
Fungal Pretreatment of Willow Sawdust with Abortiporus biennis for Anaerobic Digestion: Impact of an External Nitrogen Source



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. WSD Used
2.2. Pretreatment Conditions
2.2.1. Fungal Strains
2.2.2. Fungal Pretreatment: The Effect of External Nitrogen Source
2.2.3. Combination of Fungal with Alkali Pre-treatment
2.3. BMP Tests
2.4. Analytical Methods and Procedures
3. Results and Discussion
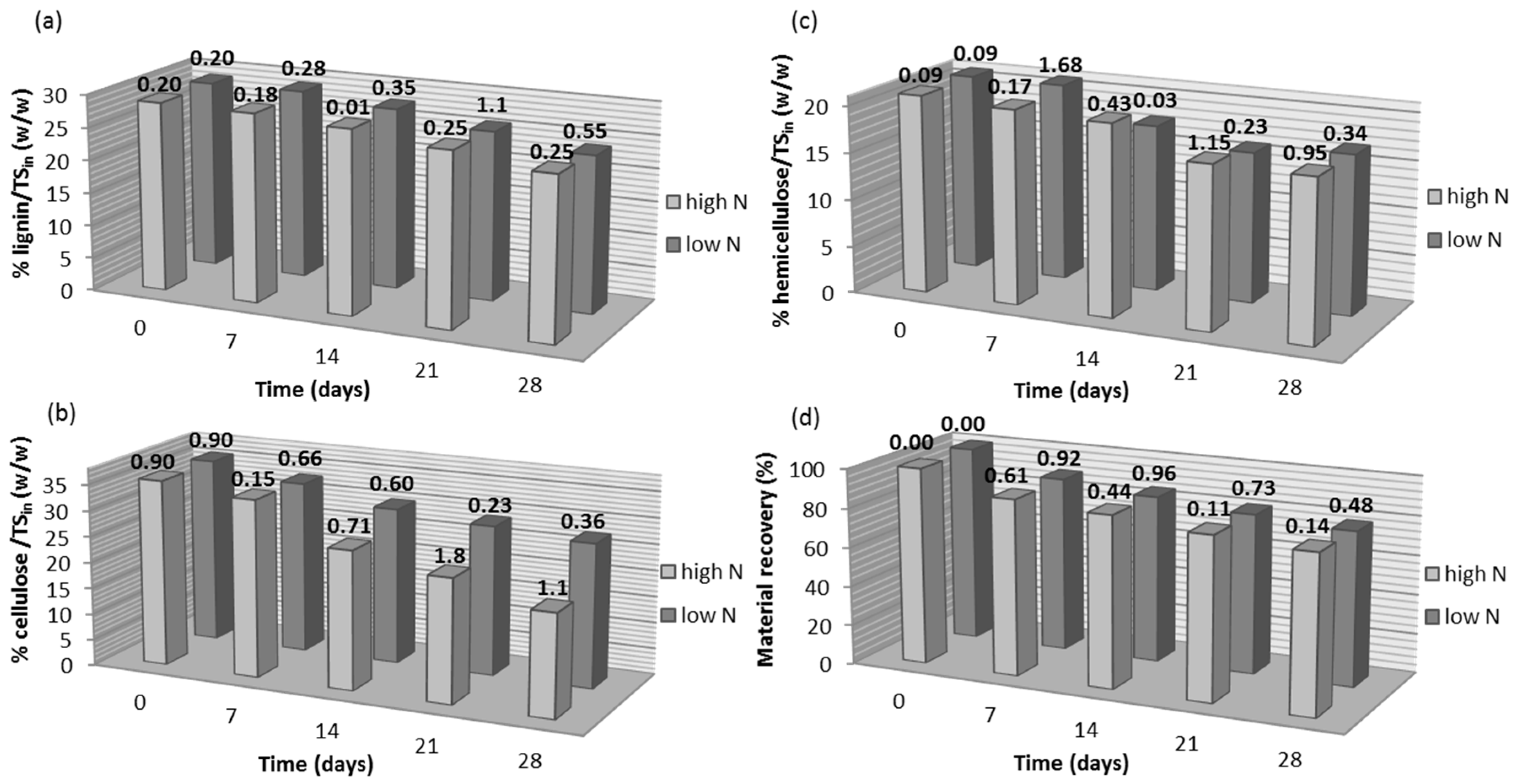
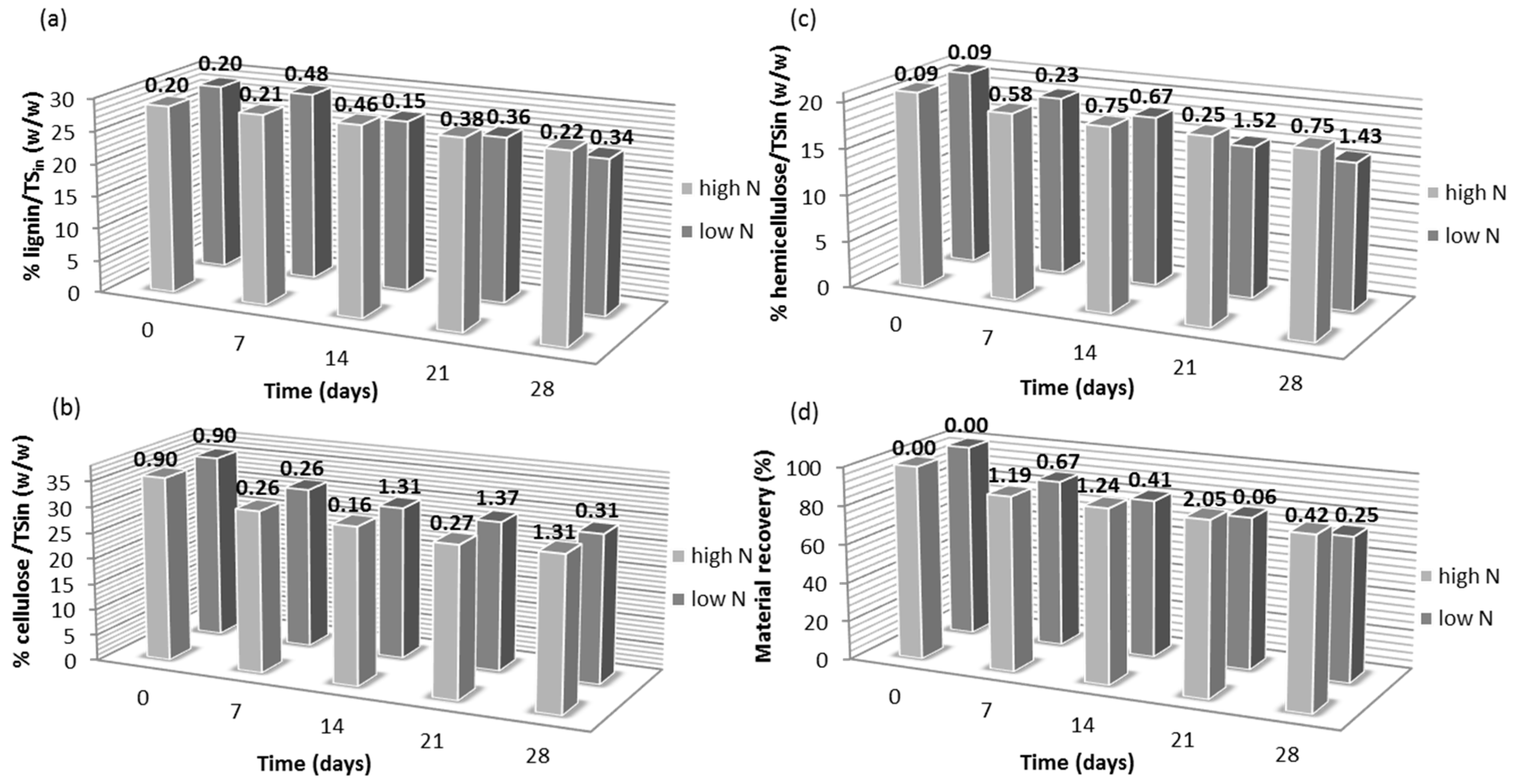
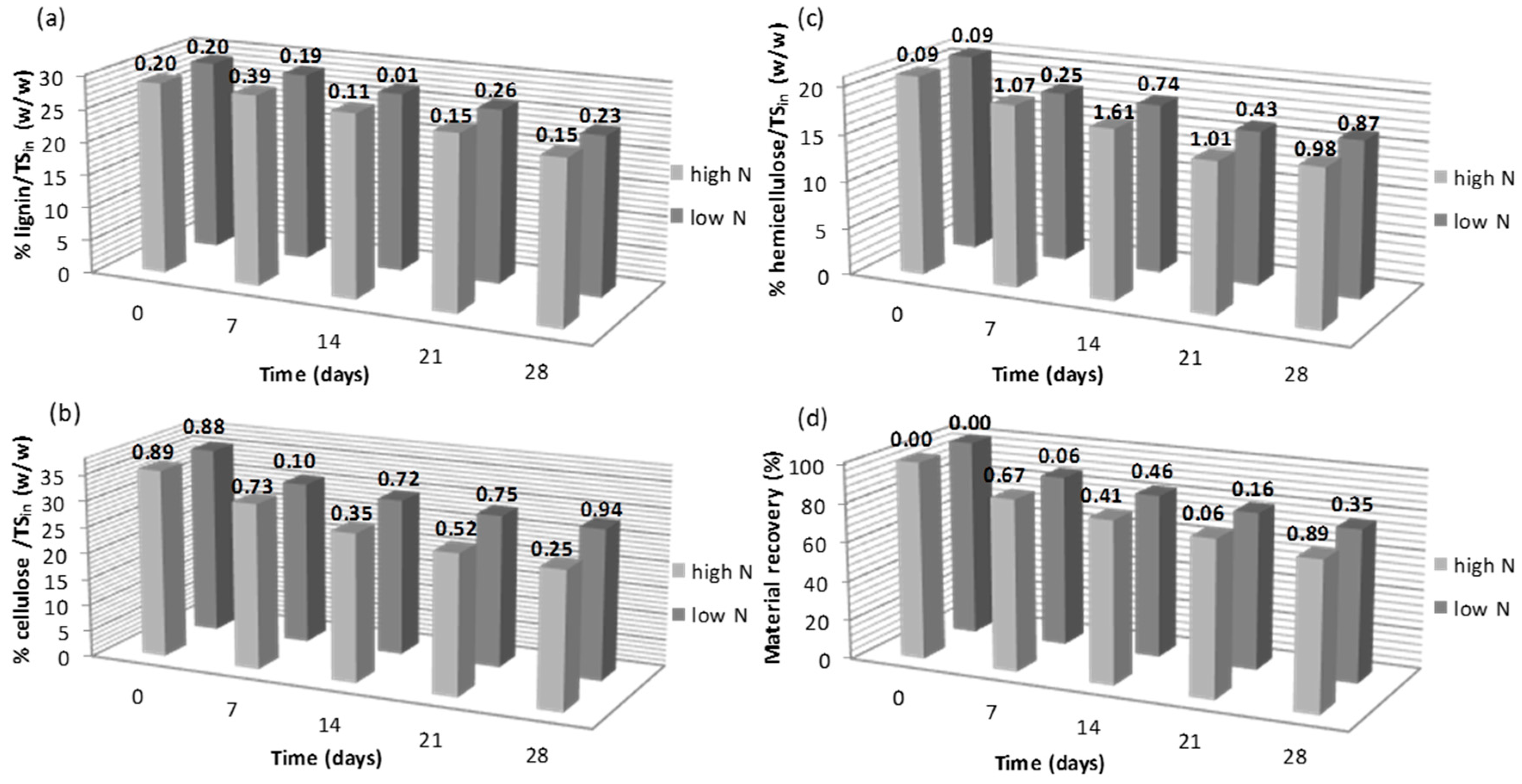
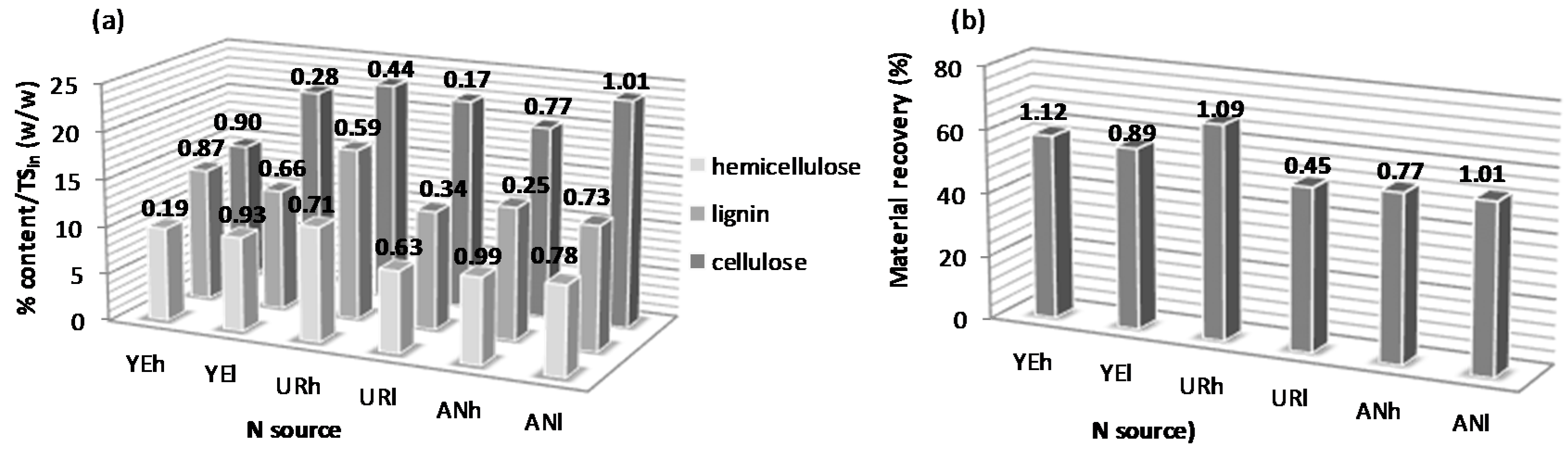
3.1. Effect of Nitrogen Supplementation on the Lignocellulosic Content during Fungal Pretreatment
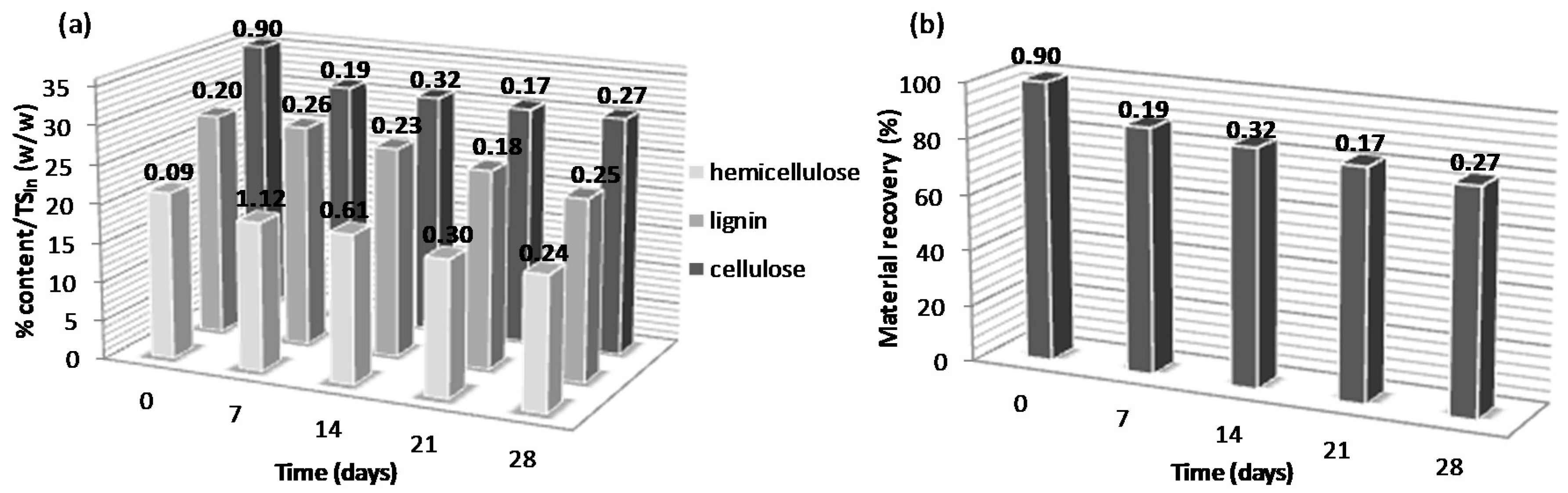
3.2. Effect of Alkaline Pretreatment on the Lignocellulosic Content of Biologically Pretreated WSD
3.3. Methanogenic Potential of Fungi and Combined Fungi/Alkali Pretreated WSD
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Börjesson, P.; Mattiasson, B. Biogas as a resource-efficient vehicle fuel. Trends Biotechnol. 2008, 26, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Rouches, E.; Herpoël-Gimbert, I.; Steyer, J.P.; Carrere, H. Improvement of anaerobic degradation by white-rot fungi pretreatment of lignocellulosic biomass: A review. Renew. Sustain. Energy Rev. 2016, 59, 179–198. [Google Scholar] [CrossRef]
- Carrere, H.; Antonopoulou, G.; Passos, F.; Affes, R.; Battimelli, A.; Lyberatos, G.; Ferrer, I. Review of pretreatment strategies for the most common anaerobic digestion feedstocks: From lab-scale research to full-scale application. Bioresour. Technol. 2016, 199, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, G.; Dimitrellos, G.; Beobide, A.S.; Vayenas, D.; Lyberatos, G. Chemical pretreatment of sunflower straw biomass: The effect on chemical composition and structural changes. Waste Biomass Valoriz. 2015, 6, 733–746. [Google Scholar] [CrossRef]
- Antonopoulou, G.; Dimitrellos, G.; Beobide, A.S.; Vayenas, D.; Lyberatos, G. Ethanol and hydrogen production from sunflower straw: The effect of pretreatment on the whole slurry fermentation. Biochem. Eng. J. 2016, 116, 65–74. [Google Scholar] [CrossRef]
- Alexandropoulou, M.; Antonopoulou, G.; Fragkou, E.; Ntaikou, I.; Lyberatos, G. Fungal pretreatment of willow sawdust and its combination with alkaline treatment for enhancing biogas production. J. Environ. Manag. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tuor, U.; Winterhalter, K.; Fiechter, A. Enzymes of white-rot fungi involved in lignin degradation and ecological determinants for wood decay. J. Biotechnol. 1995, 41, 1–17. [Google Scholar] [CrossRef]
- Dashtban, M.; Schraft, H.; Syed, T.A.; Qin, W. Fungal biodegradation and enzymatic modification of lignin. Int. J. Biochem. Mol. Biol. 2010, 1, 36–50. [Google Scholar] [PubMed]
- Amirta, R.; Tanabe, T.; Watanabe, T.; Honda, Y.; Kuwahara, M.; Watanabe, T. Methane fermentation of Japanese cedar wood pretreated with a white rot fungus Ceriporiopsis subvermispora. J. Biotechnol. 2006, 123, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Castillo, M.D.P.; Schnürer, A. Fungal Pretreatment of Straw for Enhanced Biogas Yield; Report No. 2013:279; Swedish Gas Technology Centre: Malmo, Sweden, 2013. [Google Scholar]
- Wan, C.; Li, Y. Fungal pretreatment of lignocellulosic biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass: An overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Galhaup, C.; Wagner, H.; Hinterstoisser, B.; Haltrich, D. Increased production of laccase by the wood-degrading basidiomycete Trametes pubescens. Enzym. Microb. Technol. 2002, 3, 529–536. [Google Scholar] [CrossRef]
- Buswell, J.A.; Cai, Y.J.; Chang, S.-T. Effect of nutrient nitrogen and manganese on manganese-peroxidase and laccase production of Lentinula (Lentinus) edodes. FEMS Microbiol. Lett. 1995, 128, 81–88. [Google Scholar] [CrossRef]
- Isroi; Millati, R.; Syamsiah, S.; Niklasson, C.; Cahyanto, M.N.; Lundquist, K.; Taherzadeh, M.J. Biological pretreatment of lignocelluloses with white-rot fungi and its applications: A review. Bioresources 2011, 6, 5224–5259. [Google Scholar]
- Mikiashvili, N.; Wasser, S.; Nevo, E.; Elisashvili, V. Effects of carbon and nitrogen sources on Pleurotus ostreatus ligninolytic enzyme activity. World J. Microbiol. Biotechnol. 2006, 22, 999–1002. [Google Scholar] [CrossRef]
- Sarkar, N.; Ghosh, S.K.; Bannerjee, S.; Aikat, K. Bioethanol production from agricultural wastes: An overview. Renew. Energy 2012, 37, 19–27. [Google Scholar] [CrossRef]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Combination of fungal and physicochemical processes for lignocellulosic biomass pretreatment—A review. Renew. Sustain. Energy Rev. 2016, 54, 217–234. [Google Scholar] [CrossRef]
- Owens, J.M.; Chynoweth, D.P. Biochemical methane potential of municipal solid waste (MSW) components. Water Sci. Technol. 1993, 27, 1–14. [Google Scholar]
- Skiadas, I.V.; Lyberatos, G. The periodic anaerobic baffled reactor. Water Sci. Technol. 1998, 38, 401–408. [Google Scholar] [CrossRef]
- Sluiter, A.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Extractives in Biomass: Laboratory Analytical Procedure; National Renewable Energy Laboratory: Golden, CO, USA, 2008.
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass: Laboratory Analytical Procedure; National Renewable Energy Laboratory: Golden, CO, USA, 2008.
- Joseffson, B. Rapid spectrophotometric determination of total carbohydrates. In Methods of Seawater Analysis; Grasshoff, K., Ehrhardt, M., Kremling, K., Eds.; Verlag Chemie GmbH: Weinheim, Germany, 1983; pp. 340–342. [Google Scholar]
- Waterman, P.G.; Mole, S. Analysis of phenolic plant metabolites. In Methods in Ecology; Lawton, J.H., Likens, G.E., Eds.; Oxford Blackwell Scientific Publications: London, UK, 1994. [Google Scholar]
- American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater; Franson, M.A., Ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Senkevich, S.; Ntaikou, I.; Lyberatos, G. Bioethanol production from thermochemically pre-treated olive mill solid residues using the yeast Pachysolen tannophilus. Glob. Nest J. 2012, 14, 118–124. [Google Scholar]
- Watkinson, S.; Bebber, D.; Darrah, P.; Fricker, M.; Tlalka, M.; Bodd, L. The role of wood decay fungi in the carbon and nitrogen dynamics of the forest floor. In Fungi in Biogeochemical Cycles; Gadd, G.M., Ed.; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Touratier, F.; Legendre, L.; Vezina, A. Model of bacterial growth influenced by substrate C:N ratio and concentration. Aquat. Microb. Ecol. 1999, 19, 105–118. [Google Scholar] [CrossRef]
- Allison, S.D.; LeBauer, D.S.; Ofrecio, M.R.; Reyes, R.; Ta, A.-M.; Tran, T.M. Low levels of nitrogen addition stimulate decomposition by boreal forest fungi. Soil Biol. Biochem. 2009, 41, 293–302. [Google Scholar] [CrossRef]
- D’Agostini, E.C.; Mantovani, T.R.D.; do Valle, J.S.; Paccola-Meirelles, L.D.; Colauto, N.B.; Linde, G.A. Low carbon/nitrogen ratio increases laccase production from basidiomycetes in solid substrate cultivation. Sci. Agric. 2011, 68, 295–300. [Google Scholar]
- Jurado, E.; Gavala, H.N.; Skiadas, I.V. Enhancement of methane yield from wheat straw, miscanthus and willow using aqueous ammonia soaking. Environ. Technol. 2013, 34, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Lalak, J.; Kasprzycka, A.; Martyniak, D.; Tys, J. Effect of biological pretreatment of Agropyronelongatum “BAMAR” on biogas production by anaerobic digestion. Bioresour. Technol. 2016, 200, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Vasco-Correa, J.; Li, Y. Solid-state anaerobic digestion of fungal pretreated Miscanthus sinensis harvested in two different seasons. Bioresour. Technol. 2015, 185, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Guo, X.M.; Latrille, E.; Trably, E.; Steyer, J.P.; Carrère, H. Predictive models of biohydrogen and biomethane production based on the compositional and structural features of lignocellulosic materials. Environ. Sci. Technol. 2012, 46, 12217–12225. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, K.; Wang, W.; Sun, R.-C. Improved bioconversion of poplar by synergistic treatments with white-rot fungus Trametes velutina D10149 pretreatment and alkaline fractionation. Bioresour. Technol. 2013, 130, 578–583. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Value |
|---|---|
| % TS a (w/w) | 94.53 ± 1.14 |
| % VS b (w/w) | 88.92 ± 1.21 |
| % Cellulose (w/w) | 35.59 ± 0.91 |
| % Hemicellulose (w/w) | 21.50 ± 0.89 |
| % Lignin (w/w) | 28.71 ± 0.23 |
| % Extractives (w/w) | 3.00 ± 0.13 |
| % Ash (w/w) | 5.91 ± 1.62 |
| % Proteins (w/w) | 0.69 ± 0.14 |
| % Total organic carbon (w/w) | 51.12 ± 0.93 |
| N Source | RC of Lignin 1 (%) | RC of Cellulose 1 (%) | RC of Hemicellulose 1 (%) | RC of Material Recovery 1 (%) |
|---|---|---|---|---|
| YE high | –0.97 ± 0.16 | +36.25 ± 2.05 | +0.52 ± 0.05 | –0.12 ± 0.01 |
| YE low | –1.77 ± 0.26 | +10.41 ± 1.66 | +1.66 ± 0.23 | +1.93 ± 0.22 |
| UR high | –21.61 ± 2.06 | +3.63 ± 0.38 | –13.23 ± 1.97 | –9.32 ± 1.38 |
| UR low | –1.82 ± 0.28 | +6.47 ± 1.00 | +8.52 ± 1.41 | +2.57 ± 0.35 |
| AN high | –2.43 ± 0.34 | +16.15 ± 1.09 | +5.28 ± 0.81 | +4.84 ± 1.01 |
| AN low | –4.19 ± 0.78 | +7.43 ± 0.99 | +4.28 ± 0.61 | +4.08 ± 0.58 |
| N Source | Phenols (mg/gTSin) | Sugars (mg/gTSin) |
|---|---|---|
| None (control) | 2.92 ± 0.12 | 22.60 ± 0.96 |
| YE high | 4.29 ± 0.18 | 27.56 ± 2.01 |
| YE low | 3.94 ± 0.07 | 27.17 ± 1.34 |
| UR high | 7.37 ± 0.13 | 11.50 ± 0.91 |
| UR low | 3.54 ± 0.15 | 24.08 ± 1.05 |
| AN high | 3.46 ± 0.11 | 37.60 ± 2.74 |
| AN low | 3.16 ± 0.11 | 27.02 ± 1.89 |
| N Source | RC of Lignin 1 (%) | RC of Cellulose 1 (%) | RC of Hemicellulose 1 (%) | RC of Material Recovery 1 (%) |
|---|---|---|---|---|
| YE high | –50.60 ± 3.12 | –24.78 ± 2.45 | –42.45 ± 0.90 | –29.26 ± 3.43 |
| YE low | –51.51 ± 2.45 | –21.96 ± 3.99 | –41.34 ± 3.65 | –29.06 ± 1.09 |
| UR high | –46.58 ± 5.56 | –22.21 ±2.48 | –38.12 ± 2.62 | –24.66 ± 2.74 |
| UR low | –54.50 ± 3.22 | –22.84 ± 4.01 | –43.29 ± 3.21 | –31.04 ± 4.18 |
| AN high | –53.81 ± 1.89 | –21.63 ± 5.32 | –44.41 ± 4.02 | –28.90 ± 0.93 |
| AN low | –55.42 ± 4.00 | –18.85 ± 3.45 | –42.98 ± 3.99 | –30.21 ± 1.11 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandropoulou, M.; Antonopoulou, G.; Ntaikou, I.; Lyberatos, G. Fungal Pretreatment of Willow Sawdust with Abortiporus biennis for Anaerobic Digestion: Impact of an External Nitrogen Source. Sustainability 2017, 9, 130. https://doi.org/10.3390/su9010130
Alexandropoulou M, Antonopoulou G, Ntaikou I, Lyberatos G. Fungal Pretreatment of Willow Sawdust with Abortiporus biennis for Anaerobic Digestion: Impact of an External Nitrogen Source. Sustainability. 2017; 9(1):130. https://doi.org/10.3390/su9010130
Chicago/Turabian StyleAlexandropoulou, Maria, Georgia Antonopoulou, Ioanna Ntaikou, and Gerasimos Lyberatos. 2017. "Fungal Pretreatment of Willow Sawdust with Abortiporus biennis for Anaerobic Digestion: Impact of an External Nitrogen Source" Sustainability 9, no. 1: 130. https://doi.org/10.3390/su9010130
APA StyleAlexandropoulou, M., Antonopoulou, G., Ntaikou, I., & Lyberatos, G. (2017). Fungal Pretreatment of Willow Sawdust with Abortiporus biennis for Anaerobic Digestion: Impact of an External Nitrogen Source. Sustainability, 9(1), 130. https://doi.org/10.3390/su9010130