
Relationships between Soil Crust Development and Soil Properties in the Desert Region of North China



Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Laboratory Analyses
2.4. Data Analyses
3. Results
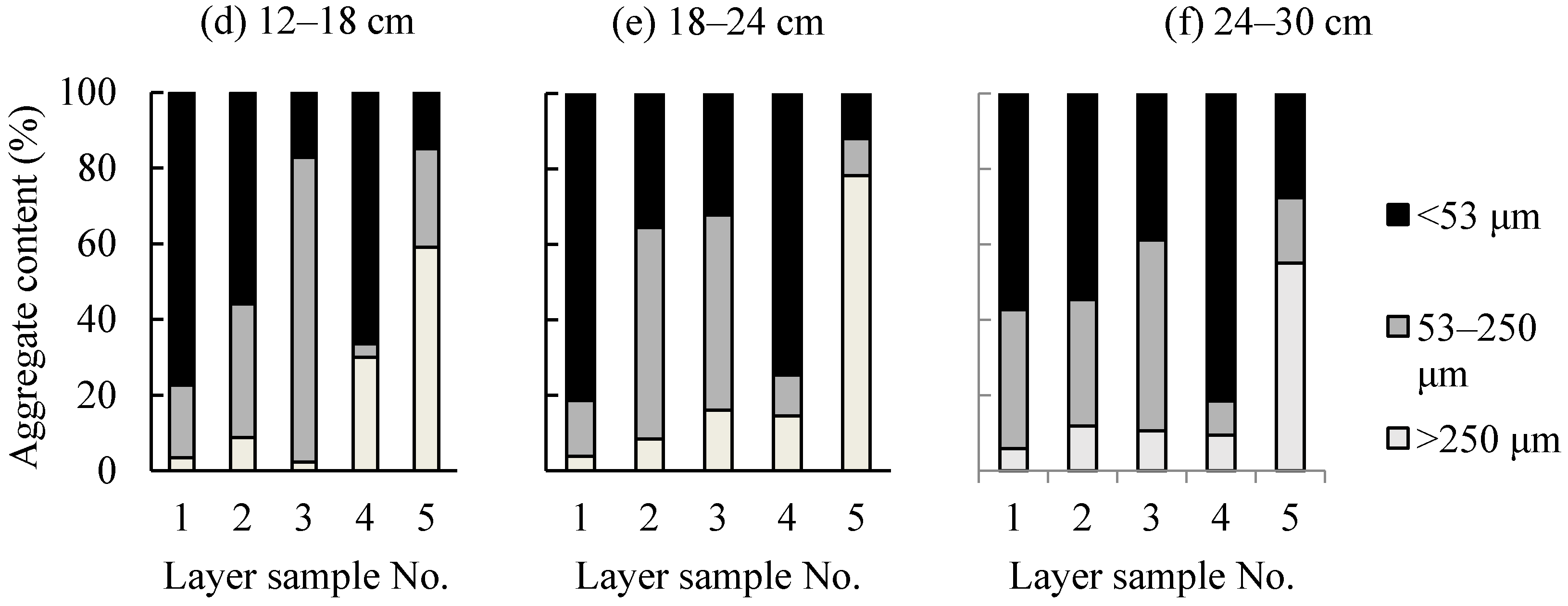
3.1. Soil Physicochemical Properties
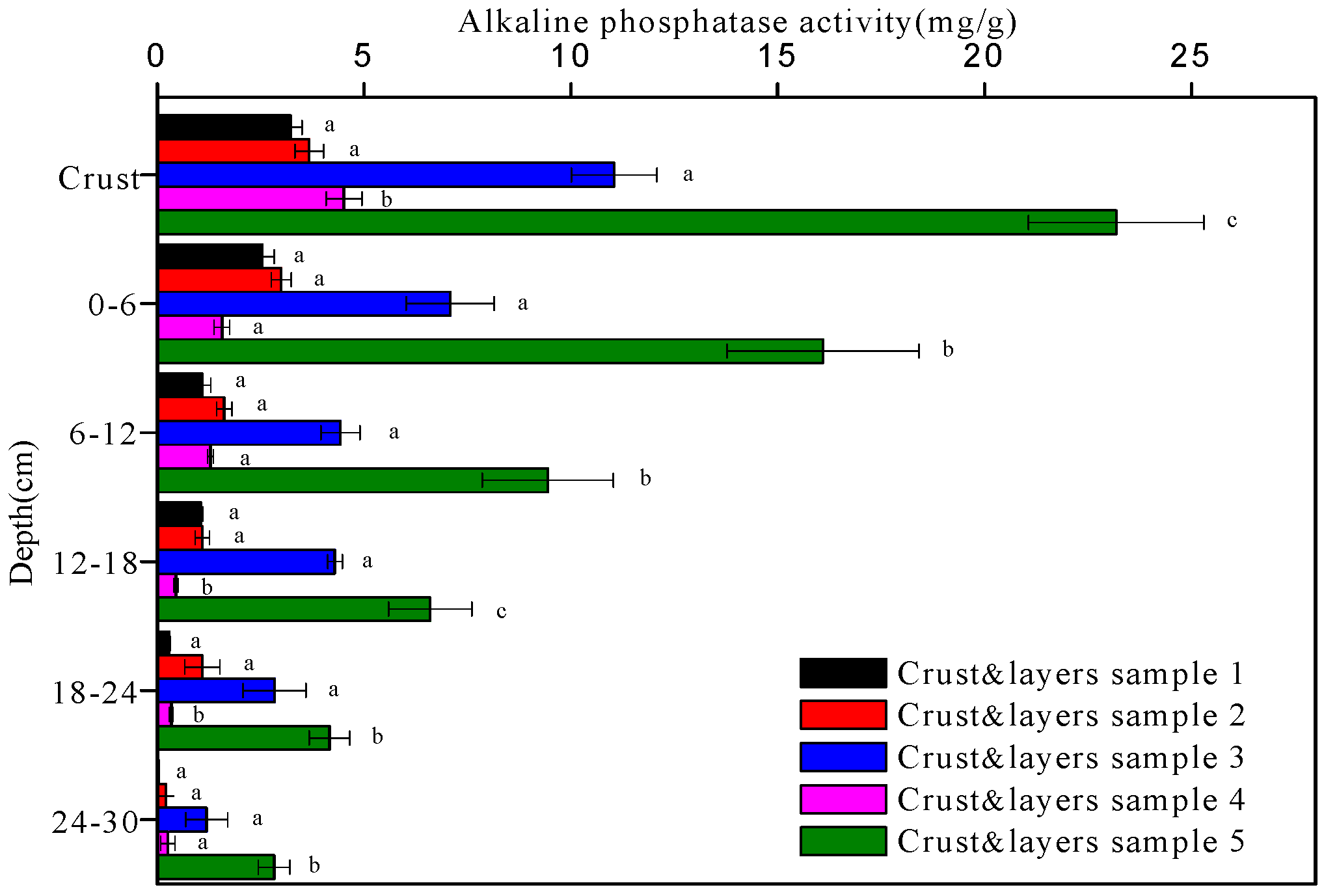
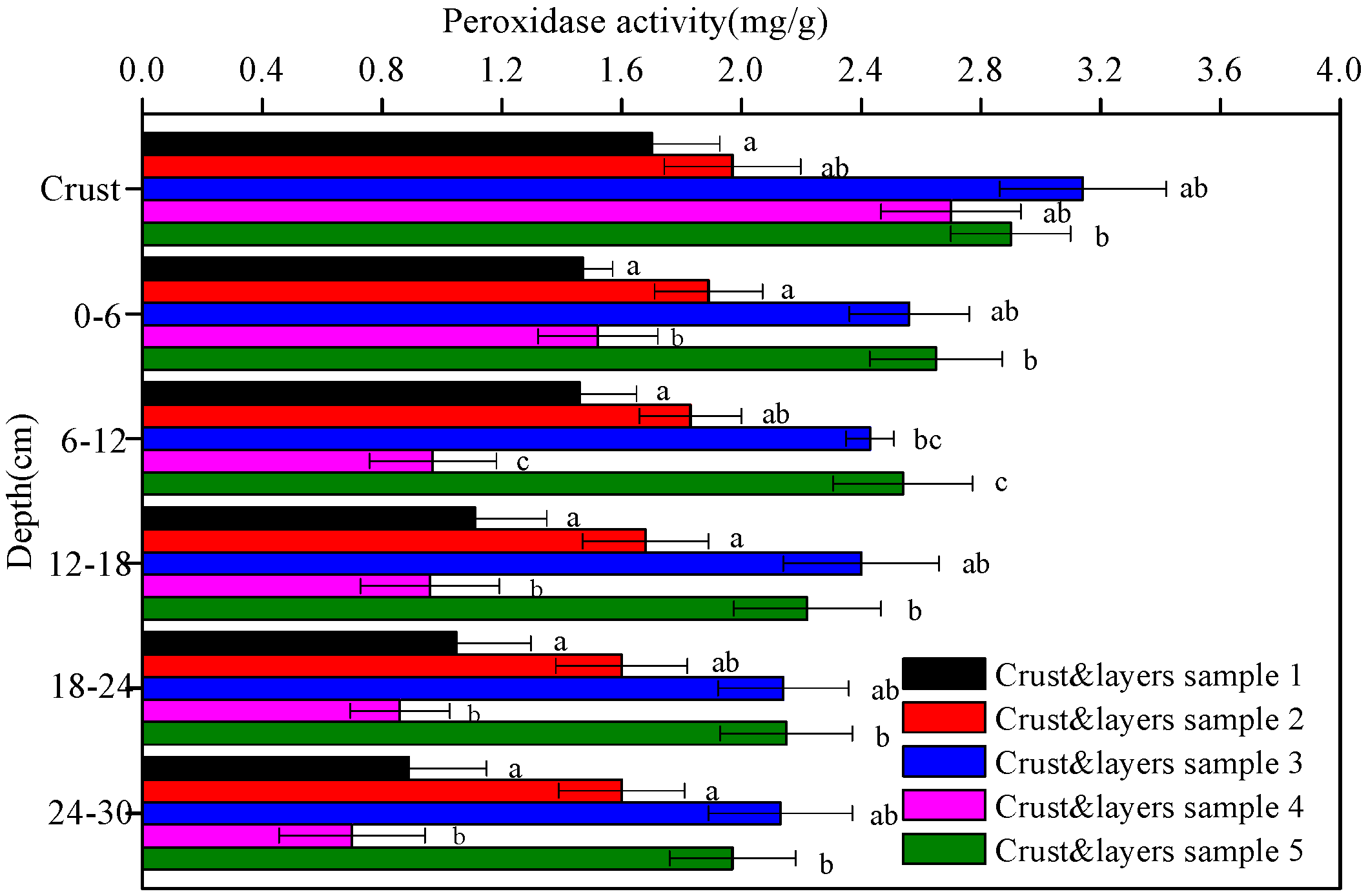
3.2. Enzyme Activities
3.3. Microbial Biomass Carbon and Nitrogen Contents
3.4. Relationships between Physicochemical Properties and Microbial Parameters of Soil Crusts
4. Discussion
4.1. Relationships between Soil Crust Development and Soil Properties
4.2. Relationships between Vegetation Types and Soil Properties
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guo, Y.; Zhao, H.; Zuo, X.; Drake, S.; Zhao, X. Biological soil crust development and its topsoil properties in the process of dune stabilization, Inner Mongolia, China. Environ. Geol. 2008, 54, 653–662. [Google Scholar] [CrossRef]
- Chamizo, S.; Cantón, Y.; Miralles, I.; Domingo, F. Biological soil crust development affects physicochemical characteristics of soil surface in semiarid ecosystems. Soil Biol. Biochem. 2012, 49, 96–105. [Google Scholar] [CrossRef]
- Enwall, K.; Nyberg, K.; Bertilsson, S.; Cederlund, H.; Stenström, J.; Hallin, S. Long-term impact of fertilization on activity and composition of bacterial communities and metabolic guilds in agricultural soil. Soil Biol. Biochem. 2007, 39, 106–115. [Google Scholar] [CrossRef]
- Prasse, R.; Bornkamm, R. Effect of microbiotic soil surface crusts on emergence of vascular plants. Plant Ecol. 2000, 150, 65–75. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Li, T.; Zhang, J. Microbiotic soil crust and its effect on vegetation and habitat on artificially stabilized desert dunes in Tengger Desert, North China. Biol. Fertil. Soils 2002, 35, 147–154. [Google Scholar]
- Eldridge, D.; Leys, J. Exploring some relationships between biological soil crusts, soil aggregation and wind erosion. J. Arid Environ. 2003, 53, 457–466. [Google Scholar] [CrossRef]
- Coxson, D. Biological soil crusts: Structure, function, and management. Bryologist 2002, 105, 500–501. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Wang, X.; Yang, W.; Zhang, D. The microstructure of microbiotic crust and its influence on wind erosion for a sandy soil surface in the Gurbantunggut Desert of Northwestern China. Geoderma 2006, 132, 441–449. [Google Scholar] [CrossRef]
- Eldridge, J.; Greene, R. Microbiotic soil crusts-a review of their roles in soil and ecological processes in the rangelands of Australia. Soil Res. 1994, 32, 389–415. [Google Scholar] [CrossRef]
- Maestre, F.; Bowker, M.; Cantón, Y.; Castillo-Monroy, A.; Cortina, J.; Escolar, C.; Escudero, A.; Lázaro, R.; Martínez, I. Ecology and functional roles of biological soil crusts in semi-arid ecosystems of Spain. J. Arid Environ. 2011, 75, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.; Thomas, A. Extracellular polysaccharides from cyanobacterial soil crusts: A review of their role in dryland soil processes. J. Arid Environ. 2011, 75, 91–97. [Google Scholar] [CrossRef]
- Kirk, J.; Beaudette, L.; Hart, M.; Moutoglis, P.; Klironomos, J.; Lee, H.; Trevors, J. Methods of studying soil microbial diversity. J. Microbiol. Methods. 2004, 58, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Chen, H.; Gong, Y.; Fan, M.; Yang, H.; Lal, R.; Kuzyakov, Y. Effects of 15 years of manure and inorganic fertilizers on soil organic carbon fractions in a wheat-maize system in the North China Plain. Nutr. Cycl. Agroecosyst. 2012, 92, 21–33. [Google Scholar] [CrossRef]
- Issa, O.; Trichet, J.; Défarge, C.; Couté, A.; Valentin, C. Morphology and microstructure of microbiotic soil crusts on a tiger bush sequence (Niger, Sahel). Catena 1999, 37, 175–196. [Google Scholar] [CrossRef]
- Yu, J.; Steinberger, Y. Vertical distribution of soil microbial biomass and its association with shrubs from the Negev Desert. J. Arid Environ. 2012, 78, 110–118. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Cruz, L.; Sotomayor-Ramirez, D.; Pérez-Alegría, L. Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl. Soil Ecol. 2007, 35, 35–45. [Google Scholar] [CrossRef]
- García, R.; Ochoa, V.; Viñegla, B.; Hinojosa, M.; Pena, R.; Liébanas, G.; Carreira, J. Soil enzymes, nematode community and selected physico-chemical properties as soil quality indicators in organic and conventional olive oil farming: influence of seasonality and site features. Appl. Soil Ecol. 2009, 41, 305–314. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Wang, X.; Liu, L.; Xiao, H. Study on soil microbiotic crust and its influences on sand-fixing vegetation in arid desert region. Acta Botanica Sin. 1999, 42, 965–970. [Google Scholar]
- Chen, L.; Xie, Z.; Hu, C.; Li, D.; Wang, G.; Liu, Y. Man-made desert algal crusts as affected by environmental factors in Inner Mongolia, China. J. Arid Environ. 2006, 67, 521–527. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, M.; Belnap, J. Potential nitrogen fixation activity of different aged biological soil crusts from rehabilitated grasslands of the hilly Loess Plateau, China. J. Arid Environ. 2010, 74, 1186–1191. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, G.; Liu, G.; Dong, Z.; Chen, T.; Zhang, M.; An, L. Bacterial diversity and distribution in the southeast edge of the Tengger Desert and their correlation with soil enzyme activities. J. Environ. Sci. 2012, 24, 2004–2011. [Google Scholar] [CrossRef]
- Chen, X.; Duan, Z. Impacts of soil crusts on soil physicochemical characteristics in different rainfall zones of the arid and semi-arid desert regions of northern China. Environ. Earth Sci. 2015, 73, 3335–3347. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci. Soc. Am. J. 1993, 57, 1071–1076. [Google Scholar] [CrossRef]
- Nelson, D.; Sommers, L. Total carbon, organic carbon, and organic matter. Methods of soil analysis. Part 2. Chem. Microbiol. Prop. 1982, 2, 539–579. [Google Scholar]
- Witt, C.; Gaunt, J.; Galicia, C.; Ottow, J.; Neue, H. A rapid chloroform-fumigation extraction method for measuring soil microbial biomass carbon and nitrogen in flooded rice soils. Biol. Fertil. Soils 2000, 30, 510–519. [Google Scholar] [CrossRef]
- Guan, S. Soil Enzymes and its Research Methods; Agriculture Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Wardle, D. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil. Biol. Rev. 1992, 67, 321–358. [Google Scholar] [CrossRef]
- Belnap, J.; Eldridge, D. Disturbance and recovery of biological soil crusts. In Biological Soil Crusts: Structure, Function, and Management; (Ecological Studies 150); Springer: Berlin/Heidelberg, Germany, 2001; pp. 363–383. [Google Scholar]
- Shamir, I.; Steinberger, Y. Vertical distribution and activity of soil microbial population in a sandy desert ecosystem. Microb. Ecol. 2007, 53, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Chenu, C.; Guerif, J. Mechanical strength of clay minerals as influenced by an adsorbed polysaccharide. Soil Sci. Soc. Am. J. 1991, 55, 1076–1080. [Google Scholar] [CrossRef]
- Yang, Z.; Hao, J.; Miao, G. Effect of immature loess subsoil fertilization in current year on rhizospheric soil biological activity and nutrient of mixture cropping. J. Soil Water Conserv. 2010, 24, 223–227. [Google Scholar]
- Pajares, S.; Gallardo, J.; Masciandaro, G.; Ceccanti, B.; Etchevers, J. Enzyme activity as an indicator of soil quality changes in degraded cultivated Acrisols in the Mexican Trans-Volcanic Belt. Land Degrad. Dev. 2011, 22, 373–381. [Google Scholar] [CrossRef]
- Bolton, H.; Smith, J.L.; Link, S.O. Soil microbial biomass and activity of a disturbed and undisturbed shrub-steppe ecosystem. Soil Biol. Biochem. 1993, 25, 545–552. [Google Scholar] [CrossRef]
- Jastrow, J.; Miller, R.; Boutton, T. Carbon dynamics of aggregate-associated organic matter estimated by carbon-13 natural abundance. Soil Sci. Soc. Am. J. 1996, 60, 801–807. [Google Scholar] [CrossRef]
- Zi, C.Z.; Shu, Q.H.; Ting, X.L.; Yong, D.W. Effect of land use patterns on stability and distributions of organic carbon in the hilly region of Western Sichuan, China. Afr. J. Biotechnol. 2013, 10, 13107–13114. [Google Scholar]
- Franzluebbers, A.; Arshad, M. Soil organic matter pools with conventional and zero tillage in a cold, semiarid climate. Soil Tillage Res. 1996, 39, 1–11. [Google Scholar] [CrossRef]
- Wezel, A.; Rajot, J.; Herbrig, C. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. J. Arid Environ. 2000, 44, 383–398. [Google Scholar] [CrossRef]
- Schlesinger, W.; Pilmanis, A. Plant-soil interactions in deserts. Biogeochemistry 1998, 42, 169–187. [Google Scholar] [CrossRef]
- Read, C.; Duncan, D.; Vesk, P.; Elith, J. Biological soil crust distribution is related to patterns of fragmentation and landuse in a dryland agricultural landscape of southern Australia. Landsc. Ecol. 2008, 23, 1093–1105. [Google Scholar] [CrossRef]
- Thomas, A.; Dougill, A. Distribution and characteristics of cyanobacterial soil crusts in the Molopo Basin, South Africa. J. Arid Environ. 2006, 64, 270–283. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Dominant Vegetation | Crust Type | Color | Thickness (cm) |
|---|---|---|---|---|
| 1 | Artemisia Ordosica | Physical Crust | Light-Colored | 0.41 |
| 2 | Artemisia Ordosica | Algal Crust | Dull Gray | 0.62 |
| 3 | Artemisia Ordosica | Moss Crust | Yellow Green | 1.53 |
| 4 | Artemisia Ordosica and Eragrostis Poaeoides | Algal Crust | Brown | 0.57 |
| 5 | Artemisia Ordosica and Eragrostis Poaeoides | Moss Crust | Yellow Green | 1.72 |
| SWC | SBD | SOMC | SMBC | SMBN | Alkaline Phosphatase | Urease | Protease | Peroxidase | |
|---|---|---|---|---|---|---|---|---|---|
| SWC a | 1.000 | −0.801 | 0.916 * | 0.956 * | 0.935 * | 0.959 ** | 0.872 | 0.914 * | 0.540 |
| SBD b | 1.000 | −0.813 | −0.854 | −0.918 * | −0.828 | −0.775 | −0.694 | −0.868 | |
| SOMC c | 1.000 | 0.926 * | 0.911 * | 0.977 ** | 0.855 | 0.959 ** | 0.732 | ||
| SMBC d | 1.000 | 0.989 ** | 0.917 * | 0.972 ** | 0.942 * | 0.700 | |||
| SMBN e | 1.000 | 0.906 * | 0.953 * | 0.895 * | 0.768 | ||||
| Alkaline Phosphatase | 1.000 | 0.809 | 0.919 * | 0.646 | |||||
| Urease | 1.000 | 0.920 * | 0.694 | ||||||
| Protease | 1.000 | 0.616 | |||||||
| Peroxidase | 1.000 |
| Source of Variation | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enzymes | Crust Type | Vegetation Cover | Depth | Crust Type × Vegetation Cover | Crust Type × Depth | Vegetation Cover × Depth | Crust Type × Vegetation Cover × Depth | |||||||
| F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | |
| Alkaline Phosphatase | 205 | 0.000 | 59 | 0.000 | 78 | 0.000 | 79 | 0.000 | 16 | 0.000 | 9.3 | 0.000 | 7.6 | 0.000 |
| Urease | 174 | 0.000 | 114 | 0.000 | 46 | 0.000 | 22 | 0.000 | 2.7 | 0.016 | 6.9 | 0.000 | 6.5 | 0.000 |
| Protease | 119 | 0.000 | 51 | 0.000 | 308 | 0.000 | 82 | 0.000 | 11 | 0.000 | 12 | 0.000 | 17 | 0.000 |
| Peroxidase | 83 | 0.000 | 9.3 | 0.005 | 15 | 0.000 | 5.5 | 0.026 | 0.3 | 0.982 | 1.7 | 0.162 | 2.6 | 0.044 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, J.; Yang, K.; Tang, Z.; Wang, Y. Relationships between Soil Crust Development and Soil Properties in the Desert Region of North China. Sustainability 2017, 9, 725. https://doi.org/10.3390/su9050725
Niu J, Yang K, Tang Z, Wang Y. Relationships between Soil Crust Development and Soil Properties in the Desert Region of North China. Sustainability. 2017; 9(5):725. https://doi.org/10.3390/su9050725
Chicago/Turabian StyleNiu, Jiping, Kai Yang, Zejun Tang, and Yitong Wang. 2017. "Relationships between Soil Crust Development and Soil Properties in the Desert Region of North China" Sustainability 9, no. 5: 725. https://doi.org/10.3390/su9050725
APA StyleNiu, J., Yang, K., Tang, Z., & Wang, Y. (2017). Relationships between Soil Crust Development and Soil Properties in the Desert Region of North China. Sustainability, 9(5), 725. https://doi.org/10.3390/su9050725