Grain Yield, Dry Weight and Phosphorus Accumulation and Translocation in Two Rice (Oryza sativa L.) Varieties as Affected by Salt-Alkali and Phosphorus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Experimental Set-Up
2.2. Experimental Sampling and Measurement
2.3. Statistical Analysis
3. Results
3.1. DW Production
3.2. P Accumulation
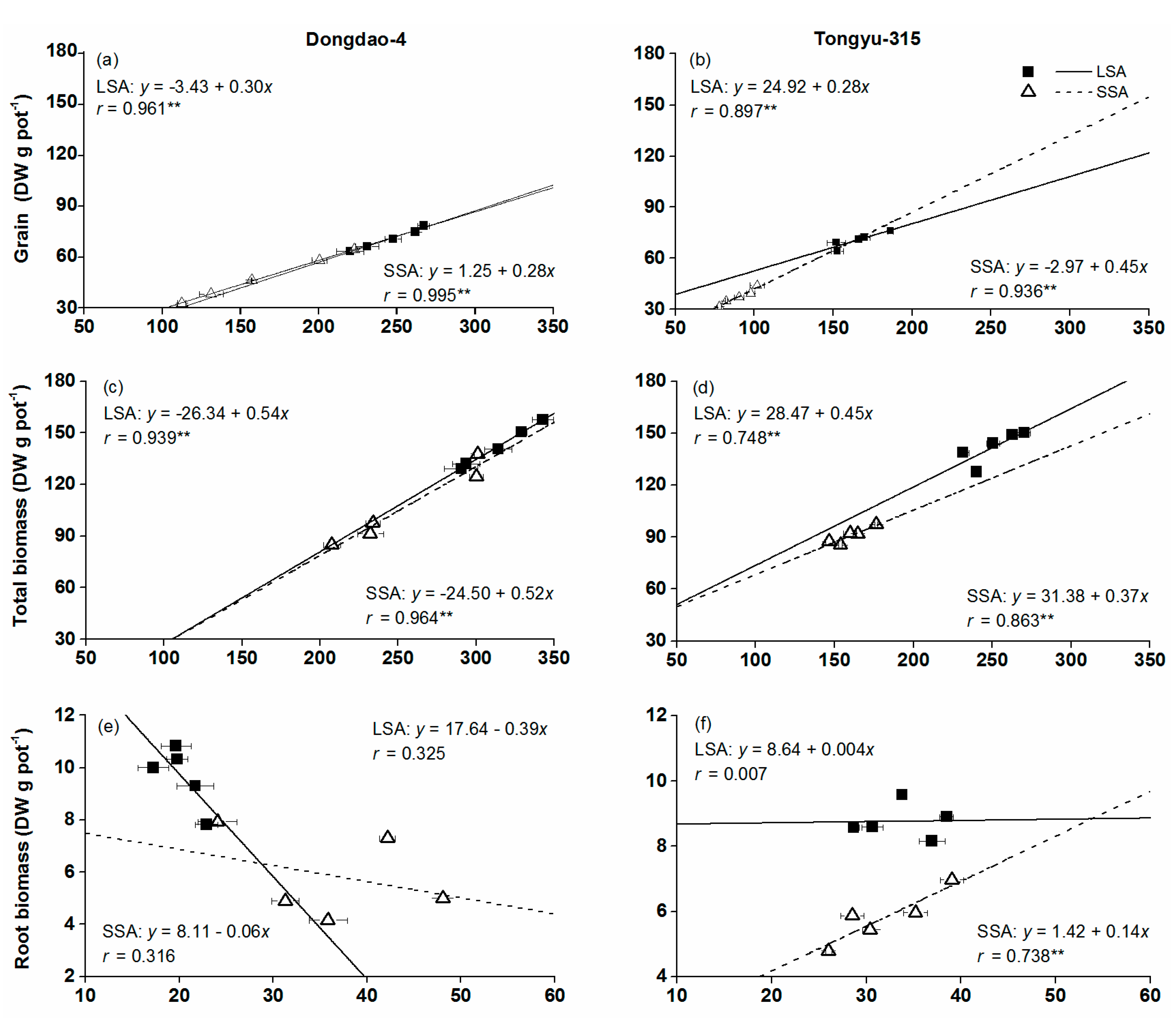
3.3. Relationship between DW and P Content at Maturity
3.4. The 1000-Grain Weight and SR
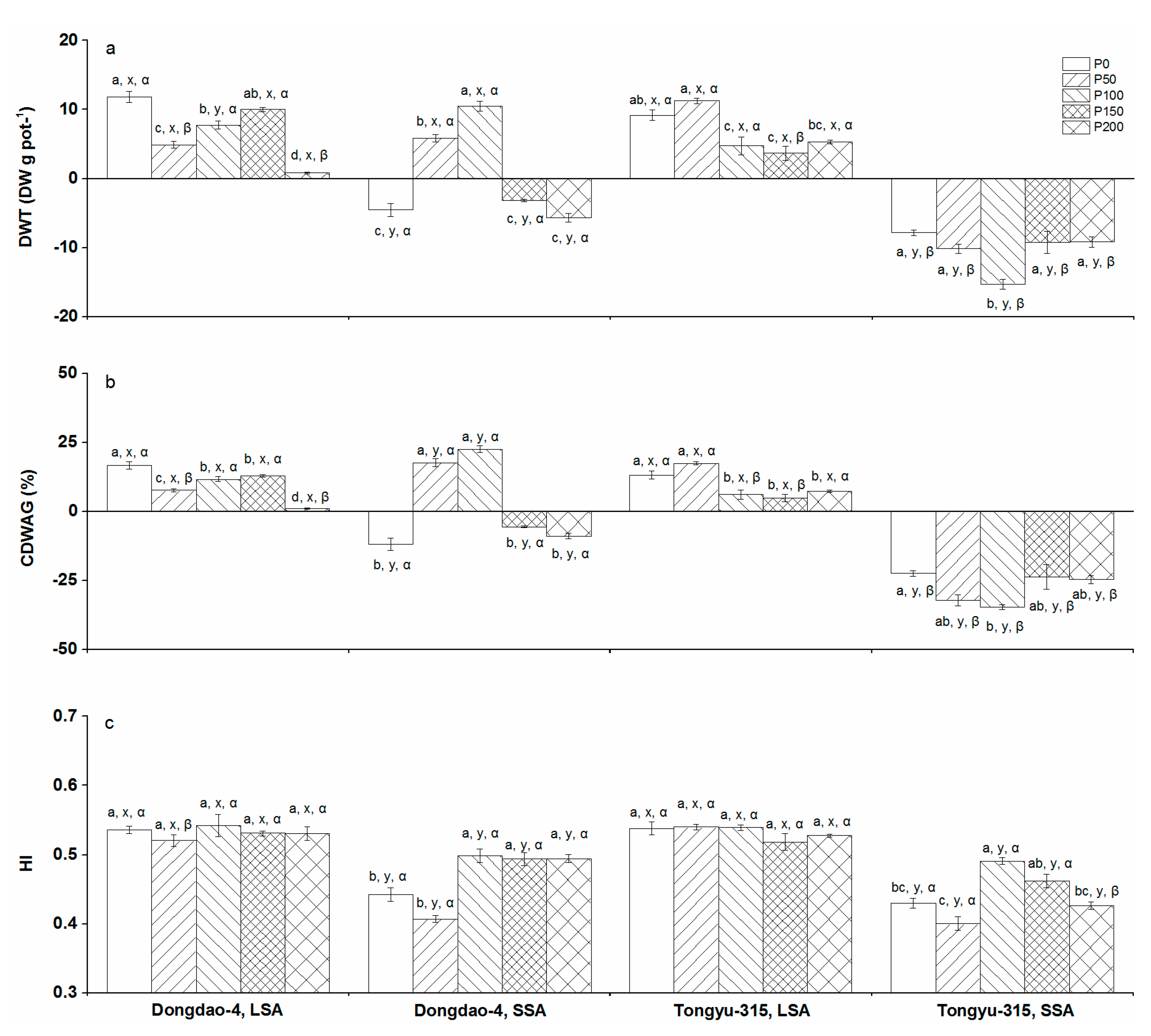
3.5. DW Translocation
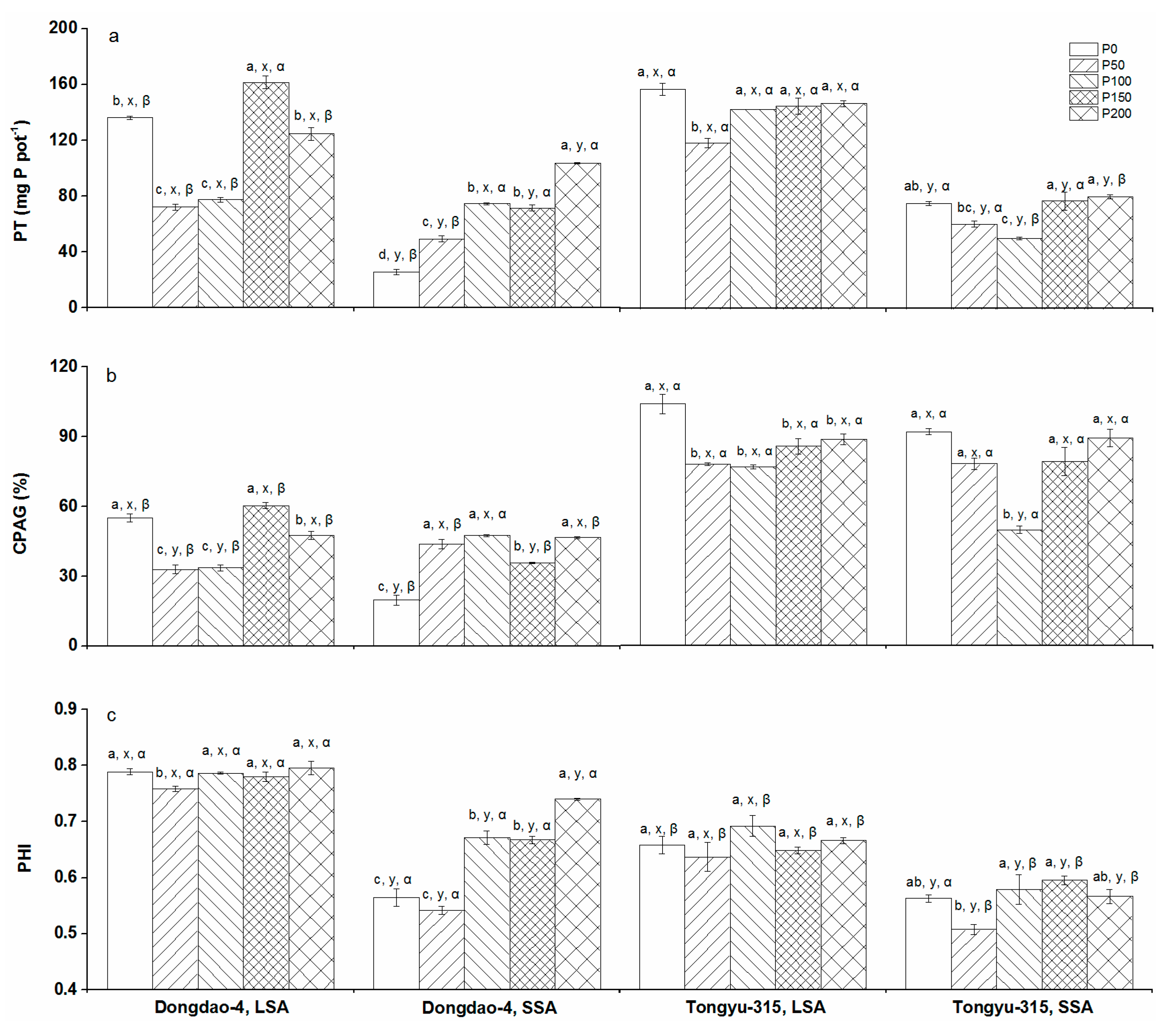
3.6. P Translocation
4. Discussion
4.1. DW and P Accumulation
4.2. Optimal P Application
4.3. DW and P Translocation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flowers, T.J.; Yeo, A.R. Breeding for salinity resistance in crop plants: Where next? Aust. J. Plant Physiol. 1995, 22, 875–884. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wu, Z.J.; Shi, Y.L.; Wang, R.Y. The resource of saline soil in the Northest China. Chin. J. Soil Sci. 2004, 5, 643–647. (In Chinese) [Google Scholar]
- Dahiya, S.; Singh, M. Effect of salinity, alkalinity and iron application on the availability of iron, manganese, phosphorus and sodium in pea (Pisum sativum L.) crop. Plant Soil 1976, 44, 697–702. [Google Scholar] [CrossRef]
- Lv, B.S.; Li, X.W.; Ma, H.Y.; Sun, Y.; Wei, L.X.; Jiang, C.J.; Liang, Z.W. Differences in growth and physiology of rice in response to different saline-alkaline stress factors. Agron. J. 2013, 105, 1119–1128. [Google Scholar] [CrossRef]
- Fageria, N.K.; Barbosa, M.P. Dry-matter and grain yield, nutrient uptake, and phosphorus use-efficiency of lowland rice as influenced by phosphorus fertilization. Commun. Soil Sci. Plan Anal. 2007, 38, 1289–1297. [Google Scholar] [CrossRef]
- Ramaekers, L.; Remans, R.; Rao, I.M.; Blair, M.W.; Vanderleyden, J. Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crops Res. 2010, 117, 169–176. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Wang, J.L.; Shuman, L.M. Transformation of phosphate in rice (Oryza sativa L) rhizosphere and its influence on phosphorus-nutrition of rice. J. Plant Nutr. 1994, 17, 1803–1815. [Google Scholar] [CrossRef]
- Qadir, M.; Noble, A.D.; Oster, J.D.; Schubert, S.; Ghafoor, A. Driving forces for sodium removal during phytoremediation of calcareous sodic and saline-sodic soils: A review. Soil Use Manag. 2005, 21, 173–180. [Google Scholar] [CrossRef]
- Sanaullah, M.; Rumpel, C.; Charrier, X.; Chabbi, A. How does drought stress influence the decomposition of plant litter with contrasting quality in a grassland ecosystem? Plant Soil 2012, 352, 277–288. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Ann. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Masoni, A.; Ercoli, L.; Mariotti, M.; Arduini, I. Post-anthesis accumulation and remobilization of dry matter, nitrogen and phosphorus in durum wheat as affected by soil type. Eur. J. Agron. 2007, 26, 179–186. [Google Scholar] [CrossRef]
- Grattan, S.R.; Grieve, C.M. Salinity mineral nutrient relations in horticultural crops. Sci. Hortic Amst. 1999, 78, 127–157. [Google Scholar] [CrossRef]
- Sairam, R.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. India 2004, 86, 407–421. [Google Scholar]
- Shabbir, G.; Hussain, N.; Bhatti, M.; Ahmad, A.; Javed, M.; Shakir, M.A. Salt tolerance potential of some selected fine rice cultivars. Field Stud. 2001, 1997, 99. [Google Scholar]
- Aslam, M.; Flowers, T.; Qureshi, R.; Yeo, A. Interaction of phosphate and salinity on the growth and yield of rice (Oryza sativa L.). J. Agron. Crop Sci. 1996, 176, 249–258. [Google Scholar] [CrossRef]
- Flowers, T.J.; Flowers, S.A. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Tian, Z.J.; Li, J.P.; Jia, X.Y.; Yang, F.; Wang, Z.C. Assimilation and translocation of dry matter and phosphorus in rice genotypes affected by salt-alkaline stress. Sustainability 2016, 8, 568. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Fabre, D.; Siband, P.; Dingkuhn, M. Characterizing stress effects on rice grain development and filling using grain weight and size distribution. Field Crops Res. 2005, 92, 11–16. [Google Scholar] [CrossRef]
- Tahir, I.S.A.; Nakata, N. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop Sci. 2005, 191, 106–115. [Google Scholar] [CrossRef]
- Elgharably, A. Wheat response to combined application of nitrogen and phosphorus in a saline sandy loam soil. Soil Sci. Plant Nutr. 2011, 57, 396–402. [Google Scholar] [CrossRef]
- Naheed, G.; Shahbaz, M.; Latif, A.; Rha, E.S. Alleviation of the adverse effects of salt stress on rice (Oryza sativa L.) by phosphorus applied through rooting medium: Growth and gas exchange characteristics. Pak. J. Bot. 2007, 39, 729–737. [Google Scholar]
- Awad, A.; Edwards, D.; Campbell, L. Phosphorus enhancement of salt tolerance of tomato. Crop Sci. 1990, 30, 123–128. [Google Scholar] [CrossRef]
- Usman, K. Effect of phosphorus and irrigation levels on yield, water productivity, phosphorus use efficiency and income of lowland rice in northwest Pakistan. Rice Sci. 2013, 20, 61–72. [Google Scholar] [CrossRef]
- Alam, M.; Ali, M.; Amin, A.; Hasanuzzaman, M. Yield attributes, yield and harvest index of three irrigated rice varieties under different levels of phosphorus. Adv. Biol. Res. 2009, 3, 132–139. [Google Scholar]
- Nieman, R.H.; Clark, R.A. Interactive effects of salinity and phosphorus nutrition on concentrations of phosphate and phosphate esters in mature photosynthesizing corn leaves. Plant Physiol. 1976, 57, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Dewit, C.T. Resource use efficiency in agriculture. Agric. Syst. 1992, 40, 125–151. [Google Scholar] [CrossRef]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source-sink relations. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Wissuwa, M.; Yano, M.; Ae, N. Mapping of QTLs for phosphorus-deficiency tolerance in rice (Oryza sativa L.). Theor. Appl. Genet. 1998, 97, 777–783. [Google Scholar] [CrossRef]
- Gao, J.P.; Chao, D.Y.; Lin, H.X. Toward understanding molecular mechanisms of abiotic stress responses in rice. Rice 2008, 1, 36–51. [Google Scholar] [CrossRef]
- Lin, W.Y.; Lin, S.I.; Chiou, T.J. Molecular regulators of phosphate homeostasis in plants. J. Exp. Bot. 2009, 60, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Weeks, R.; Tong, Y.; Davies, T.E.; Leggewie, G. Restricted spatial expression of a high-affinity phosphate transporter in potato roots. J. Cell Sci. 2003, 116, 3135–3144. [Google Scholar] [CrossRef] [PubMed]
- Raghothama, K.G.; Karthikeyan, A.S. Phosphate acquisition. Plant Soil 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Yuan, H.; Liu, D. Signaling components involved in plant responses to phosphate starvation. J. Integr. Plant Biol. 2008, 50, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chu, C. Phosphate starvation signaling in rice. Plant Signal. Behav. 2011, 6, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; Gonzalez, E.; Bustos, R.; Linhares, F.; Leyva, A.; Paz-Ares, J. The transcriptional control of plant responses to phosphate limitation. J. Exp. Bot. 2004, 55, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Sima, N.A.K.K.; Ahmad, S.T.; Alitabar, R.A.; Mottaghi, A.; Pessarakli, M. Interactive effects of salinity and phosphorus nutrition on physiological responses of two barley species. J. Plant Nutr. 2012, 35, 1411–1428. [Google Scholar] [CrossRef]
- Mohammadi-Nejad, G.; Singh, R.; Arzani, A.; Rezaie, A.; Sabouri, H.; Gregorio, G. Evaluation of salinity tolerance in rice genotypes. Int. J. Plant Prod. 2010, 4, 199–207. [Google Scholar]
- Jin, J.; Wang, G.H.; Liu, X.B.; Pan, X.W.; Herbert, S.J.; Tang, C.X. Interaction between phosphorus nutrition and drought on grain yield, and assimilation of phosphorus and nitrogen in two soybean cultivars differing in protein concentration in grains. J. Plant Nutr. 2006, 29, 1433–1449. [Google Scholar] [CrossRef]
- Martinez, V.; Lächli, A. Phosphorus translocation in salt-stressed cotton. Physiol. Plant. 1991, 83, 627–632. [Google Scholar] [CrossRef]
- Navarro, J.M.; Botella, M.A.; Cerdá, A.; Martinez, V. Phosphorus uptake and translocation in salt-stressed melon plants. J. Plant Physiol. 2001, 158, 375–381. [Google Scholar] [CrossRef]
- Rogers, M.E.; Grieve, C.M.; Shannon, M.C. Plant growth and ion relations in lucerne (Medicago sativa L.) in response to the combined effects of NaCl and P. Plant Soil 2003, 253, 187–194. [Google Scholar] [CrossRef]
- Gibson, T.S. Carbohydrate-metabolism and phosphorus salinity interactions in wheat (Triticum-Aestivum L.). Plant Soil 1988, 111, 25–35. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Sato, A.M.; Catuchi, T.A.; Ribeiro, R.V.; Souza, G.M. The use of network analysis to uncover homeostatic responses of a drought-tolerant sugarcane cultivar under severe water deficit and phosphorus supply. Acta Physiol. Plant 2010, 32, 1145–1151. [Google Scholar] [CrossRef]
- Tran, H.T.; Hurley, B.A.; Plaxton, W.C. Feeding hungry plants: The role of purple acid phosphatases in phosphate nutrition. Plant Sci. 2010, 179, 14–27. [Google Scholar] [CrossRef]
- Zribi, O.T.; Houmani, H.; Kouas, S.; Slama, I.; Ksouri, R.; Abdelly, C. Comparative study of the interactive effects of salinity and phosphorus availability in wild (Hordeum maritimum) and cultivated barley (H-vulgare). J. Plant Growth Regul. 2014, 33, 860–870. [Google Scholar] [CrossRef]
- Qadir, M.; Schubert, S. Degradation processes and nutrient constraints in sodic soils. Land Degrad. Dev. 2002, 13, 275–294. [Google Scholar] [CrossRef]
- Mahmood, I.A.; Ali, A. Response of direct seeded rice and wheat crops to phosphorus application with crop residue incorporation in saline-sodic soil. Int. J. Agric. Biol. 2015, 17, 1219–1224. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T. Effects of salinity on seed set in rice. Plant Cell Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar] [CrossRef]
- Arduini, I.; Masoni, A.; Ercoli, L.; Mariotti, M. Grain yield, and dry matter and nitrogen accumulation and remobilization in durum wheat as affected by variety and seeding rate. Eur. J. Agron. 2006, 25, 309–318. [Google Scholar] [CrossRef]
- Fang, Y.; Xu, B.C.; Turner, N.C.; Li, F.M. Grain yield, dry matter accumulation and remobilization, and root respiration in winter wheat as affected by seeding rate and root pruning. Eur. J. Agron. 2010, 33, 257–266. [Google Scholar] [CrossRef]
- Papakosta, D.K. Phosphorus accumulation and translocation in wheat as affected by cultivar and nitrogen fertilization. J. Agron. Crop Sci. 1994, 173, 260–270. [Google Scholar] [CrossRef]
- Panigrahy, M.; Rao, D.N.; Sarla, N. Molecular mechanisms in response to phosphate starvation in rice. Biotechnol. Adv. 2009, 27, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Abdullah, Z. Salinity–sodicity induced changes in reproductive physiology of rice (Oryza sativa L.) under dense soil conditions. Environ. Exp. Bot. 2003, 49, 145–157. [Google Scholar] [CrossRef]
- Naheed, G.; Shahbaz, M.; Akram, N.A. Interactive effect of rooting medium application of phosphorus and NaCl on plant biomass and mineral nutrients of rice (Oryza sativa L.). Pak. J. Bot. 2008, 40, 1601–1608. [Google Scholar]
- Abdullah, Z.; Khan, M.A.; Flowers, T. Causes of sterility in seed set of rice under salinity stress. J. Agron. Crop Sci. 2001, 187, 25–32. [Google Scholar] [CrossRef]
- Przulj, N.; Momcilovic, V. Genetic variation for dry matter and nitrogen accumulation and translocation in two-rowed spring barley II. Nitrogen translocation. Eur. J. Agron. 2001, 15, 255–265. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | pH | EC μS cm−1 | ENa+ cmol kg−1 | CEC cmol kg−1 | Organic Matter % | Total N mg kg−1 | Available N mg kg−1 | Total P mg kg−1 | Available P mg kg−1 |
|---|---|---|---|---|---|---|---|---|---|
| LSA | 8.27 | 271.0 | 1.75 | 18.00 | 1.77 | 740.4 | 153.9 | 384.0 | 35.15 |
| SSA | 9.09 | 395.7 | 3.19 | 14.81 | 1.18 | 615.1 | 88.9 | 344.2 | 28.60 |
| P Level | Soils | Shoot (DW g pot−1) | Root (DW g pot−1) | Total (DW g pot−1) | |||
|---|---|---|---|---|---|---|---|
| Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | ||
| P0 | LSA | 70.04 ± 1.08 | 67.97 ± 1.58 | 11.94 ± 0.13 | 10.78 ± 0.41 | 81.98 ± 1.16 | 78.75 ± 1.88 |
| b, x, α | ab, x, α | a, x, α | a, x, α | b, x, α | a, x, α | ||
| SSA | 42.13 ± 1.22 | 39.52 ± 1.2 | 6.40 ± 0.20 | 4.92 ± 0.15 | 48.53 ± 1.40 | 44.44 ± 1.42 | |
| c, y, α | 8ab, y, α | bc, y, α | b, y, β | c, y, α | ab, y, α | ||
| P50 | LSA | 61.56 ± 1.34 | 62.88 ± 1.14 | 9.33 ± 0.21 | 11.78 ± 0.32 | 70.89 ± 1.55 | 74.66 ± 1.33 |
| d, x, α | b, x, α | b, x, β | a, x, α | d, x, α | a, x, α | ||
| SSA | 50.62 ± 1.43 | 37.49 ± 1.33 | 7.33 ± 0.16 | 6.11 ± 0.37 | 57.95 ± 1.59 | 43.60 ± 1.67 | |
| b, y, α | ab, y, β | ab, y, α | a, y, β | b, y, α | ab, y, β | ||
| P100 | LSA | 64.06±0.57 | 67.77 ± 1.01 | 9.63 ± 0.35 | 11.09 ± 0.11 | 73.69 ± 0.86 | 78.86 ± 1.03 |
| cd, x, β | ab, x, α | b, x, β | a, x, α | cd, x, β | a, x, α | ||
| SSA | 54.22 ± 0.95 | 33.28 ± 0.54 | 7.65 ± 0.30 | 4.53 ± 0.06 | 61.88 ± 1.12 | 37.81 ± 0.57 | |
| b, y, α | b, y, β | a, y, α | a, y, β | ab, y, α | b, y, β | ||
| P150 | LSA | 78.69 ± 1.62 | 69.09 ± 0.58 | 11.06 ± 0.30 | 11.27 ± 0.51 | 89.75 ± 1.52 | 80.36 ± 1.07 |
| a, x, α | a, x, β | a, x, α | a, x, α | a, x, α | a, x, β | ||
| SSA | 55.69±1.18 | 38.49 ± 2.98 | 7.77 ± 0.10 | 4.53 ± 0.23 | 63.46 ± 1.08 | 43.02 ± 2.77 | |
| b, y, α | ab, y, β | a, y, α | a, y, β | ab, y, α | ab, y, β | ||
| P200 | LSA | 67.77 ± 0.74 | 66.13 ± 1.2 | 8.92 ± 0.29 | 12.09 ± 0.17 | 76.70 ± 0.99 | 78.22 ± 1.10 |
| bc, x, α | 6ab, x, α | b, x, β | a, x, α | bc, x, α | a, x, α | ||
| SSA | 61.57 ± 1.05 | 41.58 ± 1.40 | 6.29 ± 0.20 | 4.37 ± 0.14 | 67.86 ± 1.25 | 45.96 ± 1.49 | |
| a, y, α | a, y, β | c, y, α | a, y, β | a, y, α | a, y, β | ||
| P Level | Soils | Shoot (DW g pot−1) | Grain (DW g pot−1) | Root (DW g pot−1) | Total (DW g pot−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | ||
| P0 | LSA | 60.87 ± 0.48 | 59.99 ± 1.28 | 70.35 ± 2.00 | 69.24 ± 1.34 | 9.31 ± 0.33 | 9.58 ± 0.28 | 140.54 ± 2.73 | 138.81 ± 1.42 |
| b, x, α | c, x, α | bc, x, α | bc, x, α | b, x, α | a, x, α | bc, x, α | b, x, α | ||
| SSA | 48.10 ± 0.46 | 46.84 ± 0.82 | 38.22 ± 1.33 | 35.11 ± 0.39 | 5.00 ± 0.15 | 5.44 ± 0.19 | 91.32 ± 1.33 | 87.39 ± 0.61 | |
| c, y, α | b, y, α | d, y, α | cd, y, α | c, y, α | bc, y, α | d, y, α | b, y, α | ||
| P50 | LSA | 58.16 ± 0.96 | 55.24 ± 1.31 | 63.20 ± 2.00 | 64.29 ± 0.99 | 7.83 ± 0.17 | 8.17 ± 0.17 | 129.19 ± 3.11 | 127.70 ± 2.27 |
| c, x, α | c, x, α | d, x, α | c, x, α | c, x, α | b, x, α | d, x, α | c, x, α | ||
| SSA | 47.96 ± 1.41 | 47.80 ± 1.39 | 32.90 ± 0.77 | 31.75 ± 0.44 | 4.16 ± 0.13 | 5.97 ± 0.19 | 85.02 ± 2.15 | 85.51±1.21 | |
| c, y, α | ab, y, α | e, y, α | d, y, α | d, y, β | b, y, α | e, y, α | b, y, α | ||
| P100 | LSA | 55.61 ± 0.83 | 65.55 ± 0.49 | 66.03±0.69 | 76.08 ± 0.93 | 10.34 ± 0.28 | 8.58 ± 0.14 | 131.98 ± 0.37 | 150.21 ± 0.89 |
| d, x, β | ab, x, α | cd, x, β | a, x, α | ab, x, α | ab, x, β | cd, x, β | a, x, α | ||
| SSA | 46.50 ± 1.08 | 46.11 ± 0.37 | 46.24 ± 0.81 | 44.18 ± 1.04 | 4.90 ± 0.08 | 6.97 ± 0.15 | 97.64 ± 0.51 | 97.27 ± 1.32 | |
| c, y, α | b, y, α | c, y, α | a, y, α | c, y, β | a, y, α | c, y, α | a, y, α | ||
| P150 | LSA | 68.90 ± 1.62 | 67.80 ± 1.76 | 78.04 ± 0.78 | 72.34 ± 1.47 | 10.84 ± 0.09 | 8.90 ± 0.19 | 157.79 ± 2.43 | 149.04 ± 0.83 |
| a, x, α | a, x, α | a, x, α | ab, x, β | a, x, α | ab, x, β | a, x, α | a, x, β | ||
| SSA | 59.34±1.33 | 46.42 ± 1.55 | 57.92 ± 0.97 | 39.59 ± 0.80 | 7.31 ± 0.17 | 5.87 ± 0.13 | 124.57 ± 0.74 | 91.88 ± 1.84 | |
| b, y, α | b, y, β | b, y, α | b, y, β | b, y, α | b, y, β | b, y, α | ab, y, β | ||
| P200 | LSA | 65.89 ± 0.84 | 64.31 ± 0.91 | 74.50 ± 1.14 | 71.27 ± 0.95 | 10.01 ± 0.23 | 8.59 ± 0.27 | 150.39 ± 1.77 | 144.18 ± 1.85 |
| a, x, α | b, x, α | ab, x, α | ab, x, α | ab, x, α | ab, x, β | ab, x, α | ab, x, α | ||
| SSA | 65.61 ± 0.69 | 50.35 ± 1.27 | 64.15 ± 0.85 | 37.27 ± 1.00 | 7.94 ± 0.12 | 4.79 ± 0.24 | 137.70 ± 0.46 | 92.40 ± 2.29 | |
| a, x, α | a, y, β | a, y, α | bc, y, β | a, y, α | c, y, β | a, y, α | ab, y, β | ||
| P Level | Soils | Shoot P (mg P pot−1) | Root P (mg P pot−1) | Total P (mg P pot−1) | |||
|---|---|---|---|---|---|---|---|
| Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | ||
| P0 | LSA | 186.42 ± 2.49 | 216.76 ± 3.16 | 16.27 ± 1.27 | 19.89 ± 0.45 | 202.70 ± 2.44 | 236.66 ± 2.92 |
| b, x, β | ab, x, α | ab, x, α | ab, x, α | b, x, β | a, x, α | ||
| SSA | 108.02 ± 1.04 | 128.17 ± 3.64 | 18.61 ± 0.42 | 11.62 ± 0.45 | 126.63 ± 0.62 | 139.79 ± 3.21 | |
| d, y, β | ab, y, α | a, x, α | a, y, β | c, y, β | ab, y, α | ||
| P50 | LSA | 127.41 ± 1.85 | 182.60 ± 1.29 | 15.22 ± 0.90 | 23.81 ± 1.46 | 142.63 ± 1.62 | 206.41 ± 0.23 |
| c, x, β | c, x, α | ab, x, β | a, x, α | c, x, β | b, x, α | ||
| SSA | 128.48 ± 3.40 | 125.19 ± 2.35 | 16.09 ± 0.53 | 11.46 ± 0.65 | 144.56 ± 2.89 | 136.65 ± 2.96 | |
| c, x, α | ab, y, α | a, x, α | a, y, β | b, x, α | ab, y, α | ||
| P100 | LSA | 123.12 ± 1.88 | 206.02 ± 2.25 | 16.83 ± 0.84 | 20.78 ± 1.11 | 139.96 ± 1.96 | 226.80 ± 2.14 |
| c, x, β | b, x, α | ab, x, β | ab, x, α | c, x, β | a, x, α | ||
| SSA | 131.95 ± 3.21 | 115.36 ± 3.07 | 19.56 ± 1.53 | 9.69 ± 0.66 | 151.51 ± 4.72 | 125.05 ± 3.73 | |
| c, x, α | b, y, β | a, x, α | ab, y, β | b, x, α | b, y, β | ||
| P150 | LSA | 218.90 ± 6.61 | 219.59 ± 3.51 | 18.20 ± 0.20 | 18.61 ± 1.18 | 237.10 ± 6.75 | 238.20 ± 4.67 |
| a, x, α | a, x, α | a, x, α | b, x, α | a, x, α | a, x, α | ||
| SSA | 150.70 ± 0.81 | 133.31 ± 8.35 | 20.46 ± 1.37 | 10.97 ± 0.79 | 171.15 ± 1.91 | 144.29 ± 7.77 | |
| b, y, α | ab, y, α | a, x, α | a, y, β | a, y, α | ab, y, β | ||
| P200 | LSA | 177.82 ± 5.57 | 211.92 ± 2.28 | 14.09 ± 0.43 | 19.50 ± 0.58 | 191.92 ± 5.44 | 231.41 ± 2.87 |
| b, x, β | ab, x, α | b, x, β | ab, x, α | b, x, β | a, x, α | ||
| SSA | 165.17 ± 0.92 | 142.51 ± 1.21 | 16.81 ± 0.65 | 7.38 ± 0.14 | 181.98 ± 0.31 | 149.89 ± 1.25 | |
| a, x, α | a, y, β | a, y, α | b, y, β | a, x, α | a, y, β | ||
| P Level | Soils | Shoot P (mg P pot−1) | Grain P (mg P pot−1) | Root P (mg P pot−1) | Total P (mg P pot−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | Dongdao-4 | Tongyu-315 | ||
| P0 | LSA | 44.81 ± 1.63 | 45.69 ± 2.60 | 247.63 ± 5.33 | 151.90 ± 5.66 | 21.72 ± 1.95 | 33.79 ± 0.18 | 314.16 ± 8.86 | 231.38 ± 3.70 |
| b, x, α | b, x, α | ab, x, α | c, x, β | a, x, β | bc, x, α | abc, x, α | d, x, β | ||
| SSA | 52.93 ± 2.97 | 33.92 ± 1.38 | 131.24 ± 7.75 | 82.24 ± 0.46 | 48.08 ± 0.81 | 30.41 ± 0.82 | 232.25 ± 8.31 | 146.57 ± 2.19 | |
| ab, x, α | b, y, β | d, y, α | b, y, β | a, y, α | bc, y, β | bc, y, α | c, y, β | ||
| P50 | LSA | 47.29 ± 1.24 | 50.76 ± 2.26 | 220.24 ± 8.85 | 152.38 ± 3.83 | 22.90 ± 1.22 | 36.92 ± 1.37 | 290.43 ± 10.79 | 240.07 ± 0.63 |
| b, x, α | ab, x, α | b, x, α | c, x, β | a, x, β | ab, x, α | c, x, α | cd, x, β | ||
| SSA | 59.45 ± 1.40 | 40.70 ± 0.84 | 112.49 ± 2.59 | 77.86 ± 1.37 | 35.84 ± 2.04 | 35.19 ± 1.26 | 207.77 ± 5.50 | 153.76 ± 0.96 | |
| a, y, α | ab, y, β | d, y, α | b, y, β | bc, y, α | ab, x, α | c, y, α | bc, y, β | ||
| P100 | LSA | 42.84 ± 0.50 | 55.37 ± 1.61 | 231.08 ± 7.42 | 186.26 ± 1.92 | 19.78 ± 1.14 | 28.65 ± 0.58 | 293.70 ± 8.60 | 270.29 ± 4.09 |
| b, x, β | a, x, α | b, x, α | a, x, β | a, x, β | d, x, α | bc, x, α | a, x, α | ||
| SSA | 45.62 ± 2.95 | 35.56 ± 1.39 | 157.07 ± 0.41 | 101.83 ± 4.64 | 31.32 ± 1.46 | 39.05 ± 3.21 | 234.00 ± 4.42 | 176.44 ± 0.96 | |
| b, x, α | ab, y, β | c, y, α | a, y, β | c, y, α | a, y, α | b, y, α | a, y, β | ||
| P150 | LSA | 56.05 ± 2.37 | 54.57 ± 0.56 | 267.19 ± 3.81 | 169.70 ± 3.42 | 19.64 ± 1.59 | 38.49 ± 0.72 | 342.88 ± 6.69 | 262.76 ± 3.04 |
| a, x, α | a, x, α | a, x, α | ab, x, β | a, x, β | a, x, α | a, x, α | ab, x, β | ||
| SSA | 57.53 ± 0.97 | 38.44 ± 2.37 | 200.57 ± 4.79 | 97.71 ± 2.57 | 42.18 ± 0.84 | 28.52 ± 1.92 | 300.27 ± 4.45 | 164.67 ± 3.00 | |
| a, x, α | ab, y,β | b, y, α | a, y, β | ab, y, α | bc, y, β | a, y, α | ab, y, β | ||
| P200 | LSA | 50.03 ± 1.80 | 53.64 ± 1.44 | 261.86 ± 2.50 | 166.20 ± 2.11 | 17.22 ± 1.66 | 30.65 ± 1.12 | 329.11 ± 3.60 | 250.50 ± 4.13 |
| ab, x, α | ab, x, α | a, x, α | bc, x, β | a, x, β | cd, x, α | ab, x, α | bc, x, β | ||
| SSA | 54.15 ± 2.09 | 43.55 ± 3.03 | 222.77 ± 1.80 | 90.28 ± 3.12 | 24.08 ± 2.09 | 26.03 ± 0.30 | 301.00 ± 2.23 | 159.85 ± 4.16 | |
| ab, x, α | a, y, β | a, y, α | ab, y, β | d, x, α | c, y, α | a, y, α | b, y, β | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Li, J.; He, X.; Jia, X.; Yang, F.; Wang, Z. Grain Yield, Dry Weight and Phosphorus Accumulation and Translocation in Two Rice (Oryza sativa L.) Varieties as Affected by Salt-Alkali and Phosphorus. Sustainability 2017, 9, 1461. https://doi.org/10.3390/su9081461
Tian Z, Li J, He X, Jia X, Yang F, Wang Z. Grain Yield, Dry Weight and Phosphorus Accumulation and Translocation in Two Rice (Oryza sativa L.) Varieties as Affected by Salt-Alkali and Phosphorus. Sustainability. 2017; 9(8):1461. https://doi.org/10.3390/su9081461
Chicago/Turabian StyleTian, Zhijie, Jingpeng Li, Xinhua He, Xueying Jia, Fu Yang, and Zhichun Wang. 2017. "Grain Yield, Dry Weight and Phosphorus Accumulation and Translocation in Two Rice (Oryza sativa L.) Varieties as Affected by Salt-Alkali and Phosphorus" Sustainability 9, no. 8: 1461. https://doi.org/10.3390/su9081461
APA StyleTian, Z., Li, J., He, X., Jia, X., Yang, F., & Wang, Z. (2017). Grain Yield, Dry Weight and Phosphorus Accumulation and Translocation in Two Rice (Oryza sativa L.) Varieties as Affected by Salt-Alkali and Phosphorus. Sustainability, 9(8), 1461. https://doi.org/10.3390/su9081461