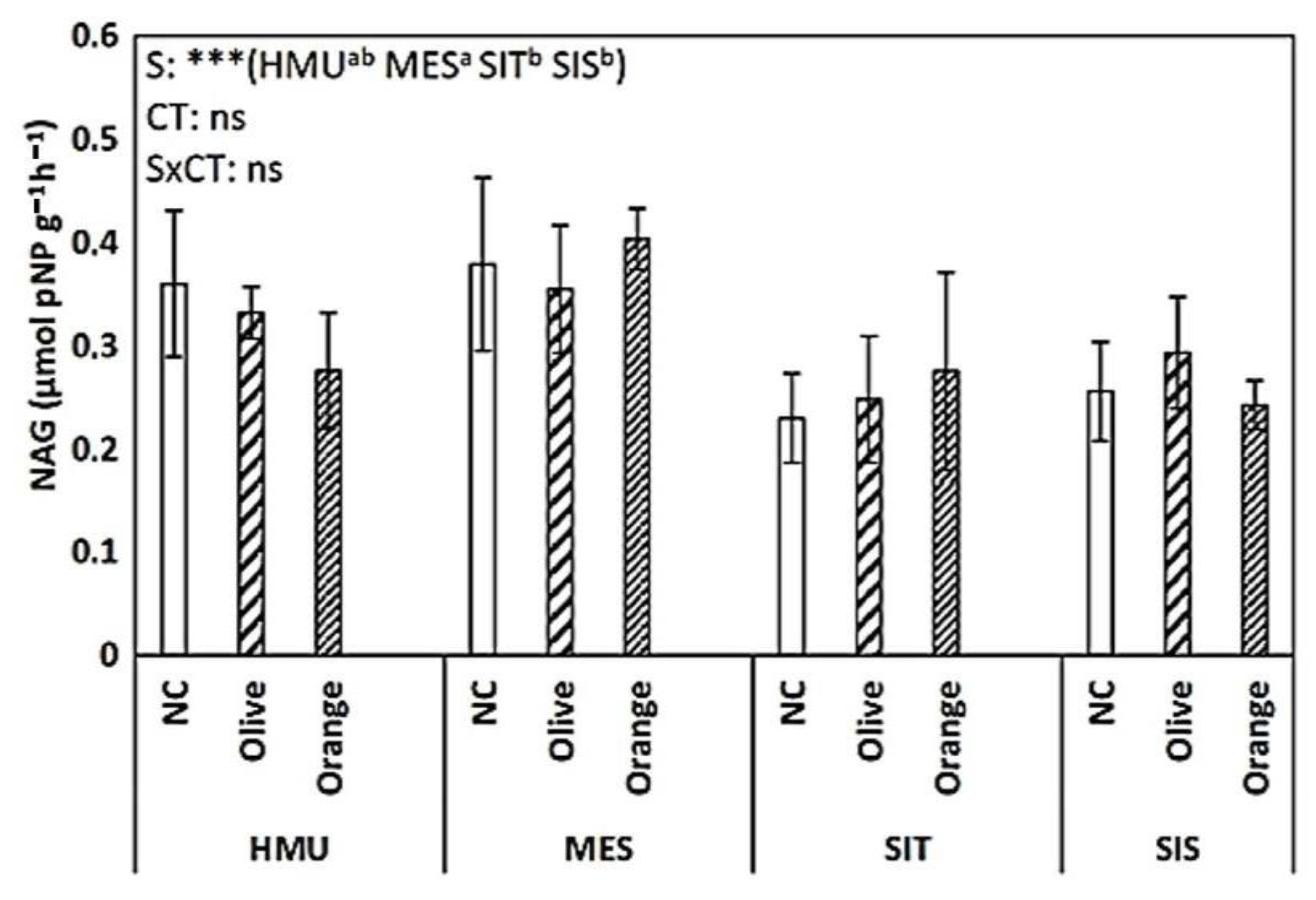
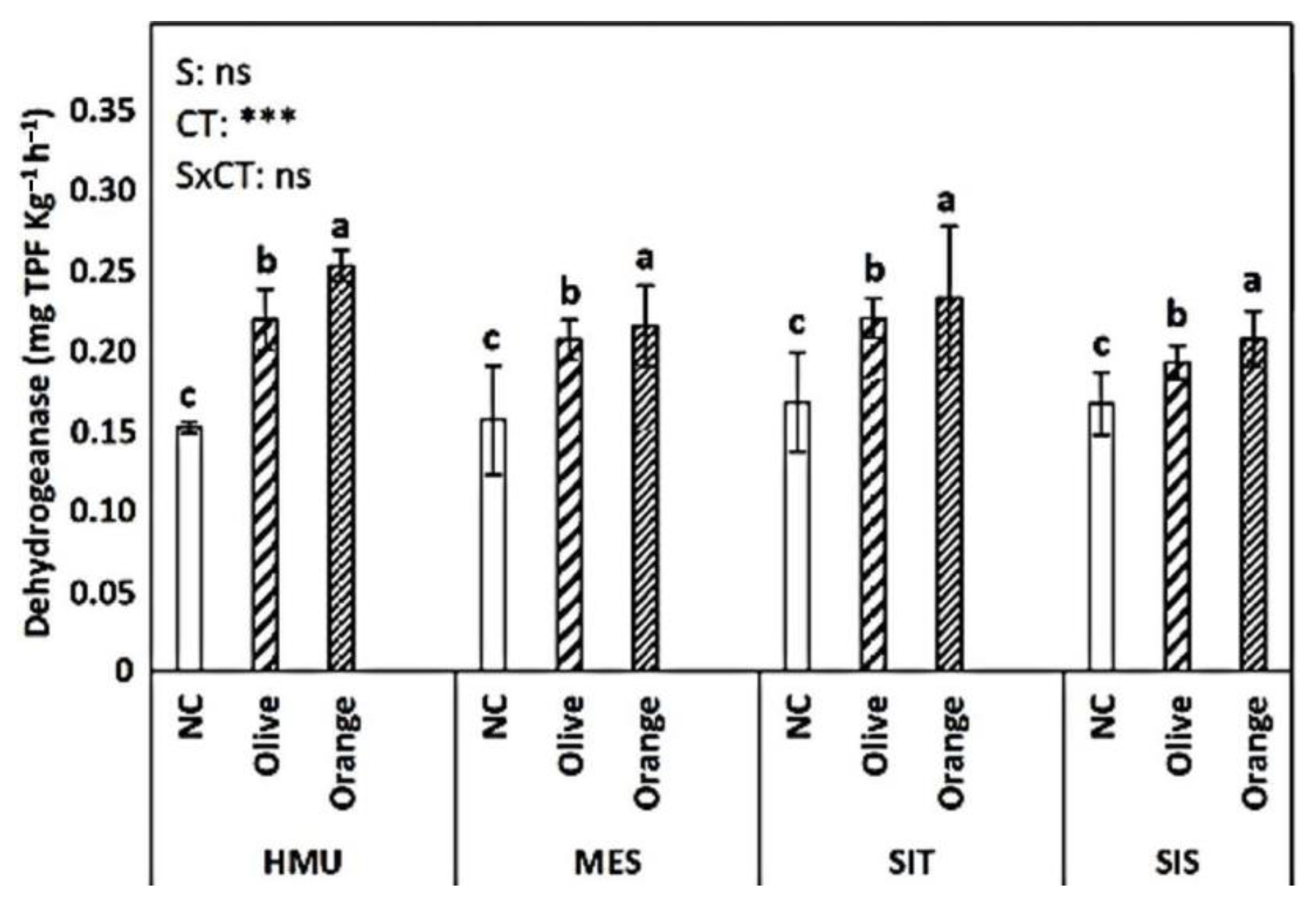
3.3. Enzyme Activities
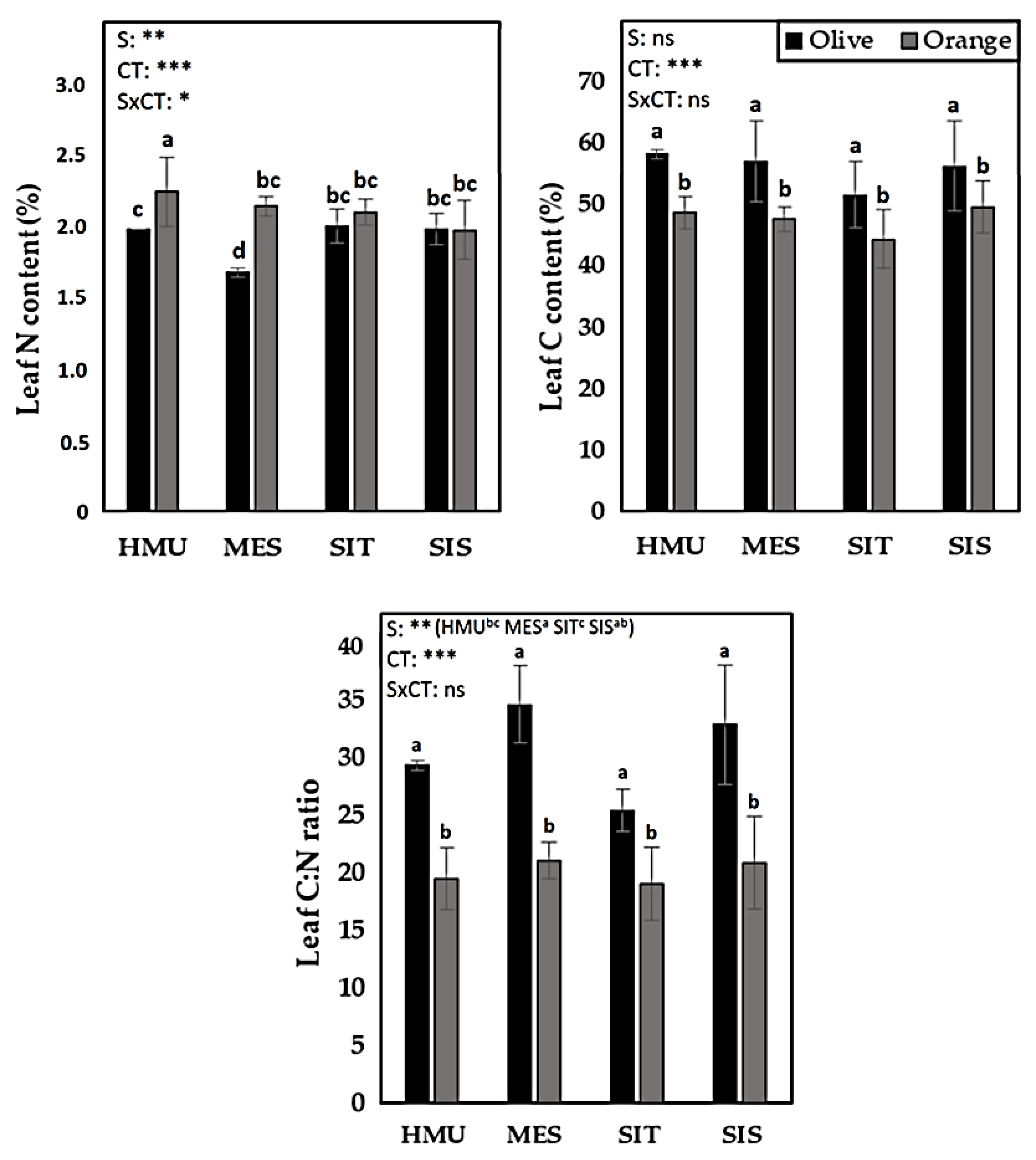
3.4. Carbon, Nitrogen, and C:N Content in Leaves
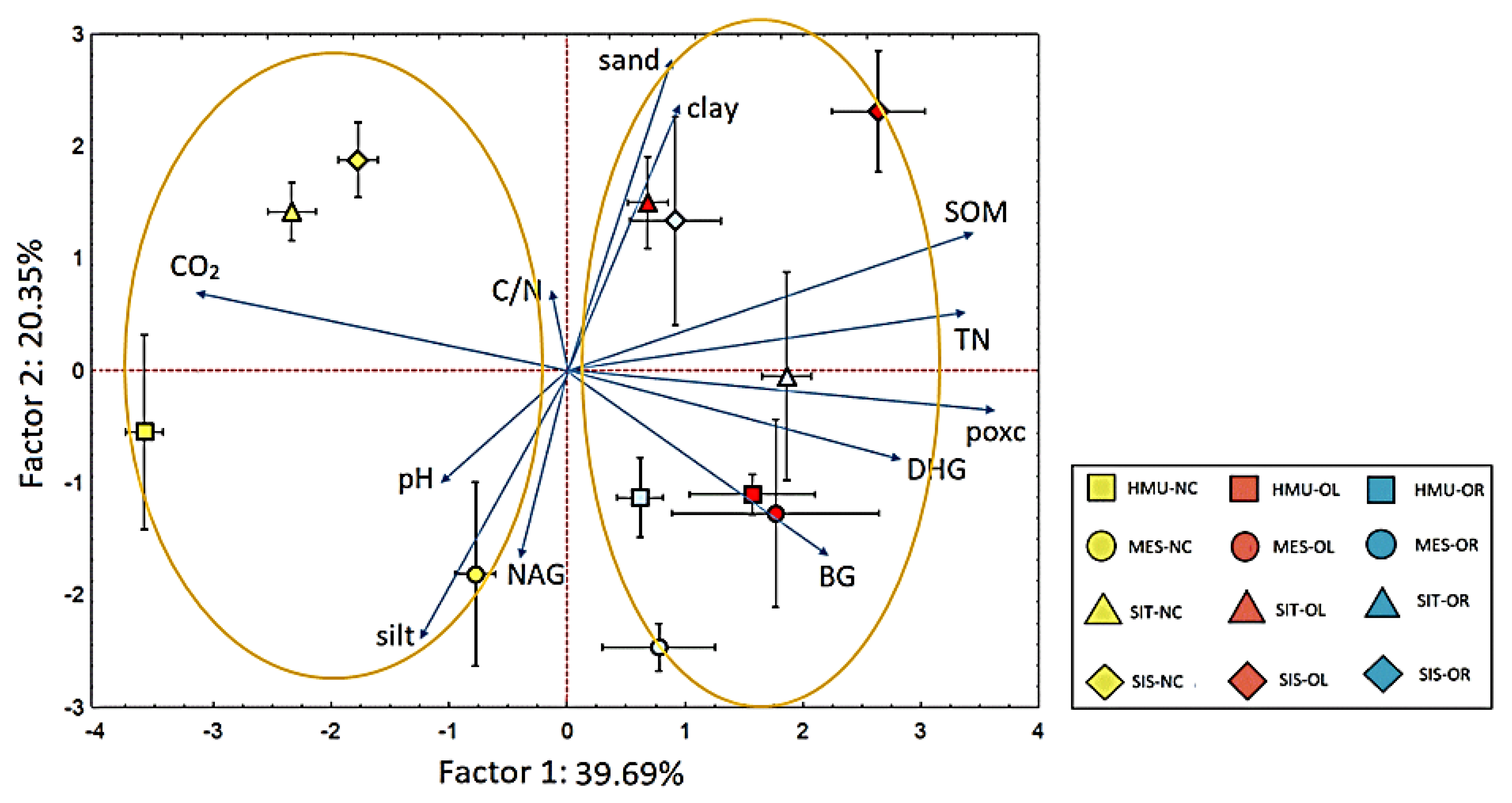
3.5. PCA and Correlation Analyses
4. Discussion
4.1. Tree Crop Species Contributed to Organic C Sequestration as Compared to Uncultivated Fields
4.2. Different Contributions to SOM Enhancement between Different Tree Crop Species
4.3. Limited Effect of Site Position on Soil Organic C Sequestration
4.4. Identifying Soil and Plant Parameters Representing Best Soil Organic C Sequestration
5. Conclusions
- Tree cropping systems (olive groves and orange orchards) can enhance organic C stock in the soil under the semi-arid soil-climatic conditions of Crete, Greece.
- Tree crops have different soil organic C sequestration potentials related to crop litter characteristics (e.g., leaf C:N ratio) and rhizosphere regulation. Olive trees showed higher soil organic C sequestration potential than orange trees.
- SOM is challenging to be described by typical soil properties due to the synergetic effect of a plethora of abiotic and biotic parameters; in our study, SOM had a positive relationship with TKN (and less with POXC) in the soil as well as with the C:N ratio of the tree litter.
- Further work should involve a more extensive survey and a more comprehensive range of plant and soil physicochemical (e.g., soil fractions) and microbial (e.g., microbial composition and structure) and biochemical soil parameters to identify the most critical ones to soil C sequestration. Another area of future research is the investigation of the coupling between the C and N cycles in semi-arid agroecosystems.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lal, R.; Negassa, W.; Lorenz, K. Carbon sequestration in soil. Curr. Opin. Environ. Sustain. 2015, 15, 79–86. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Change 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, E.; Diaz-Gaona, C.; Garcia-Laureano, R.; Reyes-Palomo, C.; Guzmán, G.I.; Ortolani, L.; Sanchez-Rodriguez, M.; Rodriguez-Estevez, V. Agroecology for adaptation to climate change and resource depletion in the Mediterranean region. A review. Agric. Syst. 2020, 181, 102809. [Google Scholar] [CrossRef]
- Fisher, J.B.; Melton, F.; Middleton, E.; Hain, C.; Anderson, M.; Allen, R.; McCabe, M.F.; Hook, S.; Baldocchi, D.; Townsend, P.A. The future of evapotranspiration: Global requirements for ecosystem functioning, carbon and climate feedbacks, agricultural management, and water resources. Water Resour. Res. 2017, 53, 2618–2626. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Meena, R.S.; Jhariya, M.K.; Yadav, D.K. Agroecological Footprints Management for Sustainable Food System; Springer: Berlin, Germany, 2021. [Google Scholar]
- Meena, R.S.; Kumar, S.; Yadav, G.S. Soil carbon sequestration in crop production. In Nutrient Dynamics for Sustainable Crop Production; Springer: Berlin, Germany, 2020; pp. 1–39. [Google Scholar]
- Paranychianakis, N.V.; Giannakis, G.; Moraetis, D.; Tzanakakis, V.A.; Nikolaidis, N.P. Crop litter has a strong effect on soil organic matter sequestration in semi-arid environments. Sustainability 2021, 13, 13278. [Google Scholar] [CrossRef]
- Pausch, J.; Kuzyakov, Y. Carbon input by roots into the soil: Quantification of rhizodeposition from root to ecosystem scale. Glob. Change Biol. 2018, 24, 1–12. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Cornelissen, J.H.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Rolston, D.E.; Horwath, W.R. Cover cropping affects soil N2O and CO2 emissions differently depending on type of irrigation. Agric. Ecosyst. Environ. 2010, 137, 251–260. [Google Scholar] [CrossRef]
- Badagliacca, G.; Ruisi, P.; Rees, R.M.; Saia, S. An assessment of factors controlling N2O and CO2 emissions from crop residues using different measurement approaches. Biol. Fertil. Soils 2017, 53, 547–561. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.; Al-Kaisi, M. Crop rotation and nitrogen fertilization effect on soil CO2 emissions in central Iowa. Appl. Soil Ecol. 2008, 39, 264–270. [Google Scholar] [CrossRef]
- Sun, W.; Canadell, J.G.; Yu, L.; Yu, L.; Zhang, W.; Smith, P.; Fischer, T.; Huang, Y. Climate drives global soil carbon sequestration and crop yield changes under conservation agriculture. Glob. Change Biol. 2020, 26, 3325–3335. [Google Scholar] [CrossRef]
- Jarecki, M.K.; Lal, R. Crop management for soil carbon sequestration. Crit. Rev. Plant Sci. 2003, 22, 471–502. [Google Scholar] [CrossRef]
- Langgut, D.; Cheddadi, R.; Carrión, J.S.; Cavanagh, M.; Colombaroli, D.; Eastwood, W.J.; Greenberg, R.; Litt, T.; Mercuri, A.M.; Miebach, A. The origin and spread of olive cultivation in the Mediterranean Basin: The fossil pollen evidence. Holocene 2019, 29, 902–922. [Google Scholar] [CrossRef]
- Proietti, S.; Sdringola, P.; Desideri, U.; Zepparelli, F.; Brunori, A.; Ilarioni, L.; Nasini, L.; Regni, L.; Proietti, P. Carbon footprint of an olive tree grove. Appl. Energy 2014, 127, 115–124. [Google Scholar] [CrossRef]
- Sofo, A.; Nuzzo, V.; Palese, A.M.; Xiloyannis, C.; Celano, G.; Zukowskyj, P.; Dichio, B. Net CO2 storage in Mediterranean olive and peach orchards. Sci. Hortic. 2005, 107, 17–24. [Google Scholar] [CrossRef]
- Testi, L.; Orgaz, F.; Villalobos, F. Carbon exchange and water use efficiency of a growing, irrigated olive orchard. Environ. Exp. Bot. 2008, 63, 168–177. [Google Scholar] [CrossRef]
- Villalobos, F.; Testi, L.; Hidalgo, J.; Pastor, M.; Orgaz, F. Modelling potential growth and yield of olive (Olea europaea L.) canopies. Eur. J. Agron. 2006, 24, 296–303. [Google Scholar] [CrossRef]
- López-Bellido, R.J.; Fontán, J.M.; López-Bellido, F.J.; López-Bellido, L. Carbon sequestration by tillage, rotation, and nitrogen fertilization in a Mediterranean Vertisol. Agron. J. 2010, 102, 310–318. [Google Scholar] [CrossRef]
- Zapata-Sierra, A.J.; Manzano-Agugliaro, F. Controlled deficit irrigation for orange trees in Mediterranean countries. J. Clean. Prod. 2017, 162, 130–140. [Google Scholar] [CrossRef]
- Yang, S.; Aydin, M.; Kitamura, Y.; Yano, T. The impact of irrigation water quality on water uptake by orange trees. Afr. J. Agric. Res. 2010, 5, 2661–2667. [Google Scholar]
- Germana, C.; Sardo, V. Determining when to initiate irrigation of orange trees. In Proceedings of the IV International Symposium on Irrigation of Horticultural Crops 664, Davis, CA, USA, 1–6 September 2003; pp. 591–597. [Google Scholar]
- Consoli, S.; Stagno, F.; Roccuzzo, G.; Cirelli, G.; Intrigliolo, F. Sustainable management of limited water resources in a young orange orchard. Agric. Water Manag. 2014, 132, 60–68. [Google Scholar] [CrossRef]
- Consoli, S.; Stagno, F.; Vanella, D.; Boaga, J.; Cassiani, G.; Roccuzzo, G. Partial root-zone drying irrigation in orange orchards: Effects on water use and crop production characteristics. Eur. J. Agron. 2017, 82, 190–202. [Google Scholar] [CrossRef]
- Tsanis, I.K.; Koutroulis, A.G.; Daliakopoulos, I.N.; Jacob, D. Severe climate-induced water shortage and extremes in Crete. Clim. Chang. 2011, 106, 667–677. [Google Scholar] [CrossRef]
- Tzanakakis, V.; Angelakis, A.; Paranychianakis, N.; Dialynas, Y.; Tchobanoglous, G. Challenges and Opportunities for Sustainable Management of Water Resources in the Island of Crete, Greece. Water 2020, 12, 1538. [Google Scholar] [CrossRef]
- Allen, H.; Randall, R.; Amable, G.; Devereux, B. The impact of changing olive cultivation practices on the ground flora of olive groves in the Messara and Psiloritis regions, Crete, Greece. Land Degrad. Dev. 2006, 17, 249–273. [Google Scholar] [CrossRef]
- Goward, J.; Whitty, M.; Ch, R. Estimating and predicting carbon sequestrated in a vineyard with soil surveys, spatial date and G.I.S. management. Bachelor’s Thesis, Engineering University of New South Wales, Kensington, Australia, 2012. [Google Scholar]
- Lal, R. Soil carbon stocks under present and future climate with specific reference to European ecoregions. Nutr. Cycl. Agroecosystems 2008, 81, 113–127. [Google Scholar] [CrossRef]
- Proietti, P.; Sdringola, P.; Brunori, A.; Ilarioni, L.; Nasini, L.; Regni, L.; Pelleri, F.; Desideri, U.; Proietti, S. Assessment of carbon balance in intensive and extensive tree cultivation systems for oak, olive, poplar and walnut plantation. J. Clean. Prod. 2016, 112, 2613–2624. [Google Scholar] [CrossRef]
- Nair, P.R.; Nair, V.D.; Kumar, B.M.; Showalter, J.M. Carbon sequestration in agroforestry systems. Adv. Agron. 2010, 108, 237–307. [Google Scholar]
- Kourgialas, N.N.; Anyfanti, I.; Karatzas, G.P.; Dokou, Z. An integrated method for assessing drought prone areas—Water efficiency practices for a climate resilient Mediterranean agriculture. Sci. Total Environ. 2018, 625, 1290–1300. [Google Scholar] [CrossRef]
- Tzoraki, O.; Kritsotakis, M.; Baltas, E. Spatial Water Use efficiency Index towards resource sustainability: Application in the island of Crete, Greece. Int. J. Water Resour. Dev. 2015, 31, 669–681. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils 1. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Nelson, D.a.; Sommers, L.E. Total carbon, organic carbon, and organic matter. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1983, 9, 539–579. [Google Scholar]
- Bremner, J.M. Nitrogen-total. Methods Soil Anal. Part 3 Chem. Methods 1996, 5, 1085–1121. [Google Scholar]
- Weil, R.R.; Islam, K.R.; Stine, M.A.; Gruver, J.B.; Samson-Liebig, S.E. Estimating active carbon for soil quality assessment: A simplified method for laboratory and field use. Am. J. Altern. Agric. 2003, 18, 3–17. [Google Scholar]
- Allison, S.D.; Jastrow, J.D. Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol. Biochem. 2006, 38, 3245–3256. [Google Scholar] [CrossRef]
- Bottomley, P.J.; Angle, J.S.; Weaver, R. Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties; John Wiley & Sons: New York, NY, USA, 2020; Volume 12. [Google Scholar]
- Kapagianni, P.D.; Topalis, I.; Gwynn-Jones, D.; Menkissoglu-Spiroudi, U.; Stamou, G.P.; Papatheodorou, E.M. Effects of plant invaders on rhizosphere microbial attributes depend on plant identity and growth stage. Soil Res. 2020, 59, 225–238. [Google Scholar] [CrossRef]
- Garcia-Franco, N.; Wiesmeier, M.; Colocho Hurtarte, L.C.; Fella, F.; Martínez-Mena, M.; Almagro, M.; Martínez, E.G.; Kögel-Knabner, I. Pruning residues incorporation and reduced tillage improve soil organic matter stabilization and structure of salt-affected soils in a semi-arid Citrus tree orchard. Soil Tillage Res. 2021, 213, 105129. [Google Scholar] [CrossRef]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef]
- Benizri, E.; Dedourge, O.; Dibattista-Leboeuf, C.; Piutti, S.; Nguyen, C.; Guckert, A. Effect of maize rhizodeposits on soil microbial community structure. Appl. Soil Ecol. 2002, 21, 261–265. [Google Scholar] [CrossRef]
- Semchenko, M.; Xue, P.; Leigh, T. Functional diversity and identity of plant genotypes regulate rhizodeposition and soil microbial activity. New Phytol. 2021, 232, 776–787. [Google Scholar] [CrossRef]
- Tamura, M.; Suseela, V.; Simpson, M.; Powell, B.; Tharayil, N. Plant litter chemistry alters the content and composition of organic carbon associated with soil mineral and aggregate fractions in invaded ecosystems. Glob. Chang. Biol. 2017, 23, 4002–4018. [Google Scholar] [CrossRef]
- Castellano, M.J.; Mueller, K.E.; Olk, D.C.; Sawyer, J.E.; Six, J. Integrating plant litter quality, soil organic matter stabilization, and the carbon saturation concept. Glob. Chang. Biol. 2015, 21, 3200–3209. [Google Scholar] [CrossRef] [Green Version]
- Redin, M.; Guénon, R.; Recous, S.; Schmatz, R.; de Freitas, L.L.; Aita, C.; Giacomini, S.J. Carbon mineralization in soil of roots from twenty crop species, as affected by their chemical composition and botanical family. Plant Soil 2014, 378, 205–214. [Google Scholar] [CrossRef]
- Talbot, J.M.; Yelle, D.J.; Nowick, J.; Treseder, K.K. Litter decay rates are determined by lignin chemistry. Biogeochemistry 2012, 108, 279–295. [Google Scholar] [CrossRef]
- Forney, D.C.; Rothman, D.H. Common structure in the heterogeneity of plant-matter decay. J. R. Soc. Interface 2012, 9, 2255–2267. [Google Scholar] [CrossRef]
- Rahman, M.M.; Tsukamoto, J.; Rahman, M.M.; Yoneyama, A.; Mostafa, K.M. Lignin and its effects on litter decomposition in forest ecosystems. Chem. Ecol. 2013, 29, 540–553. [Google Scholar] [CrossRef]
- Giannakis, G.V.; Panakoulia, S.K.; Nikolaidis, N.P.; Paranychianakis, N.V. Simulating Soil Fertility Restoration Using the CAST Model. Procedia Earth Planet. Sci. 2014, 10, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Nieto, O.M.; Castro, J.; Fernández, E.; Smith, P. Simulation of soil organic carbon stocks in a Mediterranean olive grove under different soil-management systems using the RothC model. Soil Use Manag. 2010, 26, 118–125. [Google Scholar] [CrossRef]
- Tzanakakis, V.A.; Paranychianakis, N.V. Divergent response of ammonia oxidizers to various amino acids. Appl. Soil Ecol. 2017, 114, 45–51. [Google Scholar] [CrossRef]
- Qiu, H.; Ge, T.; Liu, J.; Chen, X.; Hu, Y.; Wu, J.; Su, Y.; Kuzyakov, Y. Effects of biotic and abiotic factors on soil organic matter mineralization: Experiments and structural modeling analysis. Eur. J. Soil Biol. 2018, 84, 27–34. [Google Scholar] [CrossRef]
- Cai, A.; Feng, W.; Zhang, W.; Xu, M. Climate, soil texture, and soil types affect the contributions of fine-fraction-stabilized carbon to total soil organic carbon in different land uses across China. J. Environ. Manag. 2016, 172, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Angst, G.; Messinger, J.; Greiner, M.; Häusler, W.; Hertel, D.; Kirfel, K.; Kögel-Knabner, I.; Leuschner, C.; Rethemeyer, J.; Mueller, C.W. Soil organic carbon stocks in topsoil and subsoil controlled by parent material, carbon input in the rhizosphere, and microbial-derived compounds. Soil Biol. Biochem. 2018, 122, 19–30. [Google Scholar] [CrossRef]
- Moinet, G.Y.; Hunt, J.E.; Kirschbaum, M.U.; Morcom, C.P.; Midwood, A.J.; Millard, P. The temperature sensitivity of soil organic matter decomposition is constrained by microbial access to substrates. Soil Biol. Biochem. 2018, 116, 333–339. [Google Scholar] [CrossRef]
- Goss-Souza, D.; Mendes, L.W.; Borges, C.D.; Baretta, D.; Tsai, S.M.; Rodrigues, J.L. Soil microbial community dynamics and assembly under long-term land use change. FEMS Microbiol. Ecol. 2017, 93, fix109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennon, J.T.; Aanderud, Z.T.; Lehmkuhl, B.K.; Schoolmaster, D.R., Jr. Mapping the niche space of soil microorganisms using taxonomy and traits. Ecology 2012, 93, 1867–1879. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialisation and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Mikutta, R.; Kleber, M.; Torn, M.S.; Jahn, R. Stabilization of soil organic matter: Association with minerals or chemical recalcitrance? Biogeochemistry 2006, 77, 25–56. [Google Scholar] [CrossRef]
- Hurisso, T.T.; Culman, S.W.; Horwath, W.R.; Wade, J.; Cass, D.; Beniston, J.W.; Bowles, T.M.; Grandy, A.S.; Franzluebbers, A.J.; Schipanski, M.E.; et al. Comparison of Permanganate-Oxidizable Carbon and Mineralizable Carbon for Assessment of Organic Matter Stabilization and Mineralization. Soil Sci. Soc. Am. J. 2016, 80, 1352–1364. [Google Scholar] [CrossRef]
- Sprunger, C.D.; Martin, T.; Mann, M. Systems with greater perenniality and crop diversity enhance soil biological health. Agric. Environ. Lett. 2020, 5, e20030. [Google Scholar] [CrossRef]
- Yao, Z.; Zhang, D.; Liu, N.; Yao, P.; Zhao, N.; Li, Y.; Zhang, S.; Zhai, B.; Huang, D.; Wang, Z.; et al. Dynamics and Sequestration Potential of Soil Organic Carbon and Total Nitrogen Stocks of Leguminous Green Manure-Based Cropping Systems on the Loess Plateau of China. Soil Tillage Res. 2019, 191, 108–116. [Google Scholar] [CrossRef]
- Tong, C.; Xiao, H.; Tang, G.; Wang, H.; Huang, T.; Xia, H.; Keith, S.J.; Li, Y.; Liu, S.; Wu, J. Long-term fertilizer effects on organic carbon and total nitrogen and coupling relationships of C and N in paddy soils in subtropical China. Soil Tillage Res. 2009, 106, 8–14. [Google Scholar] [CrossRef]
- Hamarashid, N.H.; Othman, M.A.; Hussain, M.-A.H. Effects of soil texture on chemical compositions, microbial populations and carbon mineralization in soil. Egypt. J. Exp. Biol.(Bot.) 2010, 6, 59–64. [Google Scholar]
- Augustin, C.; Cihacek, L.J. Relationships between soil carbon and soil texture in the Northern Great Plains. Soil Sci. 2016, 181, 386–392. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Silt (%) | Clay (%) | Sand (%) | Soil Texture | CaCO3 (%) | ||
|---|---|---|---|---|---|---|
| HMU | Non-cultivated | 71 | 3 | 26 | Silt Loam | 14 |
| Olive trees | 75 | 1 | 25 | Silt Loam | 14 | |
| Citrus trees | 74 | 3 | 23 | Silt Loam | 13 | |
| MES | Non-cultivated | 53 | 13 | 35 | Silt Loam | 17 |
| Olive trees | 55 | 4 | 41 | Silt Loam | 18 | |
| Citrus trees | 47 | 14 | 39 | Silt Loam | 14 | |
| SIT | Non-cultivated | 30 | 14 | 57 | Sandy Loam | <1 |
| Olive trees | 23 | 20 | 57 | Sandy Loam | <1 | |
| Citrus trees | 38 | 13 | 48 | Sandy Loam | <1 | |
| SIS | Non-cultivated | 33 | 8 | 59 | Sandy Loam | <1 |
| Olive trees | 41 | 7 | 51 | Sandy Loam | <1 | |
| Citrus trees | 36 | 9 | 55 | Sandy Loam | <1 |
| Silt | Clay | Sand | NAG | BG | DHG | POXC | TKN | pH | SOM | C:N | Leaf N | Leaf C | Leaf C:N | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Silt | ||||||||||||||
| Clay | −0.82 | |||||||||||||
| Sand | −0.94 | 0.58 | ||||||||||||
| NAG | 0.43 | −0.18 | −0.50 | |||||||||||
| BG | 0.43 | −0.24 | −0.47 | 0.41 | ||||||||||
| DHG | 0.39 | −0.17 | −0.46 | −0.09 | 0.31 | |||||||||
| POXC | −0.15 | 0.01 | 0.21 | 0.16 | −0.35 | −0.37 | ||||||||
| TKN | −0.07 | −0.23 | 0.24 | −0.20 | −0.37 | −0.28 | 0.49 | |||||||
| pH | 0.30 | −0.11 | −0.36 | 0.39 | 0.28 | 0.10 | 0.18 | −0.11 | ||||||
| SOM | −0.44 | 0.34 | 0.43 | −0.30 | −0.44 | −0.36 | 0.32 | 0.43 | −0.22 | |||||
| C:N | −0.41 | 0.55 | 0.26 | −0.19 | −0.03 | −0.11 | −0.14 | −0.36 | 0.12 | 0.52 | ||||
| Leaf N | 0.04 | 0.22 | −0.20 | −0.14 | −0.01 | 0.52 | −0.10 | −0.35 | 0.28 | −0.16 | 0.18 | |||
| Leaf C | 0.15 | −0.31 | −0.03 | 0.11 | 0.08 | −0.47 | 0.12 | 0.23 | −0.22 | 0.37 | −0.02 | −0.65 | ||
| Leaf C:N | 0.10 | −0.33 | 0.05 | 0.17 | 0.08 | −0.53 | 0.12 | 0.29 | −0.25 | 0.30 | −0.10 | −0.87 | 0.93 | |
| CO2 | 0.11 | −0.23 | −0.02 | 0.00 | −0.03 | 0.06 | −0.20 | −0.11 | −0.07 | 0.05 | −0.13 | −0.09 | 0.17 | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sofoulaki, E.; Tzanakakis, V.A.; Giannopoulos, G.; Kapellakis, I.; Kabourakis, E.; Chatzistathis, T.; Monokrousos, N. Different Contribution of Olive Groves and Citrus Orchards to Soil Organic Carbon Sequestration: A Field Study in Four Sites in Crete, Greece. Sustainability 2023, 15, 1477. https://doi.org/10.3390/su15021477
Sofoulaki E, Tzanakakis VA, Giannopoulos G, Kapellakis I, Kabourakis E, Chatzistathis T, Monokrousos N. Different Contribution of Olive Groves and Citrus Orchards to Soil Organic Carbon Sequestration: A Field Study in Four Sites in Crete, Greece. Sustainability. 2023; 15(2):1477. https://doi.org/10.3390/su15021477
Chicago/Turabian StyleSofoulaki, Eleni, Vasileios A. Tzanakakis, Georgios Giannopoulos, Iosif Kapellakis, Emmanouil Kabourakis, Theocharis Chatzistathis, and Nikolaos Monokrousos. 2023. "Different Contribution of Olive Groves and Citrus Orchards to Soil Organic Carbon Sequestration: A Field Study in Four Sites in Crete, Greece" Sustainability 15, no. 2: 1477. https://doi.org/10.3390/su15021477
APA StyleSofoulaki, E., Tzanakakis, V. A., Giannopoulos, G., Kapellakis, I., Kabourakis, E., Chatzistathis, T., & Monokrousos, N. (2023). Different Contribution of Olive Groves and Citrus Orchards to Soil Organic Carbon Sequestration: A Field Study in Four Sites in Crete, Greece. Sustainability, 15(2), 1477. https://doi.org/10.3390/su15021477