1. Introduction
Structure and biochemical constituents govern the optical responses of the vegetation [
1]. This fact allows remote sensing to provide spatial and temporal information on vegetation properties, which is necessary to understand key processes and variables such as photosynthesis or health status of the plants [
2,
3,
4]. Remote sensors provide spectro-directional reflectance factors (
RF) at the top of the canopy that can inform on both vegetation structure and leaf properties. Linking these variables is not trivial since the problem is ill-posed and other factors, such as woody elements, background scattering, illumination conditions, or the viewing geometry confound their relationship [
5,
6]. Radiative Transfer Models (RTM), spectral unmixing, and data fusion are the main ways to address these issues in remote sensing [
7,
8]. RTM are physical models capable of simulating the interaction of light with vegetation at leaf and canopy levels. These models are based on the radiative transfer equation that studies the propagation of radiation through an absorptive, dispersive, and/or emissive medium. The most frequently used RTM describes the vegetation canopy as a turbid medium where infinitesimally small leaves and gaps are distributed randomly [
9]. These models assume that leaves are bi-Lambertian [
10] and that Leaf Area Index (
LAI) and Leaf Angle Distribution (
LAD) mainly control the architecture of the canopy [
11]. This 1-D approach can acceptably represent homogeneous vegetation canopies like most of the crops, grasslands, and closed forests; but might not hold for heterogeneous and discontinuous vegetation canopies [
12,
13]. More complex RTM provide a 3-D representation of the vegetation that is more appropriate for these ecosystems [
7]. 3-D RTM overcome traditional approaches in radiative transfer based on the Kubelka–Munk theory [
14,
15,
16] and use more suitable methods to perform the spatial and spectral scaling [
7]. Among the most common approaches is the Monte Carlo Ray Tracing method that simulates a chain of dispersion events experienced by a photon in its trajectory from the source to the receiver or until its absorption [
17,
18]. Monte Carlo models allow accurate representation of the canopy, especially, when a large number of photons—usually over 10
6 photons—are processed, which results in large computational costs and processing times [
19]. In recent years, increasing computing capabilities have expanded the use of Monte-Carlo-based RTM to a number of applications [
20,
21,
22,
23]. For instance, Kötz et al. 2004 [
13] demonstrated the possibility of estimating live fuel moisture and green fuel loading in a coniferous forest with 3-D RTM FLIGHT [
17]. Simulated spectra compared to images of the airborne Reflective Optics System Imaging Spectrometer (ROSIS) and Hyperion Satellite Imaging Spectrometer obtained a relative deviation of around 5% in all wavelengths. Errors found were related to uncertainties in the variables describing the canopy structure such as
LAI or the fractional Vegetation Cover (
FCover).
3-D RTM usually involve a large number of parameters to describe, in detail, the vertical and horizontal distribution of the canopy, which might require field data to accurately simulate the directional reflectance of a given pixel. For example, Schneider et al. [
24] collected substantial field data in order to accurately simulate a temperate mixed forest with a Discrete Anisotropic Radiative Transfer (DART) model [
25,
26]. Such simulations were successfully compared with images acquired with the Airborne Prism Experiment (APEX) sensor. The study analyzed the sources of error and concluded that these were partly related to an inadequate representation of small-scale structures and the variability of leaf optical properties between species and individual trees. They also carried out an analysis of the background influence on simulated
RF by replacing a background based on the spectral properties of the main species in the study area by a black 100% absorbing background. Results showed an impact that was higher than the authors expected for a relatively dense mixed forest. The impact should be much higher in ecosystems with low tree fractional cover and high clumping, such as tree-grass ecosystems (TGE).
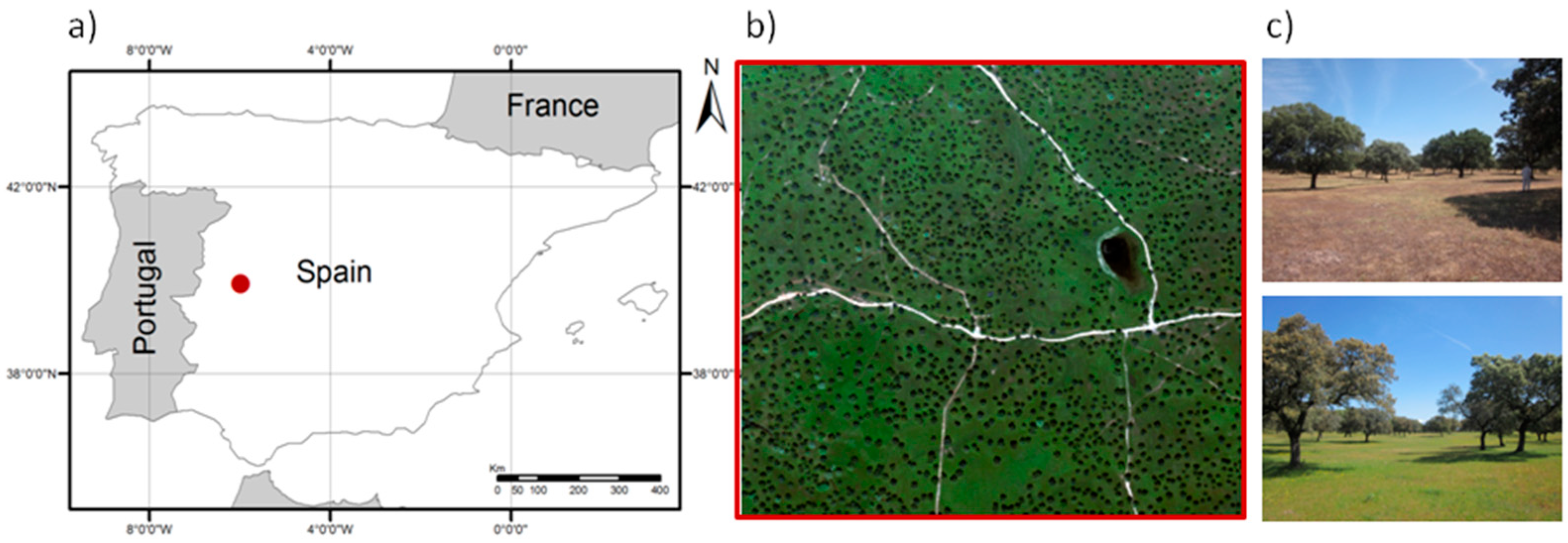
Mixed tree-grass and shrub-grass vegetation associations are one of the most spatially extensive and widely distributed forms of terrestrial vegetation on Earth and are often linked to livestock production [
27]. These ecosystems experience great pressure from changes in land use, at the same time that they are expected to increase in surface area and shift their geographical distributions as a consequence of climate change [
28]. The expected up-coming changes and increased role of TGE demand the improvement of our understanding of their functioning and our capability of monitoring their status from space.
TGE combine an overstory of evergreen or deciduous trees, sometimes a middle layer of shrubs, and a grass understory which—in some bioclimatic regions as in the Mediterranean—, can present high biodiversity as well as large spatial variability and strong temporal dynamics. Several approaches have been applied to simulate the optical properties of inhomogeneous ecosystems using physical or empirical methods [
25,
29,
30]. A number of 3-D RTM have been developed to model complex bi-layered ecosystems such as the Forest Radiative Transfer(FRT) model [
31] or the 5-scale model [
32]. These complex models are the result of an ensemble of RTM operating at different scales (leaf, canopy, different layers…). In addition, vegetation RTM are sometimes coupled to atmospheric RTM in order to simulate realistic illumination conditions [
31] or retrieve vegetation parameters from top of the atmosphere radiances observed from remote sensors [
33]. Several of these models have been tested in the third phase of the RAdiation Transfer Model Intercomparison (RAMI) project (
http://rami-benchmark.jrc.ec.europa.eu) [
34].
FLIGHT is one of the 3-D models tested in the third phase of the RAMI experiment [
34]. It presents some features that make it suitable to model vegetation in Mediterranean environments. On one side, FLIGHT is not directly coupled to a leaf RTM. The model demands leaf reflectance and transmittance factors, which can be provided externally by models or databases. This feature is convenient in those cases where RTM cannot accurately represent leaf optical properties of certain species since measurements can be directly provided. For example, in Mediterranean ecosystems, sclerophyllous species such as
Quercus ilex L. can develop different protective structures on leaf surfaces. Pacheco-Labrador et al., [
35] found that trichomes can strongly modify leaf optical properties, hampering the inversion of leaf RTM PROSPECT to estimate leaf biophysical properties. On the other hand, the FLIGHT model allows for the representation of two different types of leaves. This is suitable to represent canopies combining leaves of different ages or in different stages of development. For example, in Mediterranean ecosystems, evergreen species can present annual regrowth of new leaves which are very different from the older leaves remaining in the crown from previous years. A number of studies have used FLIGHT to simulate canopy reflectance in different ecosystems [
20,
21,
23]. Most of these works mainly focused on the representation of
RF as a function of tree crowns’ properties standing on bare soil. However, in TGE the background is also composed to vegetation and can represent high fractions of the surface. Therefore, an adequate representation of the radiative transfer of this layer is necessary to mechanistically link vegetation properties to the
RF in these ecosystems. This study aims to improve the capability of FLIGHT to simulate the spectral
RF of TGE with a grass understory by coupling this model with PROSAIL [
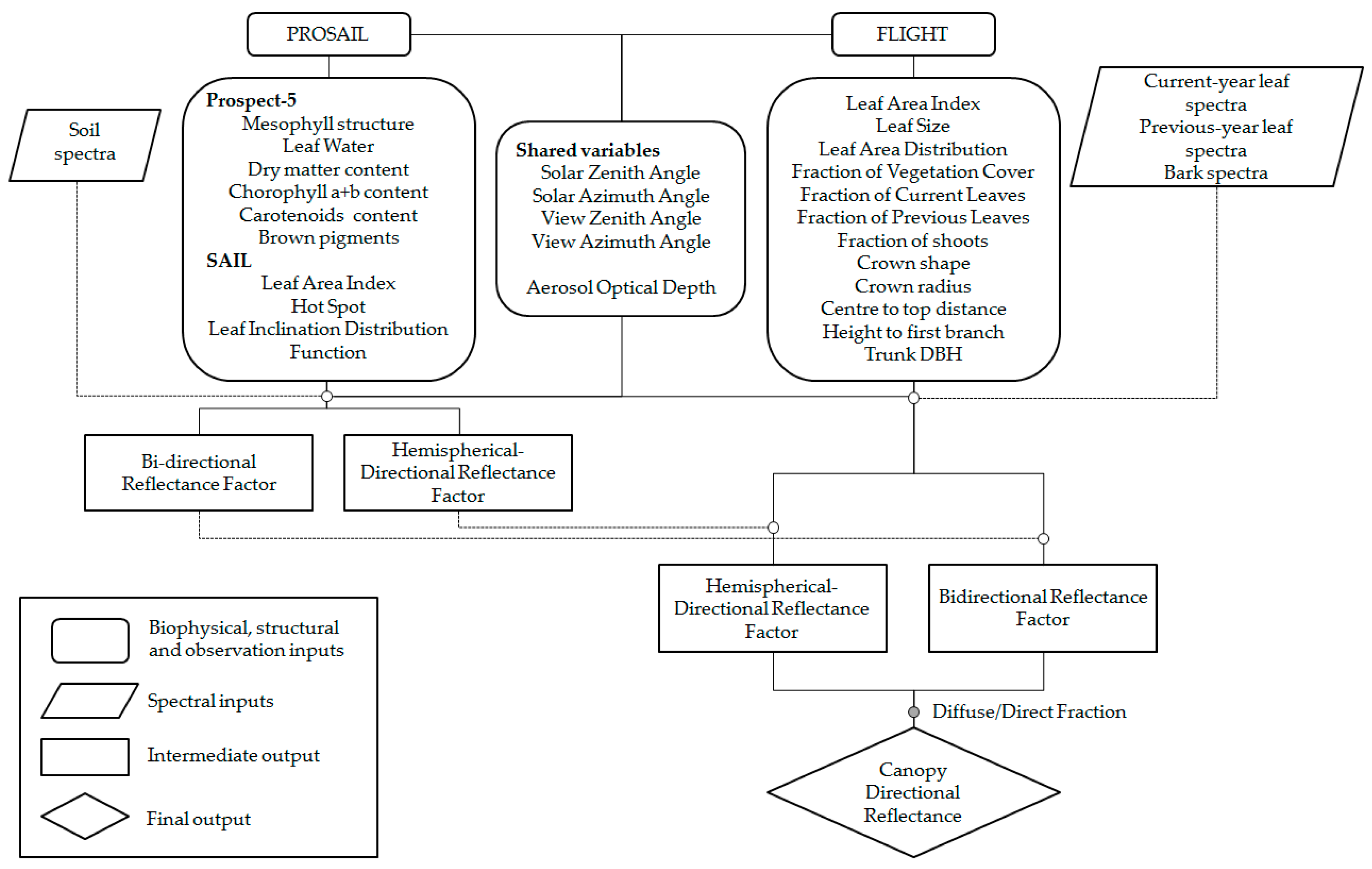
36] (hereafter PROSAIL+FLIGHT). In this model, the 3-D FLIGHT model represents tree crowns whereas the 1-D RTM PROSAIL features the underlying grassland.
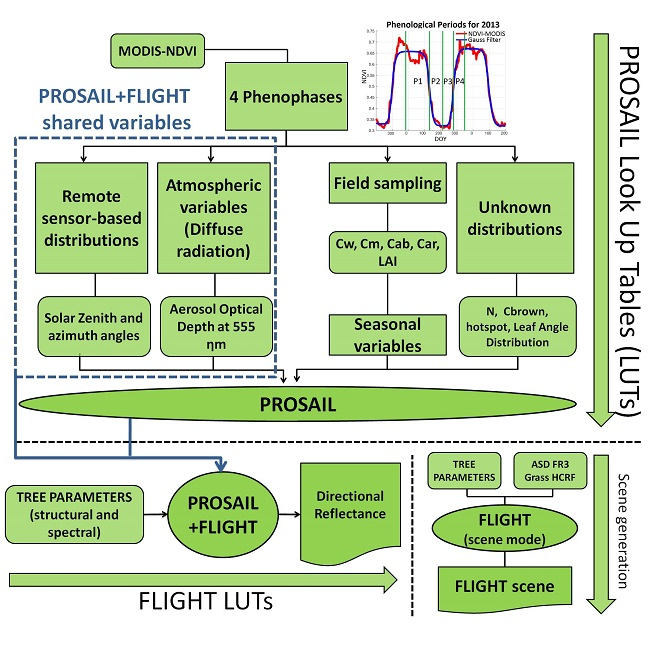
PROSAIL+FLIGHT is tested in a Mediterranean TGE against
RF measurements at proximal and remote sensing scales. First, we tested the capability of the model to represent the ranges and the seasonal variability of spectral
RF measured in the study area. To do so, the main phenological phases (hereafter, phenophases) were characterized using Moderate Resolution Imaging Spectroradiometer (MODIS) time series analysis in the study site (
Section 2.1). This allowed us to limit the ranges and define the covariance between the biophysical parameters of vegetation in each phenophase according to field measurements. We used this information to generate Look Up Tables (LUT) of
RF characteristics of each of these periods with PROSAIL and PROSAIL+FLIGHT; the RTM represented the herbaceous background and the ecosystem, respectively. Secondly, field biophysical data were used to simulate
RF with PROSAIL and PROSAIL+FLIGHT, which were compared with spectral measurements acquired in the same spots at different spatial scales (ground, airborne, and satellite). Finally, we compared
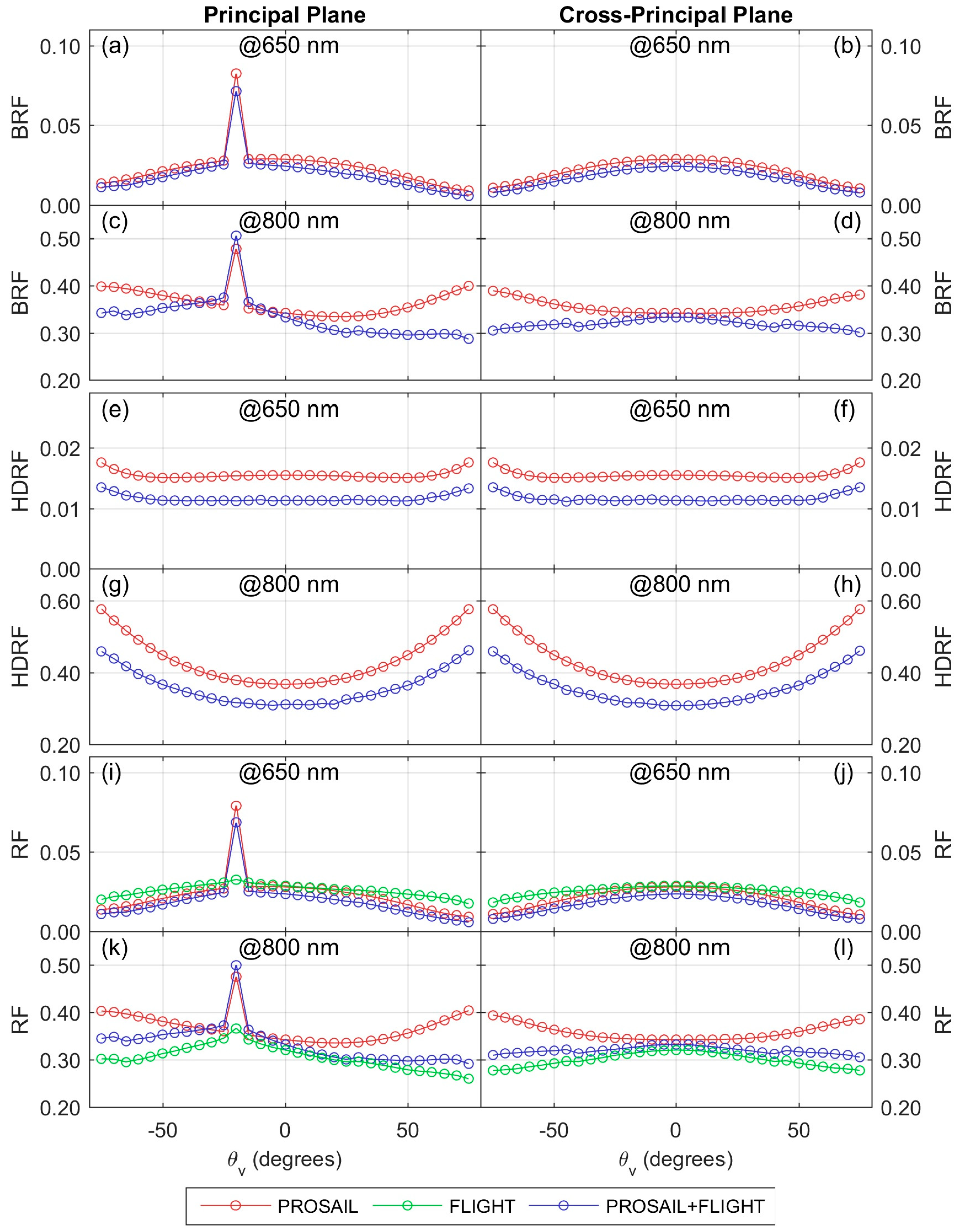
RF predicted by PROSAIL, FLIGHT, and PROSAIL+FLIGHT at different angles in the principal and the cross-principal plane.
4. Discussion
The combination of the 3-D RTM FLIGHT and the 1-D RTM PROSAIL allowed the simulation of RF representative of Mediterranean TGE as a function of the parameters of the two vegetation layers present in the ecosystem. We tested the performance of the PROSAIL and PROSAIL+FLIGHT models at grass and ecosystem scales respectively. Lower RRMSE found at close range can be, in part, explained by spatial scaling uncertainties. The comparison of both models on an ecosystem scale and at different angular configurations suggest that PROSAIL could mimic RF of the ecosystem when optical and/or biophysical properties of the grass and the tree layers are similar and observation and illumination angles are high. However, the predictive capability of the 1-D model would decrease when differences between layer properties or sun/view angles increase and the contribution of tree crowns and the 3-D structure of the ecosystem play a stronger roll on the signal.
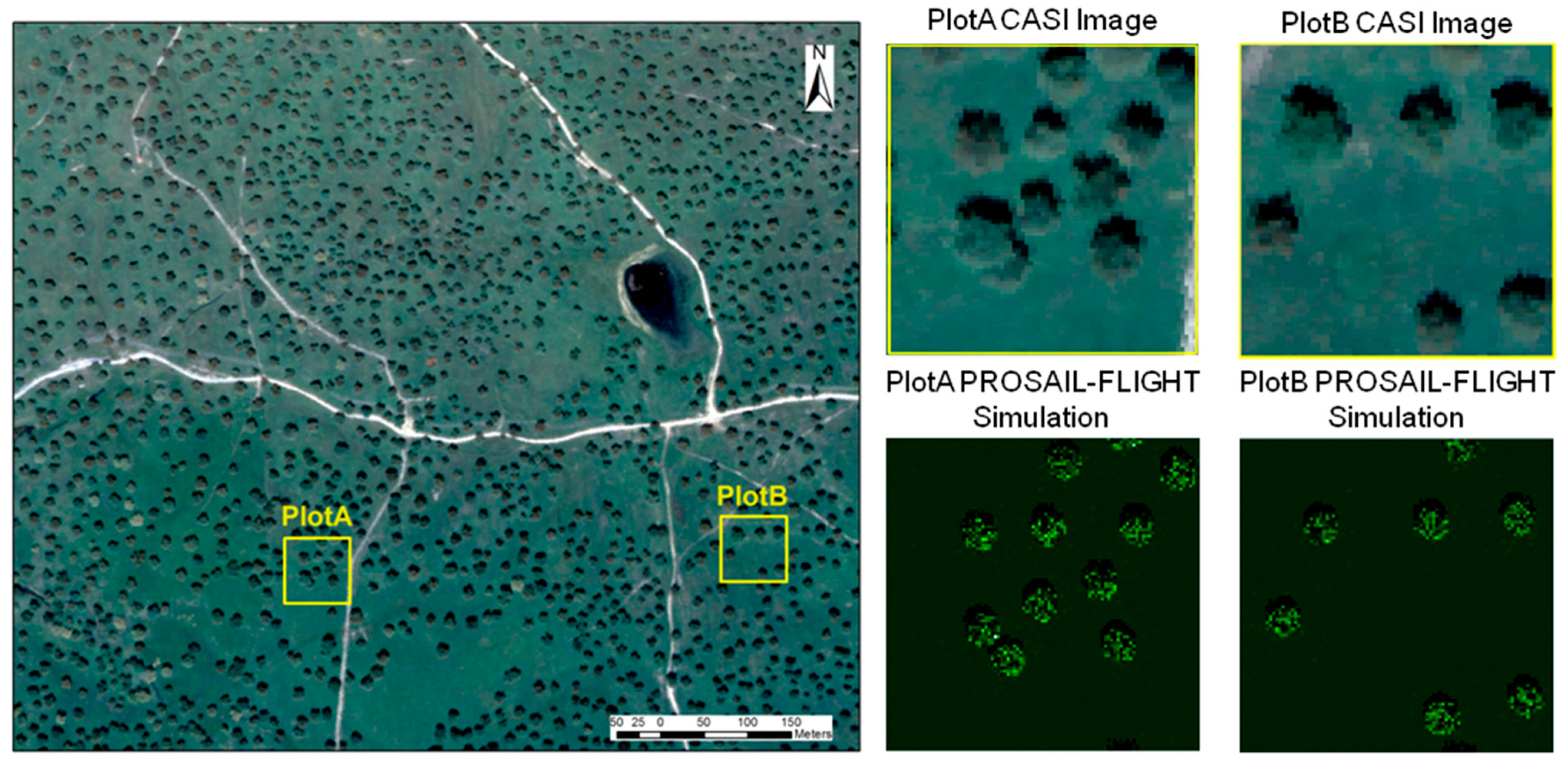
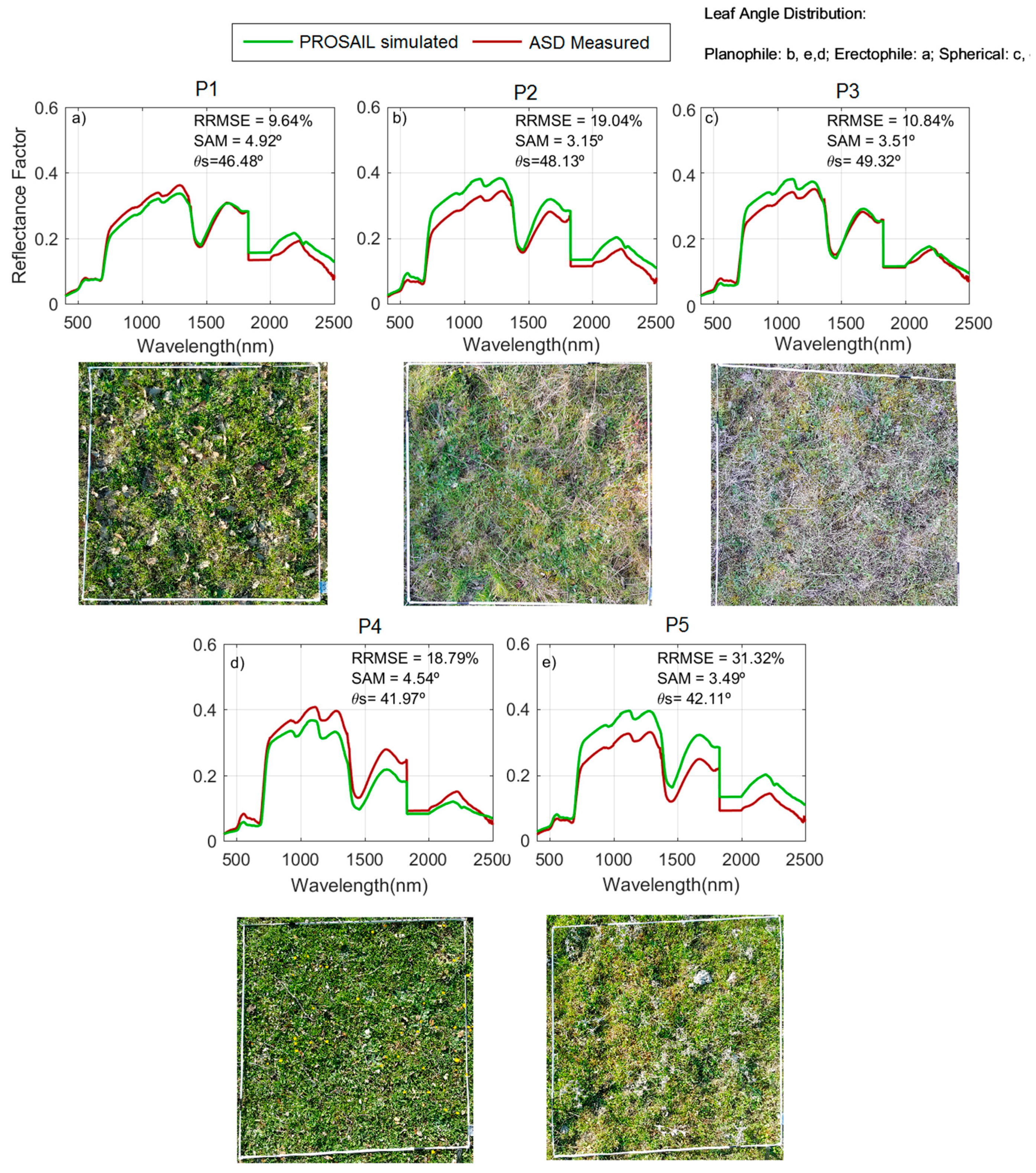
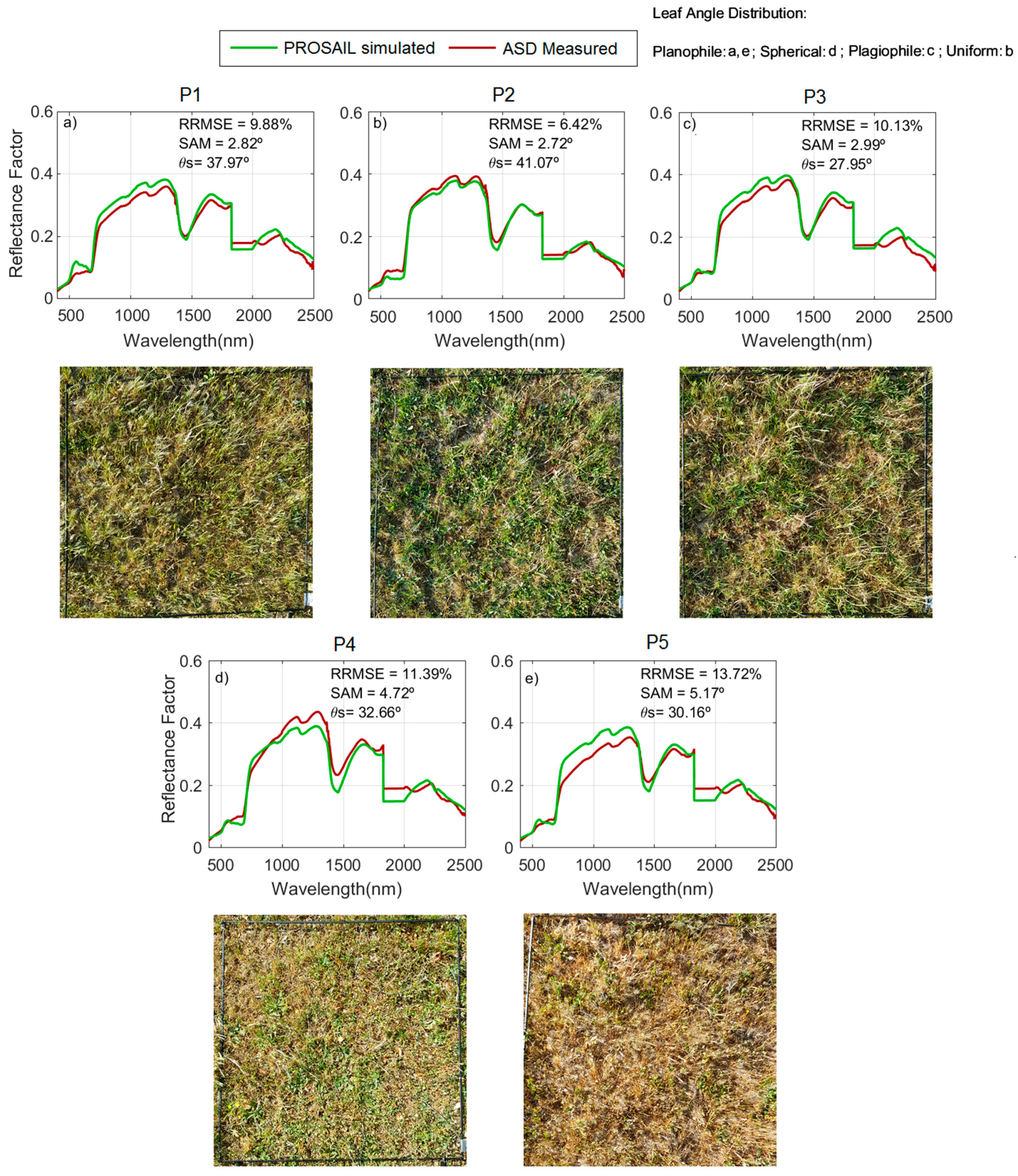
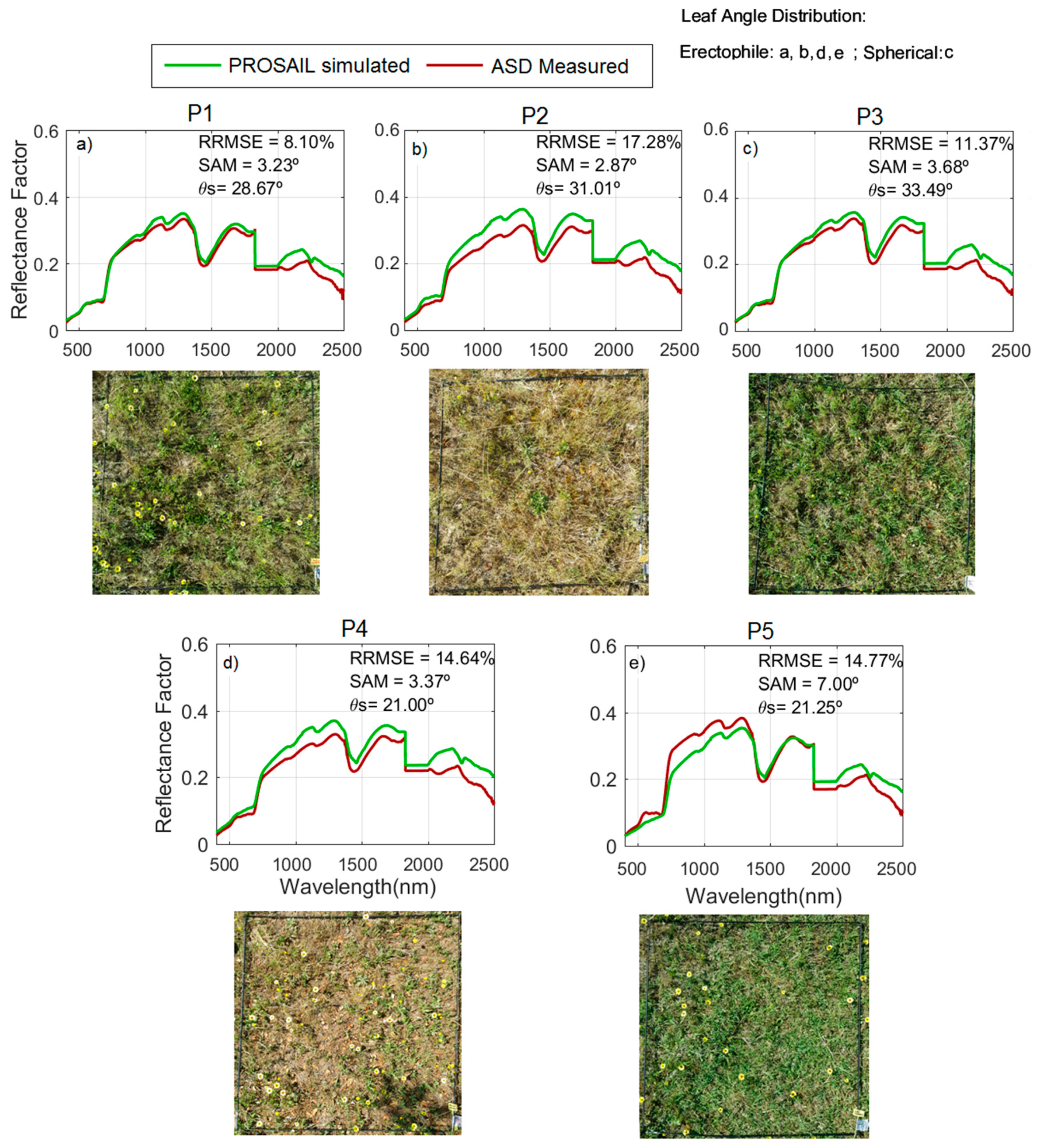
During the model evaluation, larger uncertainties were found at the ecosystem level than at the grass plot scale. These differences are, in part, explained by the larger spatial mismatch existing between the remote sensing spectral footprint and the small quadrants where biophysical-sampling-provided model inputs were taken within the 75 m × 75 m plots. The limited biophysical sampling might not be able to represent the properties of a large pixel in grassland characterized by a high spatial variability. However, in the simulations performed with Sentinel-2, see
Figure 12, the coupled PROSAIL+FLIGHT represented pixel
RF better than the PROSAIL simulation of the background, see
Figure 12b,c. Other sources of uncertainty that explain the differences between simulations and observations could be a) the uncertainties in the laboratory estimation of the biophysical variables, b) the selection of
AOT and other parameters not measured such as
LAD,
Cbrown, or the structural parameter
N, and c) inaccuracy of the models to represent vegetation radiative transfer of specific ecosystem components. In fact, our results suggest that current RTM do not properly represent the optical properties of standing death material, which induces large differences between predictions and observations when this is present. This jeopardizes the use of physical models to simulate
RF and estimate vegetation parameters in Mediterranean TGE and, in general, in grasslands which accumulate large amounts of grass litter over time. Also, PROSAIL+FLIGHT does not account for the change of angular distribution of photons falling onto the grass due to scattering and diffuse transmission within the tree crowns when
BRF is represented. Uncertainties induced by this model error would be larger in the NIR than in the VIS region, where crown transmittance and scattering are larger. At high observation and illumination angles, tree crowns will tend to occlude the same areas that are occluded to direct illumination, minimizing, in general, the observation of areas where the background
BRF is more overweighed by the model with respect to
HDRF.
PROSAIL+FLIGHT simplifies the representation of the background
BRDF without mapping a BRDF for each spectral band but rather separating the model into
BRF and
HDRF components. This approach speeds up computation but is still more accurate than assuming a Lambertian vegetation background. Errors induced by this simplification are expected to be comparable to other errors induced by model assumptions such as Lambertian symmetric leaves, omission of clumping, or the homogeneous distribution of leaves within the canopy, as well as the problems of the RTM regarding the accurate representation of vegetation optical properties ([
45], this article). Also, the simulation of the illumination conditions adopted by FLIGHT may become an oversimplification. The combination of vegetation and atmospheric radiative transfer models could improve the atmospheric correction and retrieval of vegetation parameters [
74]. Inaccuracies in the simulation of illumination conditions may account for part of the errors found in the different comparisons performed here with remote sensing data. The use of complex atmospheric RTM such as MODTRAN [
75,
76], or 6S [
77] could help to improve the quality of the simulations if their input parameters can be accurately quantified.
Field, airborne, and satellite data assessed the capability of the model to reproduce
RF measured at different scales. Due to the lack of observations of some of the input variables, see
Table 1, the comparison between simulated and measured spectra cannot be strictly considered a validation. However, it allowed us to understand which parts of the model might produce most of the uncertainties. PROSAIL and PROSAIL+FLIGHT simulations were separately compared with ground and airborne/satellite observations. This exercise revealed that most of the differences between simulated and observed
RF at the ecosystem scale were induced by the background RTM (PROSAIL), which failed to adequately represent the optical properties of death stand material. Another factor explains part of the uncertainties found in the comparison of models with remote sensing data; the high spatial variability of the grass layer hampers the use of a few samples to represent the average biophysical state of a remote sensing plot. This issue is inherent to grasslands with a high biodiversity and heterogeneous distribution of species in space and time, such as the ecosystem under study. Migliavacca et al., [
40] found that differences in
LAD related to nutrient availability could explain strong differences in the optical properties of the grassland layer in the same study site of this article. Former works accounted for 145 different species throughout the year in the study area [
44]. Such biodiversity makes unfeasible scaling through species-based metrics such as in [
78], and makes valuable use of modeling approaches that can be validated at a proximal sensing scale to later produce remote estimates of vegetation properties, as in this work.
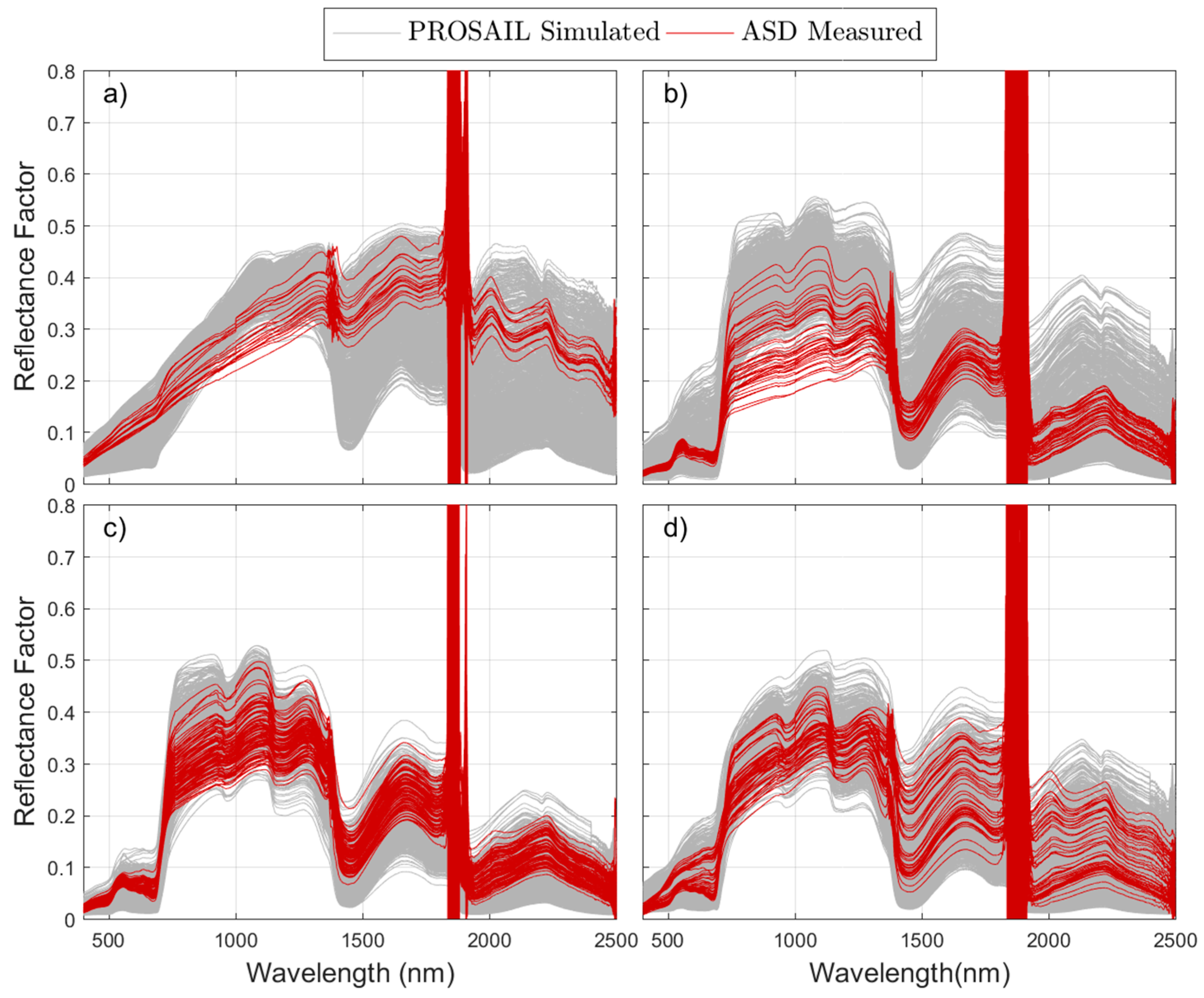
The comparison of field observations with the simulated
RF at a proximal sensing scale minimized spatial mismatch between spectral and biophysical footprints. This allowed us to detect the systematic overestimation of NIR
RF in phenophase 2: Autumn regrowth, as well as in phenophase 1: Summer drought, see
Figure 5a,b. We hypothesize that such an overestimation could be related to the presence of, not only senescent, but decomposing grass material, whose optical properties are not well represented by the PROSPECT model, see
Figure 14. This standing death material would feature a low reflectance in the NIR caused by a strong deterioration of the cell walls [
79] and the degradation of spongy mesophyll. This would translate into a change in the leaf refractive index in PROSPECT that has not been characterized. Such a model error would increase uncertainties in the estimation of biophysical variables both from RTM (this work) and empirical approaches [
80]. Future studies could focus on characterizing the optical properties of this decomposing litter, which would probably improve the simulation of grass
RF in phenophase 2 (autumn regrowth) at the beginning of the growth season. Despite these imbalances, it is noteworthy that the angular differences measured with SAM between the simulated and measured spectra of the grass layer do not generally exceed 5°; therefore, the shape of the spectra is similar but only differences in albedo are observed.
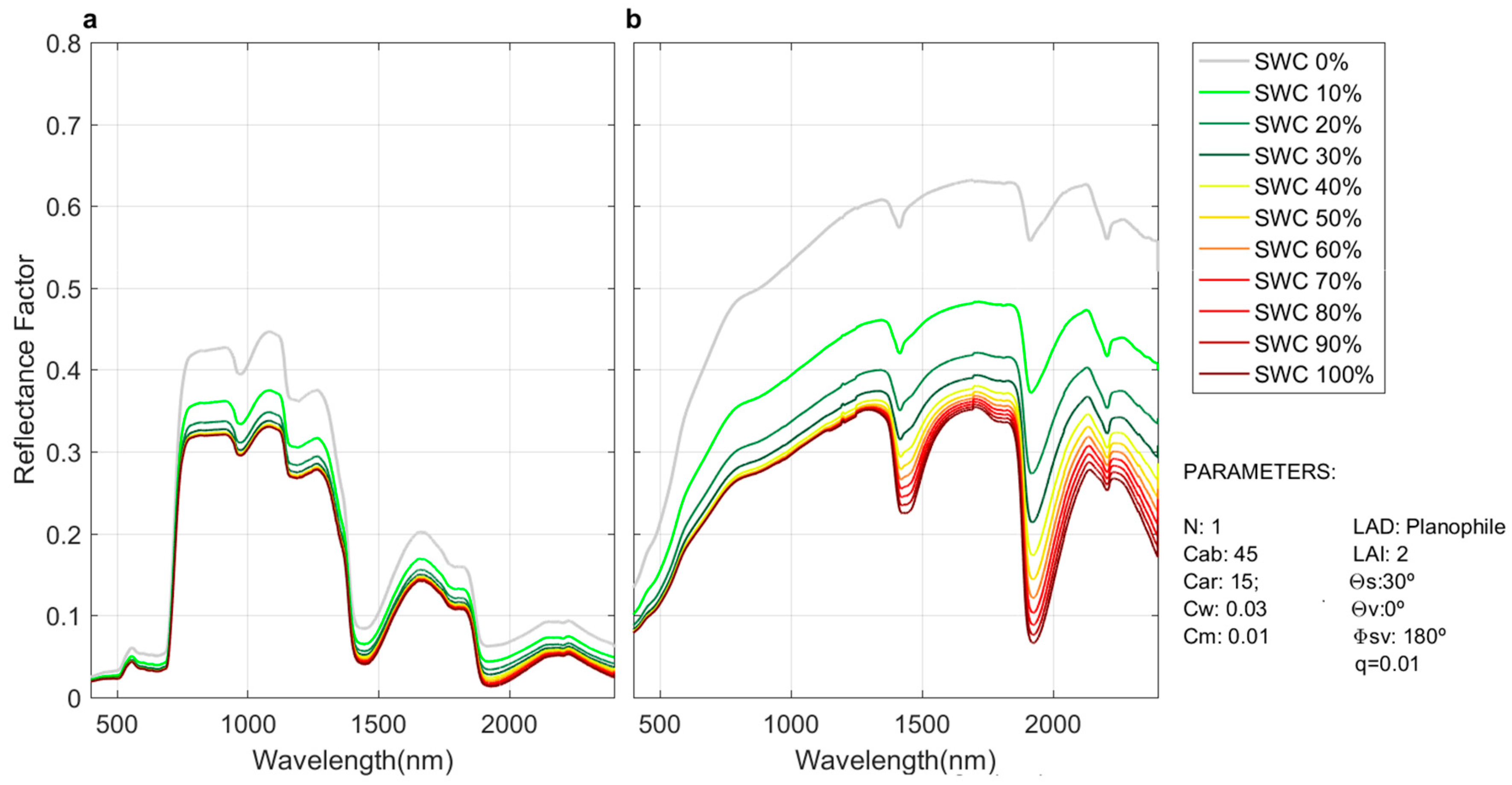
Uncertainties, in the SWIR and NIR regions, might also be related to errors in the representation of soil
RF. In this work, soil
RF was predicted as a function of
SWC measured at a depth of 5 cm at a few points located in the center of the study area.
SWC can greatly vary with meso- and micro-topography; therefore, a spatial heterogeneity in the ecosystem might not have been accurately simulated [
81,
82]. The soil influence on the observed signal depends on grass
LAD and
LAI [
83], in combination with observation and illumination angles.
Figure 15 shows simulated grass, see
Figure 15a and soil, see
Figure 15b,
RF observed at nadir for different
SWC values. As can be seen, the largest changes occur when
SWC is low (less than 20%), which can concur with
LAIgrass values between 0.1 and 2.2 during phenophase 1.
Regarding the temporal variability of the ecosystem and the effect on the modeling, phenophases that were defined according to the time series analysis over MODIS-
NDVI data were found to be appropriate to represent the phenology of the TGE under investigation. However, large overlaps between phenophases, especially between phenophase 2 and 3, reveals the strong inter-annual variability of this ecosystem. LUT design has been oriented so that each LUT is capable of representing
RF in each of the surrounding transition periods with no loss of accuracy. In addition, other sources of information could improve the phenological characterization of the ecosystem, such as phenocams [
41,
84] or spectral indices adapted to phenological analyses [
85]. Enhanced information would contribute to improving the definition of the phenophases and the classification of field remote sensing data in each of them.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}














