
The Potential Impact of Changes in Soil and Climate Conditions on Development of the Herb Layer Vegetation of Public Parks in Krakow (Southern Poland)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
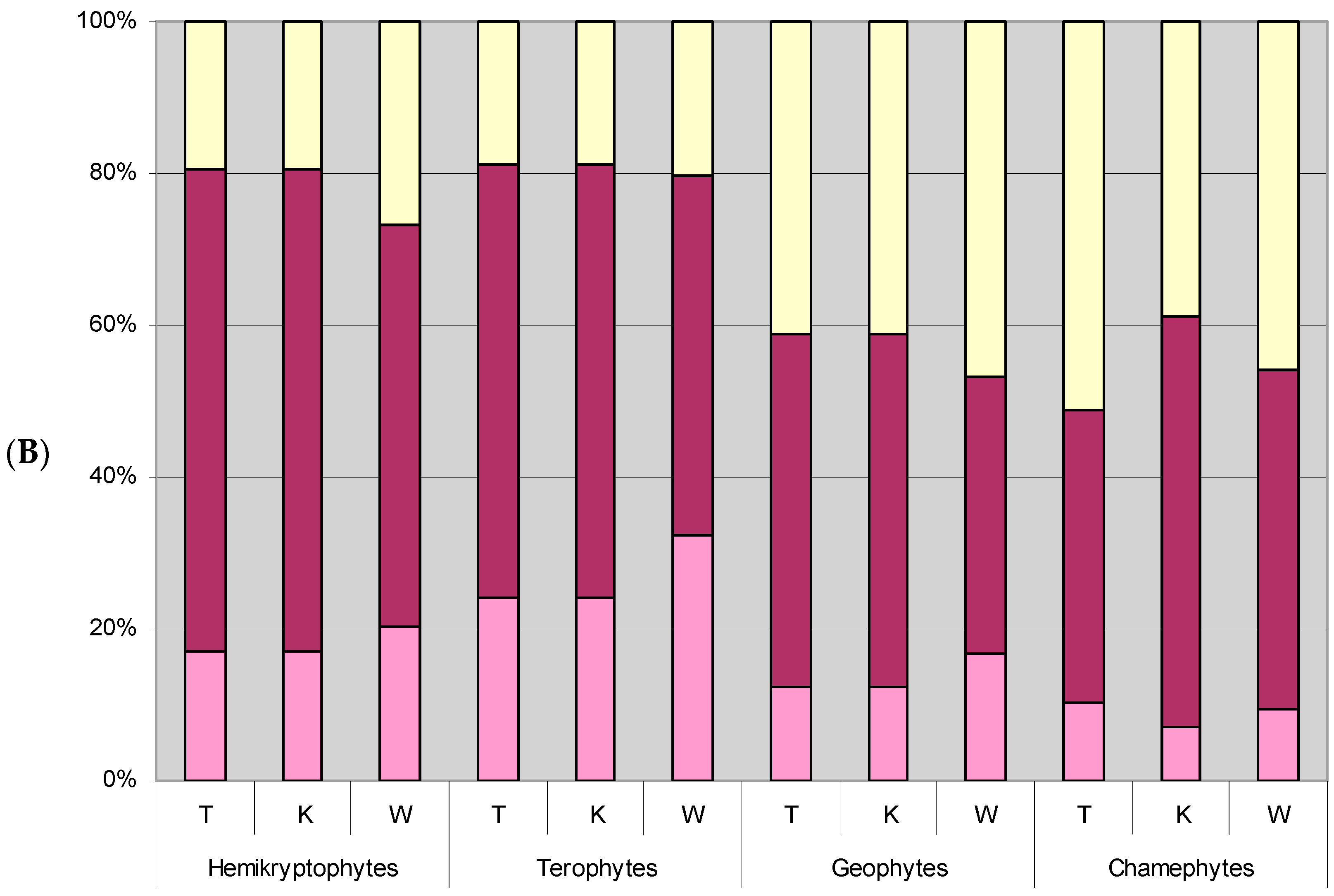{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
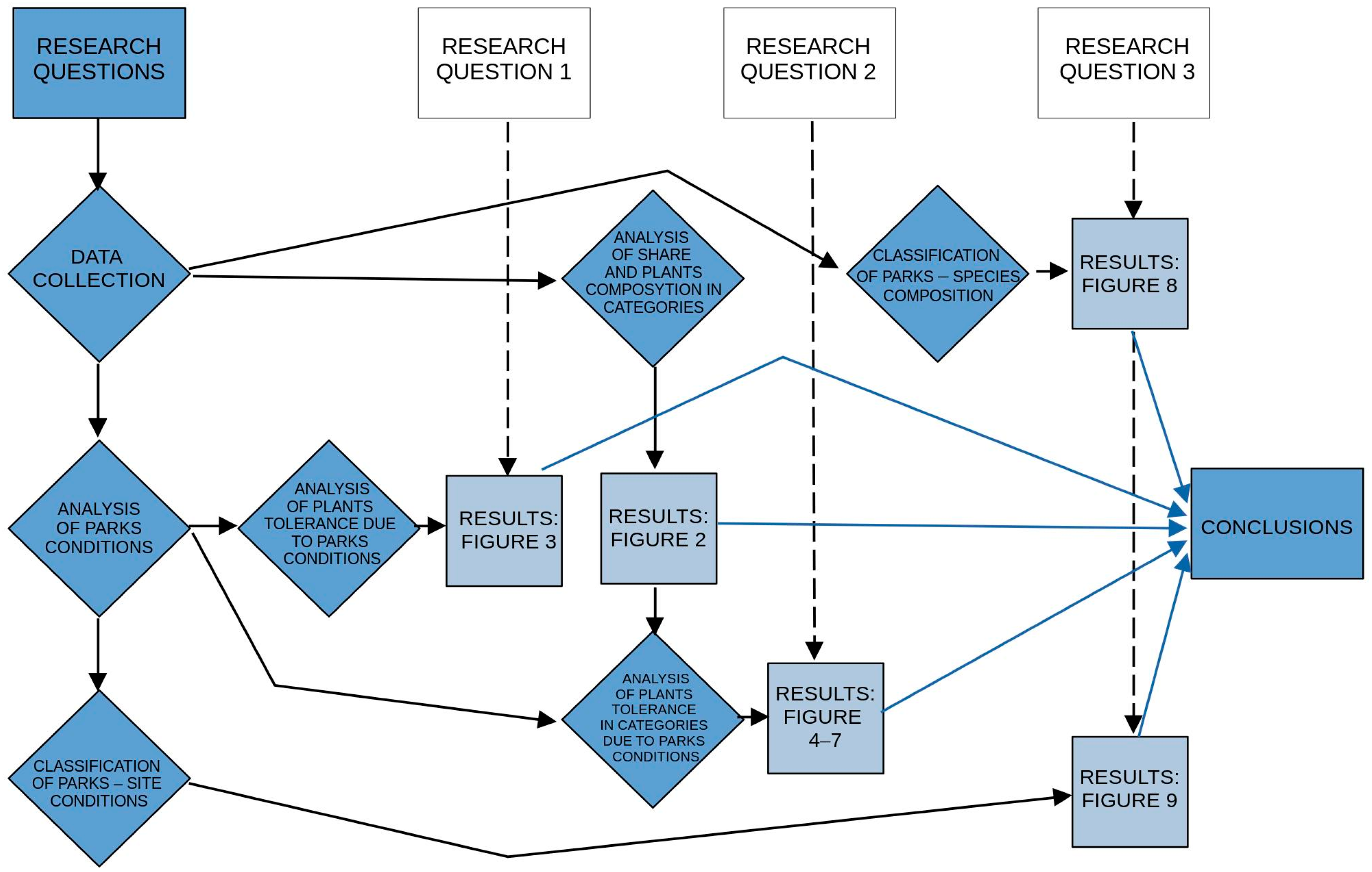
2. Materials and Methods
2.1. Study Area
2.2. Methodology
3. Results
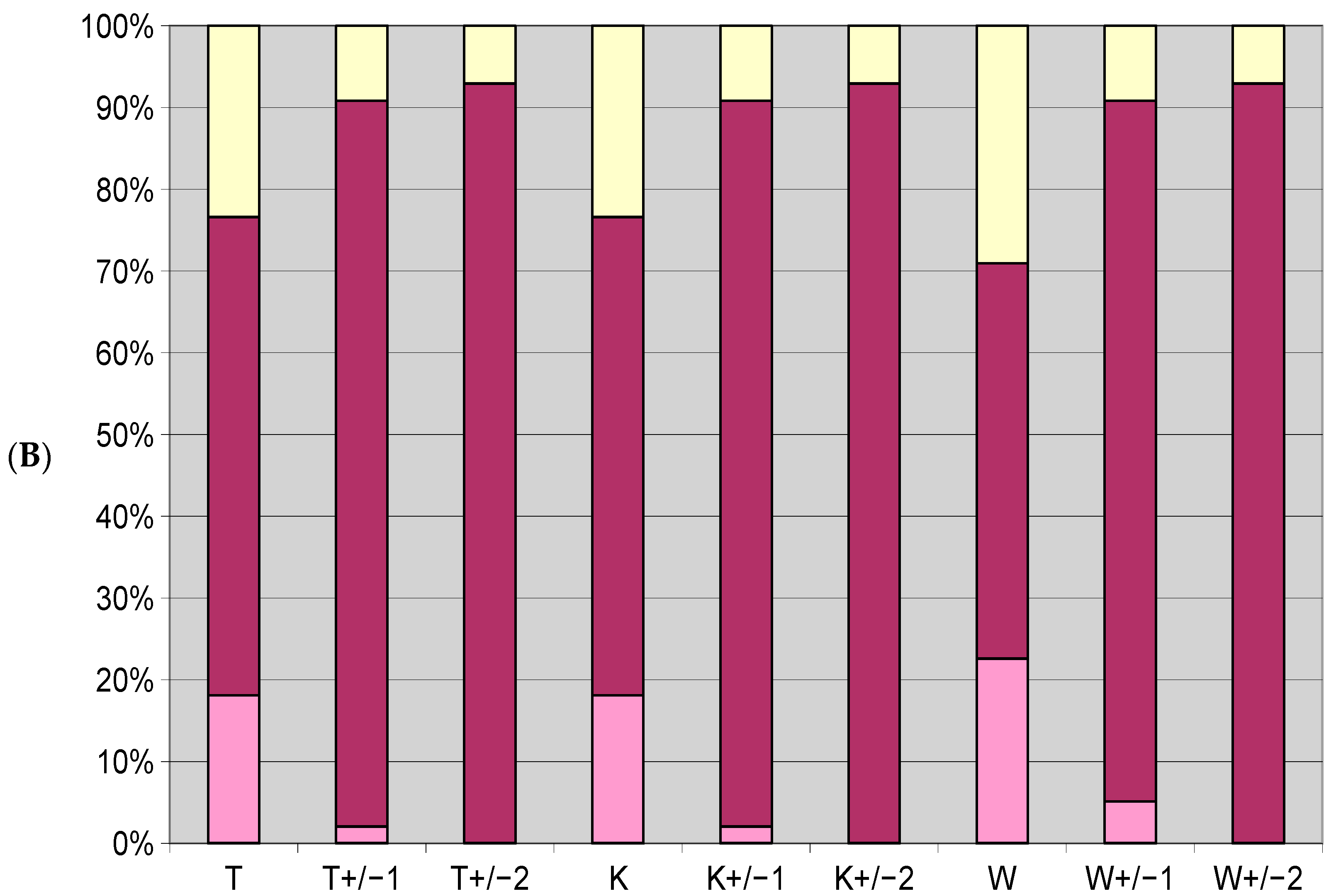
3.1. Differences in Optima and Range of Ecological Tolerance against Park Conditions of All Herb Layer Plants
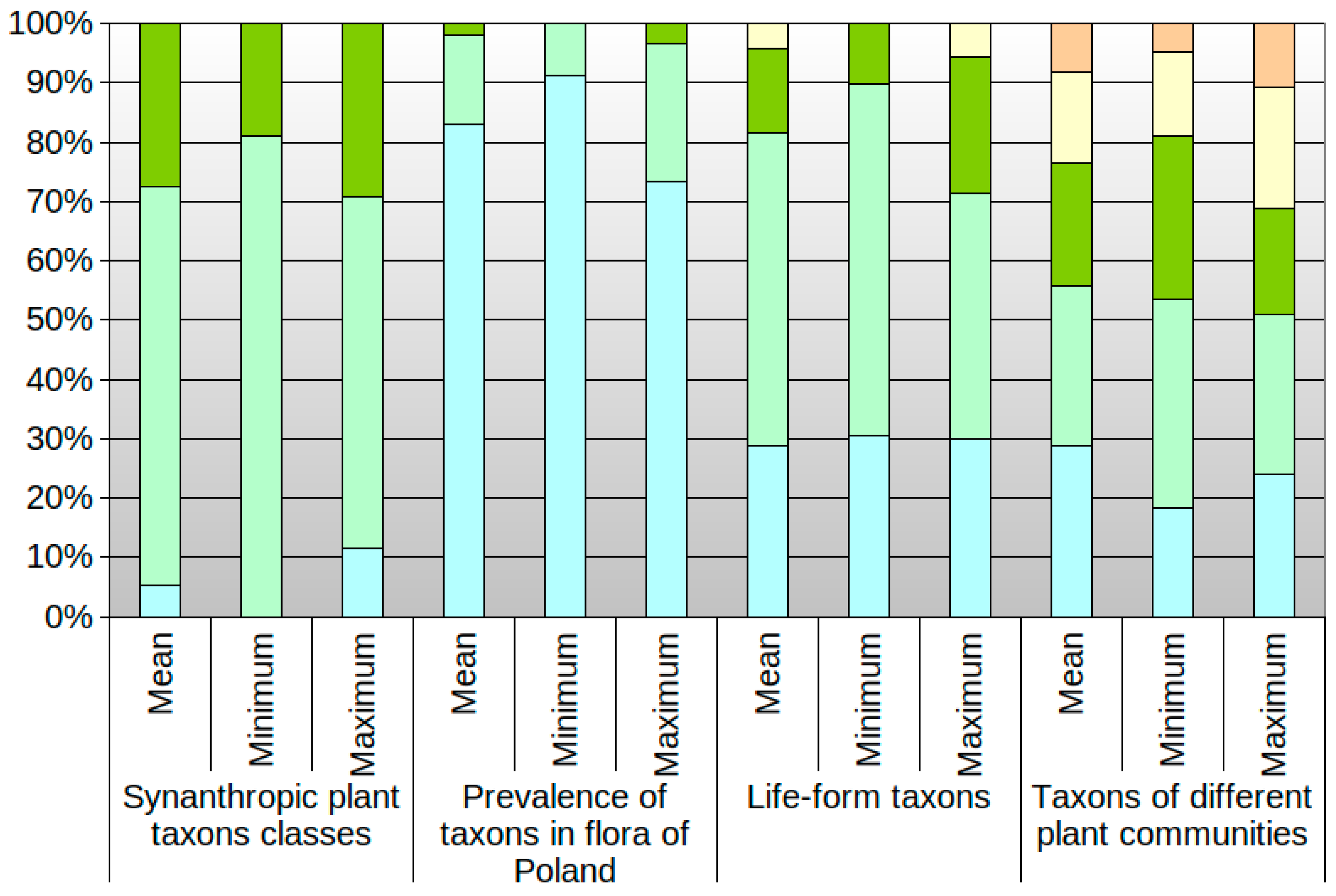
3.2. Differences in Optima and Range of Ecological Tolerance against Park Conditions of Herb Layer Plants in Groups of the Historic-Geographical Category
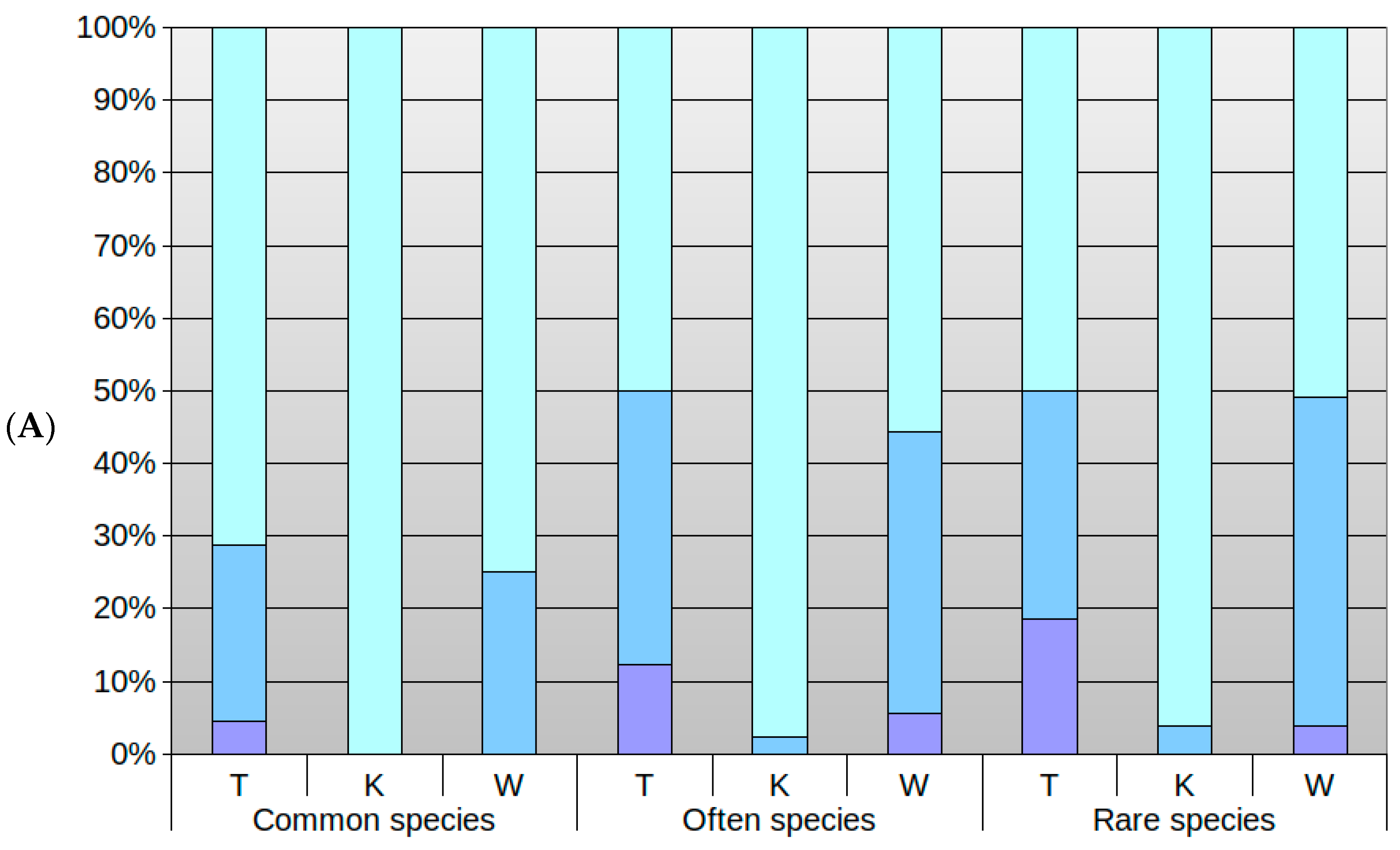
3.3. Differences in Optima and Range of Ecological Tolerance against Park Conditions of Herb Layer Plants in Groups of the Commonness of Taxa in the Flora of Poland
3.4. Differences in Optima and Range of Ecological Tolerance against Park Conditions of Herb Layer Plants in Groups of the Life Form Category
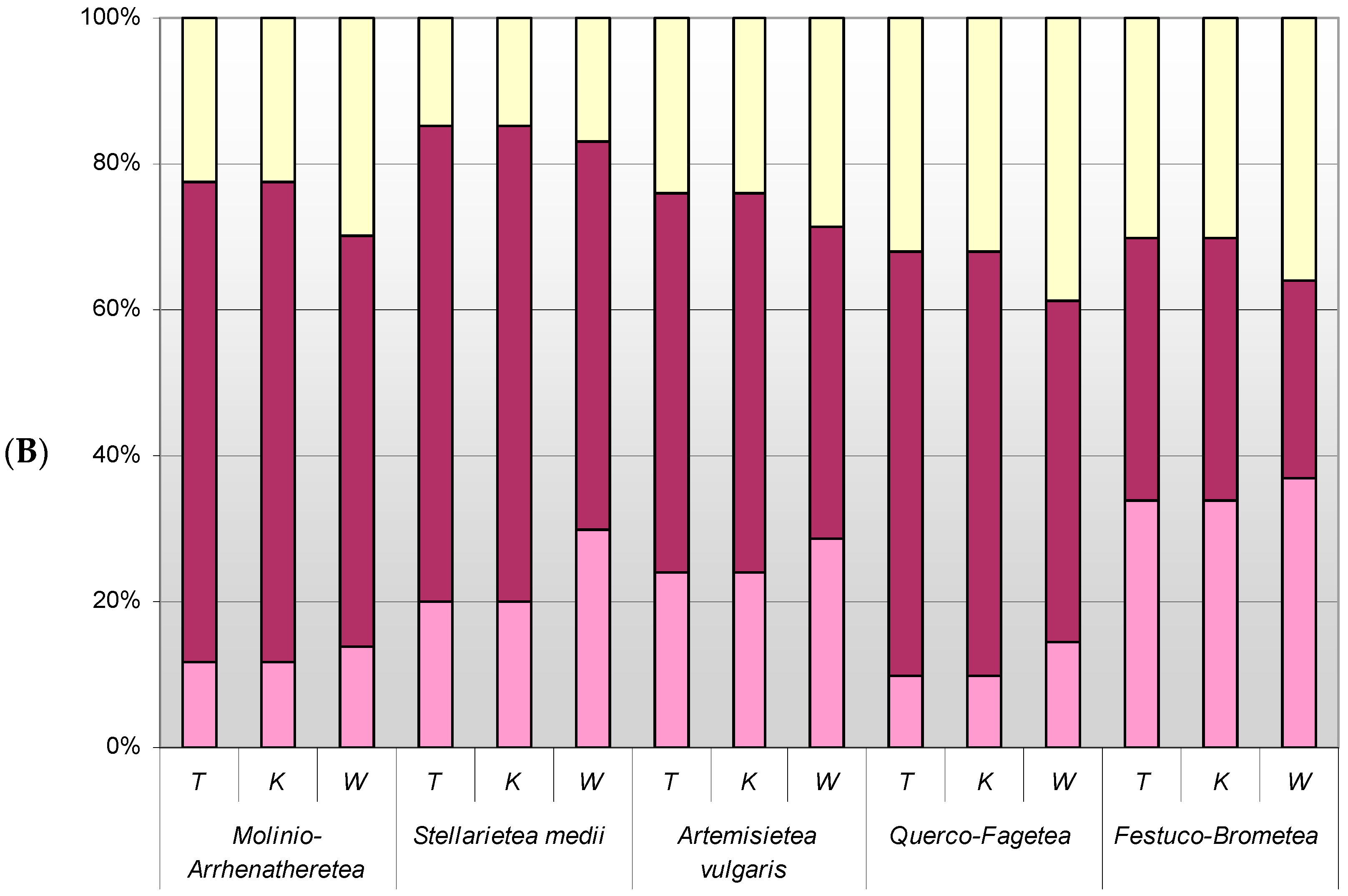
3.5. Differences in Optima and Range of Ecological Tolerance against Park Conditions of Herb Layer Plants in Groups of Different Plant Communities
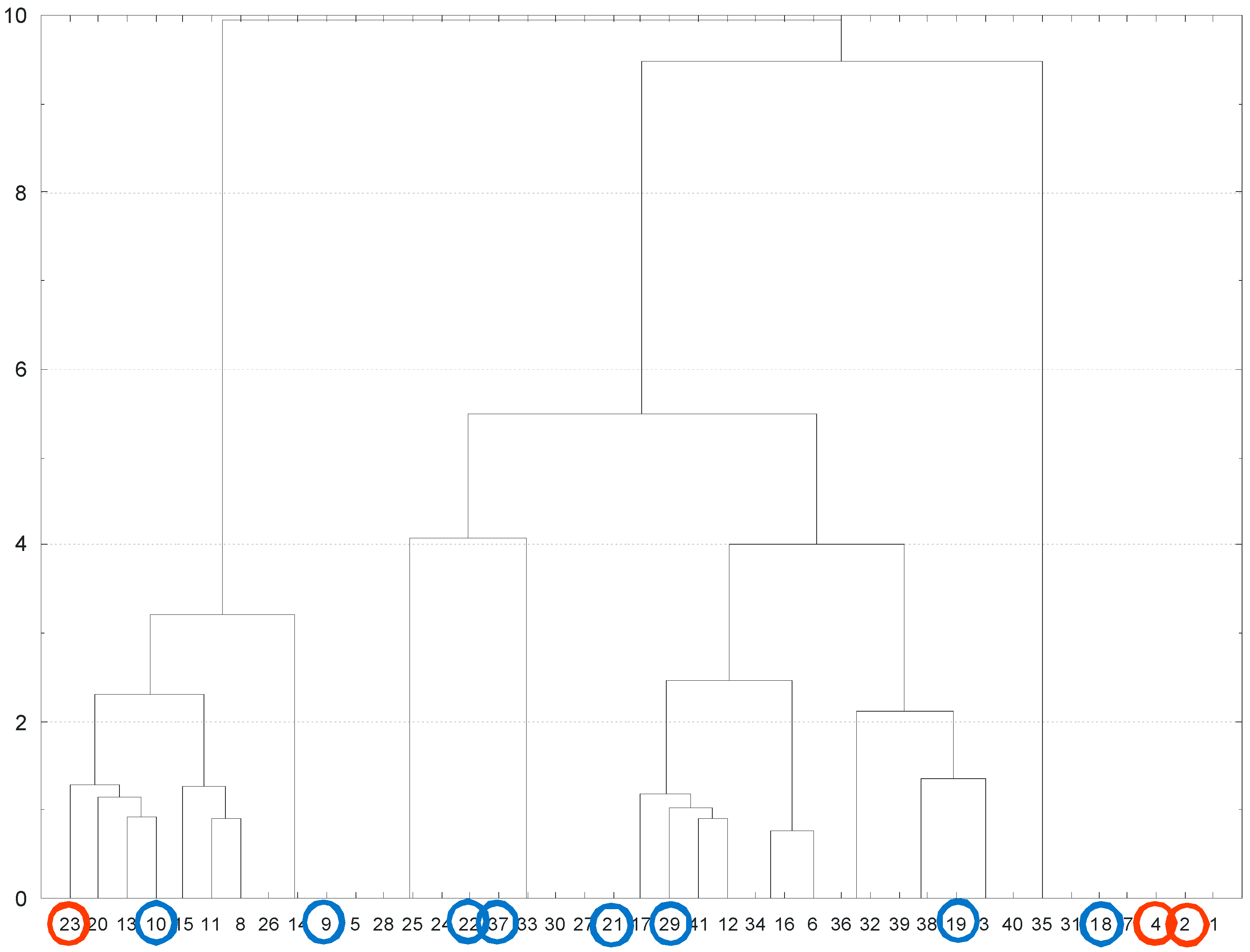
3.6. Relation between Site Conditions and the Plant Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winkler, J.; Matsui, Y.; Filla, J.; Vykydalová, L.; Jiroušek, M.; Vaverková, M.D. Responses of synanthropic vegetation to composting facility. Sci. Total Environ. 2023, 859, 160160. [Google Scholar] [CrossRef] [PubMed]
- Bordim, M.H.S.; Longo, R.M.; Bordim, B.S. Urban environmental sustainability: Analysis of the influence of vegetation in environmental parameters. Rev. De Gestão Ambient. E Sustentabilidade 2022, 11, 1–23. [Google Scholar] [CrossRef]
- Pickett, S.; Cadenasso, M. Linking ecological and built components of urban mosaics: An open cycle of ecological design. J. Ecol. 2008, 96, 8–12. [Google Scholar] [CrossRef]
- Bonthoux, S.; Voisin, L.; Bouché-Pillon, S.; Chollet, S. More than weeds: Spontaneous vegetation in streets as a neglected element of urban biodiversity. Landsc. Urban Plan. 2019, 185, 163–172. [Google Scholar] [CrossRef]
- Florgård, C. Preserved and remnant natural vegetation in cities: A geographically divided field of research. Landsc. Res. 2007, 32, 79–94. [Google Scholar] [CrossRef]
- Reaka-Kudla, M.L.; Wilson, D.E.; Wilson, E.O. (Eds.) Biodiversity II: Understanding and Protecting Our Biological Resources; Joseph Henry Press: Washington, DC, USA, 1996. [Google Scholar]
- Afuye, G.A.; Kalumba, A.M.; Orimoloye, I.R. Characterisation of Vegetation Response to Climate Change: A Review. Sustainability 2021, 13, 7265. [Google Scholar] [CrossRef]
- Marchionni, V.; Fatichi, S.; Tapper, N.; Walker, J.P.; Manoli, G.; Daly, E. Assessing vegetation response to irrigation strategies and soil properties in an urban reserve in southeast Australia. Landsc. Urban Plan. 2021, 215, 104198. [Google Scholar] [CrossRef]
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the message right on nature-based solutions to climate change. Glob. Chang. Biol. 2021, 27, 1518–1546. [Google Scholar] [CrossRef]
- Rafiee, R.; Mahiny, A.S.; Khorasani, N. Assessment of changes in urban green spaces of Mashad city using satellite data. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 431–438. [Google Scholar] [CrossRef]
- Badach, J.; Dymnicka, M.; Baranowski, A. Urban Vegetation in Air Quality Management: A Review and Policy Framework. Sustainability 2020, 12, 1258. [Google Scholar] [CrossRef]
- Uçar, Z.; Akay, A.E.; Bilici, E. Towards green smart cities: Importance of Urban forestry and urban vegetation. International Archives of the Photogrammetry. Remote Sens. Spat. Inf. Sci.-ISPRS Arch. 2020, 2020, 399–403. [Google Scholar] [CrossRef]
- Manning, R.; More, T. Recreational Values of Public Parks. Georg. Wright Forum 2002, 19, 21–30. Available online: http://www.jstor.org/stable/43597798 (accessed on 3 November 2023).
- Staniewska, A. Gardens of Historic Mental Health Hospitals and Their Potential Use for Green Therapy Purposes. Land 2022, 11, 1618. [Google Scholar] [CrossRef]
- Krzeptowska-Moszkowicz, I.; Moszkowicz, Ł.; Porada, K. Evolution of the Concept of Sensory Gardens in the Generally Accessible Space of a Large City: Analysis of Multiple Cases from Kraków (Poland) Using the Therapeutic Space Attribute Rating Method. Sustainability 2021, 13, 5904. [Google Scholar] [CrossRef]
- Krzeptowska-Moszkowicz, I.; Moszkowicz, Ł.; Porada, K. What Affects the Depth of the Human–Garden Relationship in Freely Accessible Urban Sensory Gardens with Therapeutic Features in Various Users? Sustainability 2023, 15, 14420. [Google Scholar] [CrossRef]
- Taylor, H.A. Urban Public Parks, 1840–1900: Design and Meaning. Gard. Hist. 1995, 23, 201–221. [Google Scholar] [CrossRef]
- Low, S.M.; Taplin, D.; Scheld, S. Rethinking Urban Parks: Public Space and Cultural Diversity; University of Texas Press: Austin, TX, USA, 2009. [Google Scholar]
- Gawryluk, D. Historical Public Parks in Podlasie and their Current Day Modernisation. Form. Urban Green Areas 2015, 1, 74–80. [Google Scholar]
- Bobek, W.; Łakomy, K.; Hodor, K. Contemporary processes and the selection of materials in historical urban greenery areas on example from Cracow and Warsaw. IOP Conf. Ser. Mater. Sci. Eng. 2018, 471, 092018. [Google Scholar] [CrossRef]
- Bianco, P.M.; Fanelli, G.; Tescarollo, P.; Pignatti, S. Ruderalization in a Roman Parks as a Result of Changing Managemant. Urban Habitats 2003, 1, 87–104. [Google Scholar]
- LaPaix, R.; Freedman, B. Vegetation Structure and Composition within Urban Parks of Halifax Regional Municipality, Nova Scotia, Canada. Landsc. Urban Plan. 2010, 98, 124–135. [Google Scholar] [CrossRef]
- Concepción, E.D.; Obrist, M.K.; Moretti, M.; Altermatt, F.; Baur, B.; Nobis, M.P. Impacts of urban sprawl on species richness of plants, butterflies, gastropods and birds: Not only built-up area matters. Urban Ecosyst. 2016, 19, 225–242. [Google Scholar] [CrossRef]
- Moszkowicz, Ł.; Krzeptowska-Moszkowicz, I.; Porada, K. Relationship between parameters of public parks and their surroundings and the richness, diversity and species composition of vascular herbaceous plants on the example of Krakow in Central Europe. Landsc. Online 2021, 94, 1–16. [Google Scholar] [CrossRef]
- Venter, O.; Brodeur, N.N.; Nemiroff, L.; Belland, B.; Dolinsek, I.J.; Grant, J.W. Threats to endangered species in Canada. Bioscience 2006, 56, 903–910. [Google Scholar] [CrossRef]
- Woo-Durand, C.; Matte, J.M.; Cuddihy, G.; McGourdji, C.L.; Venter, O.; Grant, J.W. Increasing importance of climate change and other threats to at-risk species in Canada. Environ. Rev. 2020, 28, 449–456. [Google Scholar] [CrossRef]
- Szopińska, E.; Kazak, J.; Kempa, O.; Rubaszek, J. Spatial Form of Greenery in Strategic Environmental Management in the Context of Urban Adaptation to Climate Change. Pol. J. Environ. Stud. 2019, 28, 2845–2856. [Google Scholar] [CrossRef]
- Apostolopoulou, D.; Tsoka, S. Climate change and built environment—The role of urban greenery as a mitigation strategy in Greek urban areas. IOP Conf. Ser. Earth Environ. Sci. 2021, 899, 012018. [Google Scholar] [CrossRef]
- Hedblom, M.; Prevot, A.C.; Grégorie, A. Science fiction blockbuster movies—A problem or a path to urban greenery? Urban For. Urban Green. 2022, 74, 127661. [Google Scholar] [CrossRef]
- MacGillivray, C.W.; Grime, J.P.; The Integrated Screening Programme (ISP) Team. Testing predictions of the resistance and resilience of vegetation subjected to extreme events. Funct. Ecol. 1995, 9, 640–649. [Google Scholar] [CrossRef]
- Steel, Z.L.; Foster, D.; Coppoletta, M.; Lydersen, J.M.; Stephens, S.L.; Paudel, A.; Collins, B.M. Ecological resilience and vegetation transition in the face of two successive large wildfires. J. Ecol. 2021, 109, 3340–3355. [Google Scholar] [CrossRef]
- Czochański, J.; Wiśniewski, P. River valleys as ecological corridors—Structure, function and importance in the conservation of natural resources. Ecol. Quest. 2018, 29, 77–87. [Google Scholar] [CrossRef]
- Szczerek, E. The Problem of Densification of Large-Panel Housing Estates upon the Example of Cracow. Land 2021, 10, 1359. [Google Scholar] [CrossRef]
- Hilgard, E.W. A Report on the Relations of Soil to Climate; Bulletin, U.S. Department of Agriculture, Weather Bureau: Washington, DC, USA, 1892; Volume 3, pp. 1–59.
- Eagleson, P.S. Climate, soil, and vegetation: 6. Dynamics of the annual water balance. Water Resour. Res. 1978, 14, 749–764. [Google Scholar] [CrossRef]
- Sevink, J. Soil development in the coastal dunes and its relation to climate. Landscape Ecol. 1991, 6, 49–56. [Google Scholar] [CrossRef]
- Sanchis, M.P.S.; Torri, D.; Borselli, L.; Poesen, J. Climate effects on soil erodibility. Earth Surf. Process. Landf. 2008, 33, 1082–1097. [Google Scholar] [CrossRef]
- Huntley, B. How Plants Respond to Climate Change: Migration Rates, Individualism and the Consequences for Plant Communities. Ann. Bot. 1991, 67, 15–22. Available online: http://www.jstor.org/stable/42758387 (accessed on 17 November 2023). [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Wilson, E.O.; MacArthur, R.H. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; Volume 1. [Google Scholar]
- Honnay, O.; Endels, P.; Vereecken, H.; Hermy, M. The role of patch area and habitat diversity in explaining native plant species richness in disturbed suburban forest patches in northern Belgium. Divers. Distrib. 1999, 5, 129–141. [Google Scholar] [CrossRef]
- Couvreur, M.; Christiaen, B.; Verheyen, K.; Hermy, M. Large herbivores as mobile links between isolated nature reserves through adhesive seed dispersal. Appl. Veg. Sci. 2004, 7, 229–236. [Google Scholar] [CrossRef]
- Kowarik, I. Some responses of flora and vegetation to urbanization in Central Europe. In Urban Ecology: Plants and Plant Communities in Urban Environments; Sukopp, H., Hejny, S., Kowarik, I., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1990; pp. 45–74. [Google Scholar]
- Ranta, P.; Viljanen, V. Vascular plants along an urban-rural gradient in the city of Tampere, Finland. Urban Ecosyst. 2011, 14, 361–376. [Google Scholar] [CrossRef]
- Czortek, P.; Pielech, R. Surrounding landscape influences functional diversity of plant species in urban parks. Urban For. Urban Green. 2020, 47, 126525. [Google Scholar] [CrossRef]
- Cain, M.C.; Damman, H.; Muir, A. Seed dispersal and the Holocene migration of woodland herbs. Ecol. Monogr. 1998, 68, 325–347. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2021, 41, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Waring, G.L.; Cobb, N.S. The impact of plant stress on herbivore population dynamics. Insect-Plant Interact. 1992, 4, 167–226. [Google Scholar]
- Ahmad, P.; Prasad, M.N.V. (Eds.) Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Springer Science & Business Media: New York, NY, USA, 2011. [Google Scholar]
- Grime, J.P. Plant Strategies and Vegetation Processes Vegetation Processes; John Wiley & Sons, Limited: Chichester, UK, 1979. [Google Scholar]
- Sandvik, S.M.; Totland, Ø. Short-term effects of simulated environmental changes on phenology, reproduction, and growth in the late-flowering snowbed herb Saxifraga stellaris L. Ecoscience 2000, 7, 201–213. [Google Scholar] [CrossRef]
- Shuman, B.; Newby, P.; Huang, Y.; Webb, T. Evidence for the close climatic control of New England vegetation history. Ecology 2004, 85, 1297–1310. [Google Scholar] [CrossRef]
- Van Der Veken, S.; Bossuyt, B.; Hermy, M. Climate gradients explain changes in plant community composition of the forest understory: An extrapolation after climate warming. Belg. J. Bot. 2004, 137, 55–69. [Google Scholar]
- Moszkowicz, Ł.; Krzeptowska-Moszkowicz, I. Impact of the public parks location in the city on the richness and diversity of herbaceous vascular plants on the example of Krakow Southern Poland. Plants Urban Areas Landsc. 2020, 98–103. [Google Scholar] [CrossRef]
- Hamrick, J.L. Response of forest trees to global environmental changes. For. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Breshears, D.; Huxman, T.; Adams, H.; Zou, C.; Davison, J. Vegetation synchronously leans upslope as climate warms. Proc. Natl. Acad. Sci. USA 2008, 105, 11591–11592. [Google Scholar] [CrossRef]
- Kapfer, J.; Hédl, R.; Jurasinski, G.; Kopecký, M.; Schei, F.H.; Grytnes, J.A. Resurveying historical vegetation data—Opportunities and challenges. Appl. Veg. Sci. 2017, 20, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Williams, S.E.; Shoo, L.P.; Isaac, J.L.; Hoffmann, A.A.; Langham, G. Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biol. 2008, 6, e3252008. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.; Agudo, R. The Future of Species Under Climate Change: Resilience or Decline? Science 2013, 341, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Kondracki, J. Geografia Regionalna Polski. Wydanie 2 Poprawione; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2000. [Google Scholar]
- Kornaś, J.; Medwecka-Kornaś, A. Szata roślinna Krakowa. Folia Geogr. Ser. Geogr.-Phys. 1974, 8, 153–169. [Google Scholar]
- Wójcik, A. Budowa geologiczna.—W. In Atlas Kampusu 600-Lecia Odnowienia Uniwersytetu Jagiellońskiego; Jędrychowski, I., Ed.; Instytut Geografi i i Gospodarki Przestrzennej UJ: Kraków, Poland, 2007. [Google Scholar]
- Trzcińska-Tacik, H. Flora synantropijna Krakowa. Zesz. Nauk. UJ Kraków Rozpr. 1979, 32, 1–249. [Google Scholar]
- Dubiel, E. Mapa roślinności rzeczywistej miasta Krakowa [Map of actual vegetation of the city of Cracow]. Zesz. Nauk. UJ Prace Bot. 1991, 22, 121–133. [Google Scholar]
- Dubiel, E.; Szwagrzyk, J. (Eds.) Atlas Roślinności Rzeczywistej Krakowa; Urząd Miasta Krakowa, Wydział Kształtowania Środowiska: Kraków, Poland, 2008. [Google Scholar]
- Berdau, F. Flora Cracoviensis; Typis, C. R. Universitatis Jagiellonicae: Cracoviae, Poland, 1859; s. viii + 448. [Google Scholar]
- Raciborski, M. Zmiany zaszłe we florze okolic Krakowa w ciągu ostatnich lat dwudziestu pięciu pod względem roślin dziko rosnących. Spraw. Komis. Fizjogr. 1884, 18, 99–126. [Google Scholar]
- Hess, M. Klimat Krakowa. Folia Geographica. Ser. Geogr.-Phys. 1974, 8, 45–102. [Google Scholar]
- Matuszko, D. (Ed.) Klimat Krakowa w XX Wieku; Instytut Geografii i Gospodarki Przestrzennej Uniwersytetu Jagiellońskiego: Kraków, Poland, 2007; pp. 1–251. [Google Scholar]
- Bokwa, A. Wieloletnie Zmiany Struktury Mezoklimatu Miasta na Przykładzie Krakowa; Instytut Geografii i Gospodarki Przestrzennej Uniwersytetu Jagiellońskiego: Kraków, Poland, 2010; pp. 1–258. [Google Scholar]
- Piotrowicz, K. Sezonowa i Wieloletnia Zmienność Typów Pogody w Krakowie; Instytut Geografii i Gospodarki Przestrzennej Uniwersytetu Jagiellońskiego: Kraków, Poland, 2010; pp. 1–314. [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefeaßpflanzen Mitteleuropas; Scripta Geobotanica: Göttingen, Germany, 1974; Volume 9, pp. 1–97. [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefeaßpflanzen Mitteleuropas. 2; Auflage. Scripta Geobotanica: Göttingen, Germany, 1979; Volume 9, pp. 1–122. [Google Scholar]
- Frank, D.; Klotz, S. Biologisch-Ökologische Daten zur Flora der DDR (Biological-Ecological Data on the Flora of the GDR); Wissenschaftliche Beiträge der Martin-Luther-Universität Halle-Wittenberg: Halle, Germany, 1988; pp. 1–168. [Google Scholar]
- Zarzycki, K.; Trzcinska-Tacik, H.; Rózanski, W.; Szalag, Z.; Wolek, J.; Korzeniak, U. Ecological Indicator Values of Vascular Plants of Vascular Plants of Poland; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002. [Google Scholar]
- Persson, S. Ecological Indicator Values as an Aid in the Interpretation of Ordination Diagrams. J. Ecol. 1981, 69, 71–84. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Gremmen, N.J.M. Ecological amplitudes of plant species and the internal consistency of Ellenberg’s indicator values for moisture. Vegetatio 1987, 69, 79–87. [Google Scholar] [CrossRef]
- Berg, C.; Welk, E.; Jaeger, E. Revising Ellenberg’s indicator values for continentality based on global vascular plant species distribution. Appl. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Roo-Zielińska, E. Porównanie europejskich skal ekologicznych liczb wskaźnikowych w ocenie środowiska fizycznogeograficznego na podstawie charakterystycznych gatunków roślin wrzosowisk i ubogich muraw bliźniczkowych z klasy Nardo-Callunetea= A comparison of European scales of ecological indicator values in assessing the natural environment on the basis of species characteristic for heaths and poor grasslands of class Nardo-Callunetea. Przegląd Geogr. 2018, 90, 403–434. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Scripta Geobotanica: Göttingen, Germany, 1991; Volume 18, pp. 1–248. [Google Scholar]
- Kornaś, J. Geograficzno-historyczna klasyfikacja roślin synantropijnych. Mat. Zakł. Fitosoc. Stos. UW 1968, 25, 33–41. [Google Scholar]
- Chmiel, J. Zróżnicowanie Przestrzenne Flory Jako Podstawa Ochrony Przyrody w Krajobrazie Rolniczym; Bogucki Wydawnictwo Naukowe: Poznañ, Poland, 2006; pp. 1–250. [Google Scholar]
- Zając, A.; Zając, M. A tentative list of segetal and ruderal apophytes in Poland.—Prowizoryczna lista apofitów segetalnych i ruderalnych w Polsce. Zesz. Nauk. Uniw. Jagiellońskiego. Pr. Bot. 1992, 24, 7–23. [Google Scholar]
- Zając, A.; Zając, M. Atlas Rozmieszczenia Roślin Naczyniowych w Polsce: Dodatek/Distribution Atlas of Vascular Plants in Poland: Appendix; Instytut Botaniki Uniwersytetu Jagiellońskiego: Kraków, Poland, 2019. [Google Scholar]
- Zarzycki, K.; Mirek, Z. Red list of plants and fungi in Poland. In Czerwona Lista Roślin i Grzybów Polski; Wojewoda, W., Szeląg, Z., Eds.; Instytut Botaniki im. W. Szafera PAN: Kraków, Poland, 2006. [Google Scholar]
- Raunkiær, C. Planterigets Livsformer og deres Betydning for Geografien; Gyldendalske Boghandel-Nordisk Forlag: København and Kristiania, Denmark, 1907; pp. 1–132. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; PWN: Warszawa, Poland, 2004. [Google Scholar]
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Haeuser, E.; Dawson, W.; van Kleunen, M. The effects of climate warming and disturbance on the colonization potential of ornamental alien plant species. J. Ecol. 2017, 105, 1698–1708. [Google Scholar] [CrossRef]
- Turbelin, A.; Catford, J.A. Invasive plants and climate change. In Climate Change; Elsevier: Amsterdam, The Netherlands, 2021; pp. 515–539. [Google Scholar] [CrossRef]
- Chelli, S.; Wellstein, C.; Campetella, G.; Canullo, R.; Tonin, R.; Zerbe, S.; Gerdol, R. Climate change response of vegetation across climatic zones in Italy. Clim. Res. 2017, 71, 249–262. [Google Scholar] [CrossRef]
- Brusca, R.C.; Wiens, J.F.; Meyer, W.M.; Eble, J.; Franklin, K.; Overpeck, J.T.; Moore, W. Dramatic response to climate change in the Southwest: Robert Whittaker’s 1963 Arizona Mountain plant transect revisited. Ecol. Evol. 2013, 3, 3307–3319. [Google Scholar] [CrossRef]
- Sætersdal, M.; Birks, H.J.B. A comparative ecological study of Norwegian mountain plants in relation to possible future climatic change. J. Biogeogr. 1997, 24, 127–152. [Google Scholar] [CrossRef]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will Climate Change Promote Alien Plant Invasions? In Biological Invasions; Nentwig, W., Ed.; Ecological Studies 193; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar] [CrossRef]
- Liu, Y.; Oduor, A.M.O.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.; van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Matlack, G.R. Plant species migration in mixed-history forest landscape in eastern North America. Ecology 1994, 75, 1491–1502. [Google Scholar] [CrossRef]
- Brunet, J.; Oheimb, G. Migration of vascular plants to secondary woodlands in souther Sweden. J. Ecol. 1998, 86, 429–438. [Google Scholar] [CrossRef]
- Orczewska, A.; Ferens, M. Migration of herb layer species into the poorest post-agricultural pine woods adjacent to ancient Pine forests. Pol. J. Ecol. 2011, 59, 75–85. [Google Scholar]
- Moszkowicz, L. Distribution of vascular plant species in woodland patches of Ojców National Park (southern Poland) in relation to seed dispersal. Acta Soc. Bot. Pol. 2016, 85, 3484. [Google Scholar] [CrossRef]
- Kostrakiewicz-Gierałt, K.; Gmyrek, K.; Pliszko, A. The Effect of the Distance from a Path on Abiotic Conditions and Vascular Plant Species in the Undergrowth of Urban Forests and Parks. Int. J. Environ. Res. Public Health 2022, 19, 5621. [Google Scholar] [CrossRef]
- Krzeptowska-Moszkowicz, I.; Moszkowicz, Ł. Selected problems of Ailanthus altissima (Mill.) Swingle presence in urban spaces: The case of the city centre of Kraków. In Proceedings of the Plants Urban Areas Landscape International Scientific Conference, Nitra, Slovakia, 14–15 May 2014. [Google Scholar] [CrossRef]
- Lamsal, P.; Kumar, L.; Aryal, A.; Atreya, K. Invasive alien plant species dynamics in the Himalayan region under climate change. Ambio 2018, 47, 697–710. [Google Scholar] [CrossRef]
- Peterken, G.F. Natural Woodland. Ecology and Conservation in Northern Temperate Regions; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Kim, J.D.; Park, G.E.; Lim, J.-H.; Yun, C.W. Vegetation Type Classification and Endemic-Rare Plants Investigation in Forest Vegetation Area Distributed by Vulnerable Species to Climate Change, Mt. Jiri. J. Korean Soc. For. Sci. 2018, 107, 113–125. [Google Scholar] [CrossRef]
- Austin, M.P. Continuum Concept, Ordination Methods, and Niche Theory. Annu. Rev. Ecol. Syst. 1985, 16, 39–61. Available online: http://www.jstor.org/stable/2097042 (accessed on 9 December 2021). [CrossRef]
- Silvertown, J. Plant coexistence and the niche. Trends Ecol. Evol. 2004, 19, 605–611. [Google Scholar] [CrossRef]
- Anacker, B.L.; Gogol-Prokurat, M.; Leidholm, K.; Schoenig, S. Climate change vulnerability assessment of rare plants in California. Madroño 2013, 60, 193–210. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Shaver, G.R.; Giblin, A.E.; Nadelhoffer, K.J.; Laundre, J.A. Responses of arctic tundra to experimental and observed changes in climate. Ecology 1995, 76, 694–711. [Google Scholar] [CrossRef]
- Schmiedel, U.; Dengler, J.; Etzold, S. Vegetation dynamics of endemic-rich quartz fields in the Succulent Karoo, South Africa, in response to recent climatic trends. J. Veg. Sci. 2012, 23, 292–303. [Google Scholar] [CrossRef]
- Halevy, A.H. Recent advances in control of flowering and growth habit of geophytes. Acta Hortic. 1990, 266, 35–42. [Google Scholar] [CrossRef]
- Eppich, B.; Dede, L.; Ferenczy, A.; Ágnes, G.; Horváth, L.; Isépy, I.; Priszter, S.; Hufnagel, L. Climatic effects on the phenology of geophytes. Appl. Ecol. Environ. Res. 2009, 7, 253–266. [Google Scholar] [CrossRef]
- Borochov, A.; Spiegelstein, H.; Weiss, D. Dormancy and storage of geophytes. Acta Hortic. 1997, 430, 405–410. [Google Scholar] [CrossRef]
- Laiviņš, M. Environmental changes related dynamics of the number of sites of rare indigenous. Balt. For. 1990, 3, 9–18. [Google Scholar]
- Hill, M.O.; Roy, D.B.; Thompson, K. Hemeroby, urbanity and ruderality: Bioindicators of disturbance and human impact. J. Appl. Ecol. 2002, 39, 708–720. [Google Scholar] [CrossRef]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moszkowicz, Ł.; Krzeptowska-Moszkowicz, I.; Porada, K.; Zieliński, M. The Potential Impact of Changes in Soil and Climate Conditions on Development of the Herb Layer Vegetation of Public Parks in Krakow (Southern Poland). Sustainability 2024, 16, 451. https://doi.org/10.3390/su16010451
Moszkowicz Ł, Krzeptowska-Moszkowicz I, Porada K, Zieliński M. The Potential Impact of Changes in Soil and Climate Conditions on Development of the Herb Layer Vegetation of Public Parks in Krakow (Southern Poland). Sustainability. 2024; 16(1):451. https://doi.org/10.3390/su16010451
Chicago/Turabian StyleMoszkowicz, Łukasz, Izabela Krzeptowska-Moszkowicz, Karolina Porada, and Miłosz Zieliński. 2024. "The Potential Impact of Changes in Soil and Climate Conditions on Development of the Herb Layer Vegetation of Public Parks in Krakow (Southern Poland)" Sustainability 16, no. 1: 451. https://doi.org/10.3390/su16010451
APA StyleMoszkowicz, Ł., Krzeptowska-Moszkowicz, I., Porada, K., & Zieliński, M. (2024). The Potential Impact of Changes in Soil and Climate Conditions on Development of the Herb Layer Vegetation of Public Parks in Krakow (Southern Poland). Sustainability, 16(1), 451. https://doi.org/10.3390/su16010451