The Response of Vegetation Phenology and Productivity to Drought in Semi-Arid Regions of Northern China

Abstract
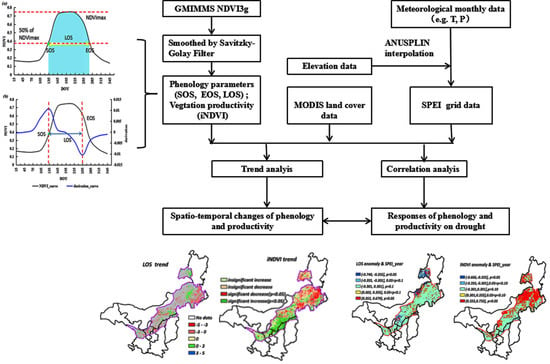
:
1. Introduction
2. Data and Methods
2.1. Study Area
2.2. Data Source and Pre-Processing
2.2.1. GIMMS NDVI 3g Dataset
2.2.2. Drought Index and Meteorological Data
2.2.3. Other Auxiliary Data
2.3. Extraction of Phenological Metrics and Vegetation Productivity
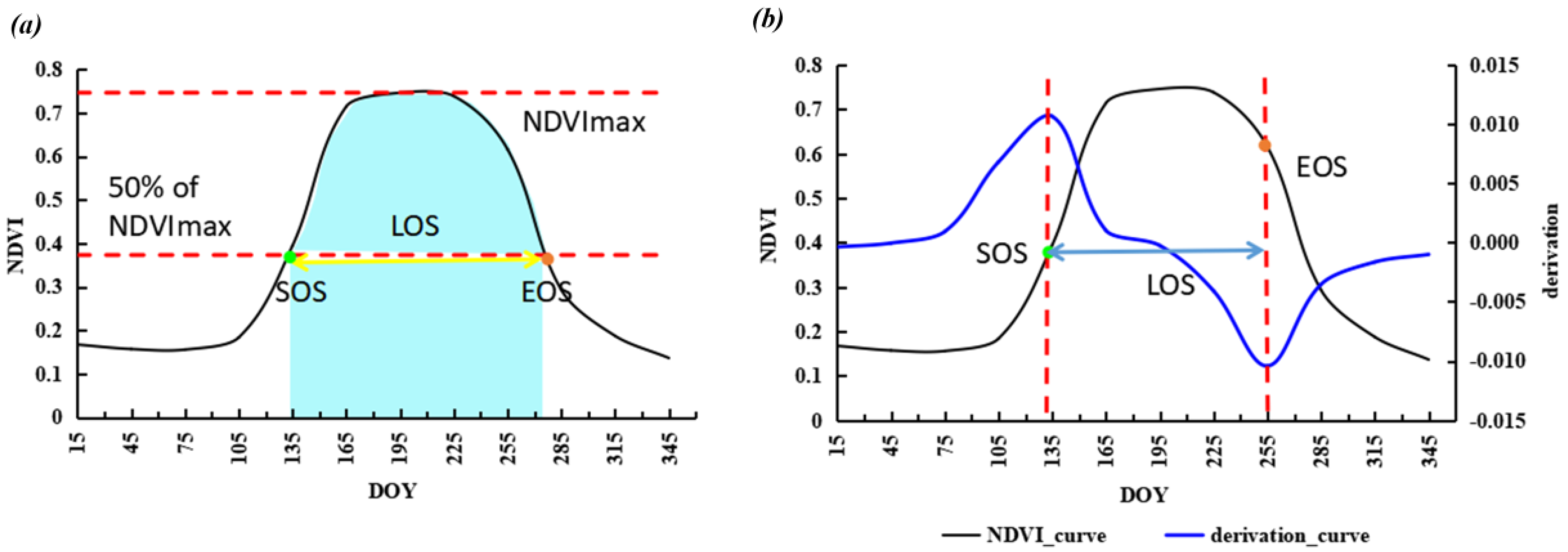
2.3.1. Dynamic Threshold Method (m1)
2.3.2. Derivative Method Based on the Fitted Double Logistic Function (m2)
2.4. Statistics
2.4.1. Trend Analysis
2.4.2. Correlation Analysis
3. Results
3.1. Climatic Variation Analysis in Semi-Arid Regions of Northern China
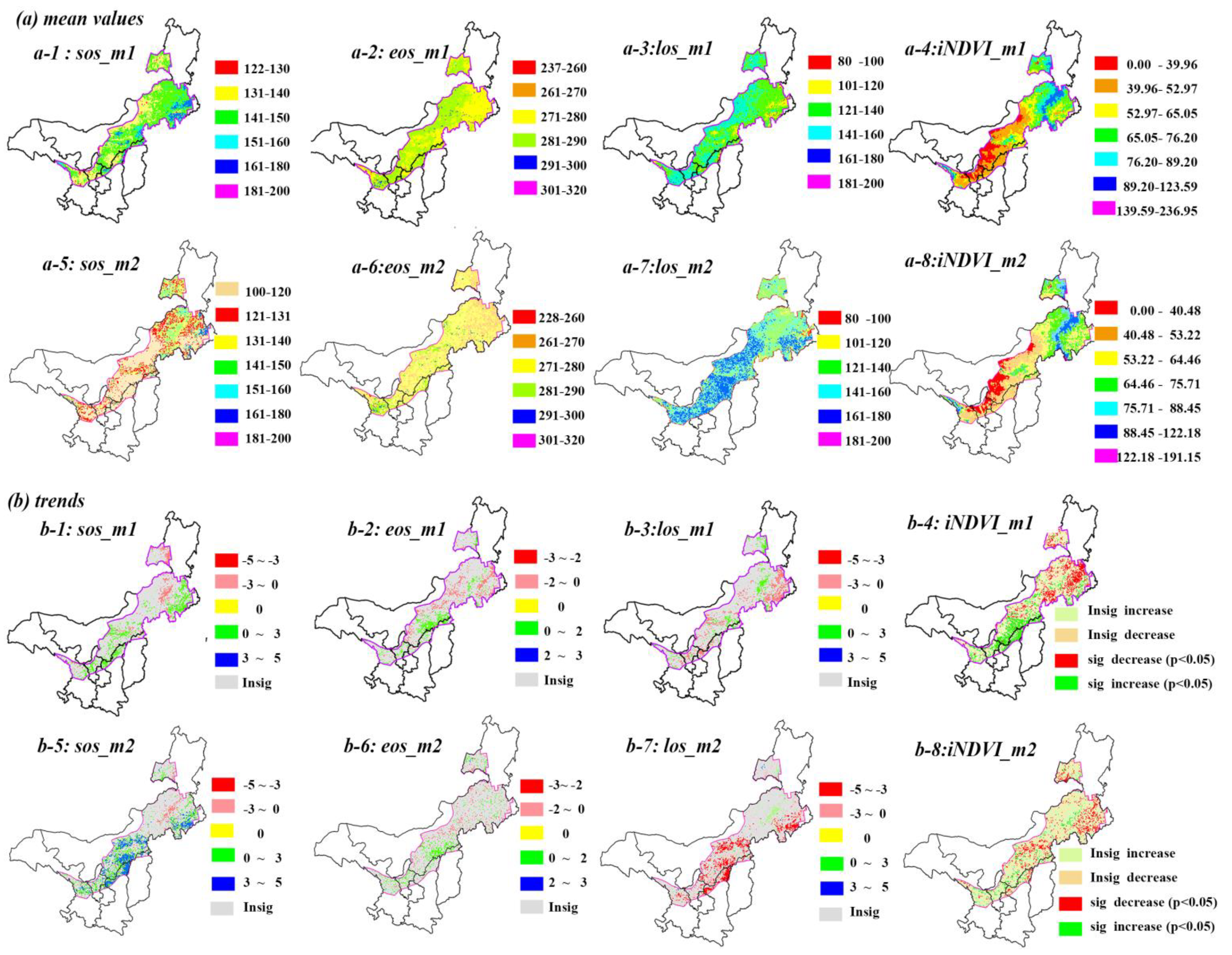
3.2. Spatial-Temporal Patterns of Vegetation Phenology and Productivity
3.3. Response of Phenology and Productivity to Drought
3.4. The Phenology and Productivity among Different Land Cover Types
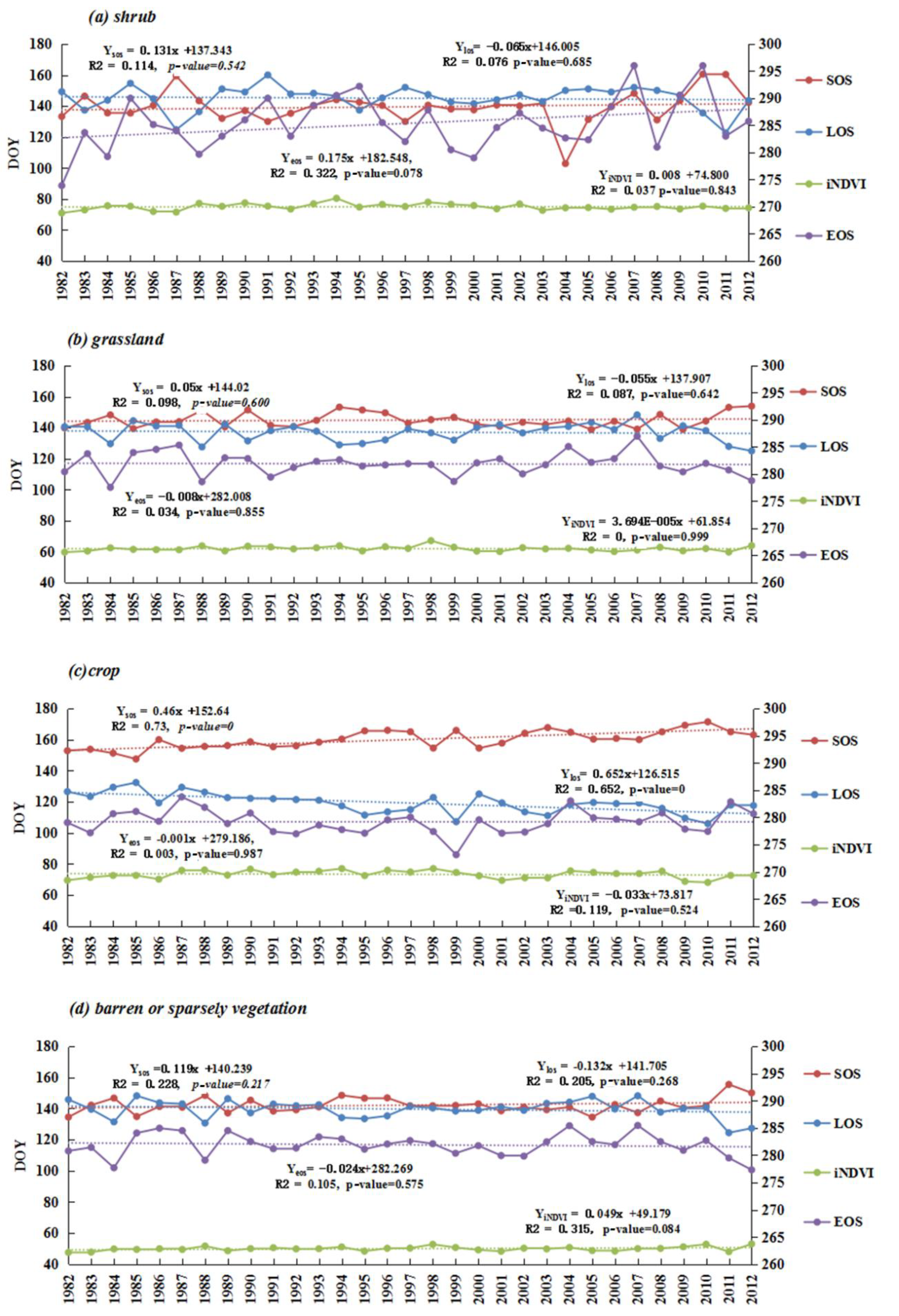
3.4.1. The Spatio-Temporal Trends of Phenology among Different Land Cover Types
3.4.2. The Response of Phenology and Productivity among Different Land Cover Types
4. Discussions
4.1. Spatio-Temporal Trends of Vegetation Phenology and Productivity
4.2. The Responses of Vegetation Phenology and Productivity to Drought
4.3. Analysis at Different Land Cover Types
4.4. Uncertainty Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Poulter, B.; Pederson, N.; Liu, H.; Zhu, Z.; D’Arrigo, R.; Ciais, P.; Davi, N.; Frank, D.; Leland, C.; Myneni, R. Recent trends in inner Asian forest dynamics to temperature and precipitation indicate high sensitivity to climate change. Agric. For. Meteorol. 2013, 178, 31–45. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.; Ju, W.; Wang, S.; Wu, X.; He, M.; Zhu, G. Impacts of droughts on carbon sequestration by China’s terrestrial ecosystems from 2000 to 2011. Biogeosciences 2014, 11, 2583. [Google Scholar] [CrossRef]
- Hu, S.; Mo, X.; Lin, Z. Projections of spatial-temporal variation of drought in north China. Arid Land Geogr. 2015, 38, 239–248. [Google Scholar]
- Huang, J.; Zhai, J.; Jiang, T.; Wang, Y.; Li, X.; Wang, R.; Xiong, M.; Su, B.; Fischer, T. Analysis of future drought characteristics in China using the regional climate model CCLM. Clim. Dyn. 2017, 50, 1–19. [Google Scholar] [CrossRef]
- Stocker, T. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Ni, J. Plant functional types and climate along a precipitation gradient in temperate grasslands, North-East China and south-east Mongolia. J. Arid Environ. 2003, 53, 501–516. [Google Scholar] [CrossRef]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- D’Odorico, P.; Bhattachan, A.; Davis, K.F.; Ravi, S.; Runyan, C.W. Global desertification: Drivers and feedbacks. Adv. Water Resour. 2013, 51, 326–344. [Google Scholar] [CrossRef]
- Huang, L.; He, B.; Chen, A.; Wang, H.; Liu, J.; Lu, A.; Chen, Z. Drought dominates the interannual variability in global terrestrial net primary production by controlling semi-arid ecosystems. Sci. Rep. 2016, 6, 24639. [Google Scholar] [CrossRef] [PubMed]
- Ivits, E.; Horion, S.; Fensholt, R.; Cherlet, M. Drought footprint on european ecosystems between 1999 and 2010 assessed by remotely sensed vegetation phenology and productivity. Glob. Chang. Biol. 2014, 20, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yu, G.; Fan, J.; Wen, X. Effects of drought on ecosystem carbon and water processes: A review at different scales. Prog. Geogr. 2006, 25, 12–20. [Google Scholar]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Huete, A.; Moran, S.; Ponce Campos, G.; Eamus, D. Abrupt shifts in phenology and vegetation productivity under climate extremes. J. Geophys. Res. Biogeosci. 2015, 120, 2036–2052. [Google Scholar] [CrossRef]
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.; Liu, J. Changes in satellite-derived vegetation growth trend in temperate and Boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Zhou, Y.Z.; Jia, G.S. Precipitation as a control of vegetation phenology for temperate steppes in China. Atmos. Ocean. Sci. Lett. 2016, 9, 162–168. [Google Scholar] [CrossRef]
- Wu, C.; Hou, X.; Peng, D.; Gonsamo, A.; Xu, S. Land surface phenology of China’s temperate ecosystems over 1999–2013: Spatial–temporal patterns, interaction effects, covariation with climate and implications for productivity. Agric. For. Meteorol. 2016, 216, 177–187. [Google Scholar] [CrossRef]
- Wu, X.; Liu, H. Consistent shifts in spring vegetation green-up date across temperate biomes in China, 1982–2006. Glob. Chang. Biol. 2013, 19, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Kawamura, K.; Ishikawa, N.; Goto, M.; Wulan, T.; Alateng, D.; Yin, T.; Ito, Y. Modis normalized difference vegetation index (NDVI) and vegetation phenology dynamics in the inner Mongolia grassland. Solid Earth 2015, 6, 1185–1194. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Piao, S.; Ciais, P.; Li, J.; Friedlingstein, P.; Koven, C.; Chen, A. Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. Proc. Natl. Acad. Sci. USA 2011, 108, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fu, Y.H.; Zeng, Z.; Huang, M.; Li, X.; Piao, S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob. Chang. Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, G.; Li, Z.; Ye, X.; Wang, M.; Gong, L. Impacts of climate change on net primary productivity in arid and semiarid regions of China. Chin. Geogr. Sci. 2015, 26, 35–47. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Zhang, X.; Tarpley, D.; Sullivan, J.T. Diverse responses of vegetation phenology to a warming climate. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef]
- Wang, T.; Song, X.; Yan, C.; Li, S.; Xie, J. Remote sensing analysis on aeolian desertification trends in northern China during 1975–2010. J. Desert Res. 2011, 31, 1351–1356. [Google Scholar]
- Glade, F.E.; Miranda, M.D.; Meza, F.J.; van Leeuwen, W.J. Productivity and phenological responses of natural vegetation to present and future inter-annual climate variability across semi-arid river basins in chile. Environ. Monit. Assess. 2016, 188, 676. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Yokozawa, M.; Zhang, Z.; Hayashi, Y.; Ishigooka, Y. Land surface phenology dynamics and climate variations in the north east China transect (NECT), 1982–2000. Int. J. Remote Sens. 2008, 29, 5461–5478. [Google Scholar] [CrossRef]
- Cui, T.; Martz, L.; Guo, X. Grassland phenology response to drought in the Canadian prairies. Remote Sens. 2017, 9, 1258. [Google Scholar] [CrossRef]
- United Nations Environment Programme. World Atlas of Desertification; UNEP: Nairobi, Kenya, 1992. [Google Scholar]
- Miao, L.; Ye, P.; He, B.; Chen, L.; Cui, X. Future climate impact on the desertification in the dry land Asia using AVHRR gimms NDVI3g data. Remote Sens. 2015, 7, 3863–3877. [Google Scholar] [CrossRef]
- Pinzon, J.; Tucker, C. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Gimms NDVI3g Data. Available online: https://ecocast.arc.nasa.gov/data/pub/gimms/3g.v0/ (accessed on 5 May 2018).
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality ndvi time-series data set based on the savitzky–golay filter. Remote Sens. Environ. 2014, 91, 332–344. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- China Meteorological Data Sharing Service System. Available online: http://cdc.cma.gov.cn/ (accessed on 5 May 2018).
- Liu, S.; Kang, W.; Wang, T. Drought variability in inner mongolia of northern China during 1960–2013 based on standardized precipitation evapotranspiration index. Environ. Earth Sci. 2016, 75, 145. [Google Scholar] [CrossRef]
- Speicalc. Available online: http://digital.csic.es/handle/10261/10002 (accessed on 5 May 2018).
- Yu, M.; Li, Q.; Hayes, M.J.; Svoboda, M.D.; Heim, R.R. Are droughts becoming more frequent or severe in China based on the standardized precipitation evapotranspiration index: 1951–2010? Int. J. Climatol. 2014, 34, 545–558. [Google Scholar] [CrossRef]
- Hutchinson, M.F.; Xu, T. Anusplin Version 4.2 User Guide; The Australian National University: Canberra, Australia, 2004. [Google Scholar]
- Liu, Z.; Li, L.; McVicar, T.; Van Niel, T.; Yang, Q.; Li, R. Introduction of the professional interpolation software for meteorology data: Anusplin. Meteorol. Mon. 2008, 34, 92–100. [Google Scholar]
- Qian, Y.; Lv, H.; Zhang, Y. Application and assessment of spatial interpolation method on daily meteorological elements based on anusplin software. J. Meteorol. Environ. 2010, 26, 7–15. [Google Scholar]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. Modis collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- China Western Environment and Ecology Science Data Center. Available online: http://westdc.westgis.ac.cn (accessed on 5 May 2018).
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Hou, X.; Niu, Z.; Gao, S.; Huang, N. Monitoring vegetation phenology in farming-pastoral zone using SPOT-VGT NDVI data. Trans. Chin. Soc. Agric. Eng. 2013, 29, 142–150. [Google Scholar]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yokozawa, M.; Toritani, H.; Shibayama, M.; Ishitsuka, N.; Ohno, H. A crop phenology detection method using time-series modis data. Remote Sens. Environ. 2005, 96, 366–374. [Google Scholar] [CrossRef]
- Fan, D.; Zhao, X.; Zhu, W.; Zheng, Z. Review of influencing factors of accuracy of plant phenology monitoring based on remote sensing data. Prog. Geogr. 2016, 35, 304–319. [Google Scholar]
- Beck, P.S.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using modis NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Yu, L.; Liu, T.; Bu, K.; Yan, F.; Yang, J.; Chang, L.; Zhang, S. Monitoring the long term vegetation phenology change in northeast China from 1982 to 2015. Sci. Rep. 2017, 7, 14770. [Google Scholar] [CrossRef] [PubMed]
- Sen, P.K. Estimates of the regression coefficient based on kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Theil, H. A rank-invariant method of linear and polynomial regression analysis. In Henri Theil’s Contributions to Economics and Econometrics; Springer: Berlin, Germany, 1992; pp. 345–381. [Google Scholar]
- Fensholt, R.; Rasmussen, K.; Kaspersen, P.; Huber, S.; Horion, S.; Swinnen, E. Assessing land degradation/recovery in the African sahel from long-term earth observation based primary productivity and precipitation relationships. Remote Sens. 2013, 5, 664–686. [Google Scholar] [CrossRef] [Green Version]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Chang. Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, H.; Zhang, Z.; Guo, X.; Li, X.; Chen, C. Spatial and temporal changes in vegetation phenology at middle and high latitudes of the northern hemisphere over the past three decades. Remote Sens. 2015, 7, 10973–10995. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, H.; Shen, M.; Liang, W.; Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob. Chang. Biol. 2015, 21, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xu, J.; Chen, Y.; Li, W.; Yang, Y.; Hong, Y.; Li, Z. Understanding the dynamic coupling between vegetation cover and climatic factors in a semiarid region-a case study of inner Mongolia, China. Ecohydrology 2013, 6, 917–926. [Google Scholar] [CrossRef]
- Hinckley, T.; Dougherty, P.; Lassoie, J.; Roberts, J.; Teskey, R. A severe drought: Impact on tree growth, phenology, net photosynthetic rate and water relations. Am. Midl. Nat. 1979, 102, 307–316. [Google Scholar] [CrossRef]
- Chapin, F.S.; Maston, P.; Mooney, H.A. Principles of Terrestrial Ecosystem; Higher Education Press: Beijing, China, 2005; pp. 168–183. [Google Scholar]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fu, Y.H.; Zhu, Z.; Liu, Y.; Liu, Z.; Huang, M.; Janssens, I.A.; Piao, S. Delayed autumn phenology in the northern hemisphere is related to change in both climate and spring phenology. Glob. Chang. Biol. 2016, 22, 3702–3711. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.F.; Richardson, A.D. The timing of autumn senescence is affected by the timing of spring phenology: Implications for predictive models. Glob. Chang. Biol. 2015, 21, 2634–2641. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, X.; Li, Y.; Wang, T.; Shen, M.; Du, M.; He, H.; Li, Y.; Luo, W.; Ma, M. Spatiotemporal pattern of gross primary productivity and its covariation with climate in China over the last thirty years. Glob. Chang. Biol. 2017, 24, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the european continent from NDVI3g data and modis time series. Remote Sens. 2013, 6, 257–284. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of earth observation based global long term vegetation trends—Comparing gimms and modis global NDVI time series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Fensholt, R.; Nielsen, T.T.; Stisen, S. Evaluation of AVHRR pal and gimms 10-day composite NDVI time series products using spot-4 vegetation data for the African continent. Int. J. Remote Sens. 2006, 27, 2719–2733. [Google Scholar] [CrossRef]
- Schucknecht, A.; Erasmi, S.; Niemeyer, I.; Matschullat, J. Assessing vegetation variability and trends in north-eastern Brazil using AVHRR and modis NDVI time series. Eur. J. Remote Sens. 2013, 46, 40–59. [Google Scholar] [CrossRef]
- Fensholt, R.; Rasmussen, K.; Nielsen, T.T.; Mbow, C. Evaluation of earth observation based long term vegetation trends—Intercomparing NDVI time series trend analysis consistency of sahel from avhrr gimms, terra modis and spot vgt data. Remote Sens. Environ. 2009, 113, 1886–1898. [Google Scholar] [CrossRef]
- Mao, D.; Wang, Z.; Luo, L.; Ren, C. Integrating AVHRR and modis data to monitor NDVI changes and their relationships with climatic parameters in northeast China. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 528–536. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variables | Independent Variables | Sig Negative (p < 0.05) | Sig Positive (p < 0.05) | Total (p < 0.05) | |||
|---|---|---|---|---|---|---|---|
| m1 | m2 | m1 | m2 | m1 | m2 | ||
| SOS anomaly | SPEI-May | 8.28% | 11.04% | 2.00% | 4.48% | 10.28% | 15.52% |
| EOS anomaly | SPEI-Oct | 0.27% | 1.19% | 18.16% | 7.03% | 18.43% | 8.22% |
| LOS anomaly | SPEI_year | 10.45% | 8.07% | 4.78% | 3.92% | 15.23% | 11.99% |
| iNDVI anomaly | SPEI_year | 0.03% | 0.44% | 37.85% | 7.53% | 37.88% | 7.97% |
| Grassland | Shrub | Barren/Sparsely Veg | Cropland | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Dependent Variables | Independent Variables | r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| SOS anomaly | SPEI-May | −0.04 | 0.85 | 0.14 | 0.44 | −0.16 | 0.50 | −0.05 | 0.78 | |
| EOS anomaly | SPEI-Oct | 0.19 * | 0.03 | 0.14 | 0.27 | 0.31 * | 0.06 | 0.25 | 0.18 | |
| LOS anomaly | SPEI_year | 0.41 * | 0.02 | 0.19 | 0.32 | 0.28 | 0.12 | 0.27 | 0.14 | |
| iNDVI anomaly | SPEI_year | 0.54 * | 0.00 | 0.34 * | 0.05 | 0.36 * | 0.04 | 0.32 | 0.07 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, W.; Wang, T.; Liu, S. The Response of Vegetation Phenology and Productivity to Drought in Semi-Arid Regions of Northern China. Remote Sens. 2018, 10, 727. https://doi.org/10.3390/rs10050727
Kang W, Wang T, Liu S. The Response of Vegetation Phenology and Productivity to Drought in Semi-Arid Regions of Northern China. Remote Sensing. 2018; 10(5):727. https://doi.org/10.3390/rs10050727
Chicago/Turabian StyleKang, Wenping, Tao Wang, and Shulin Liu. 2018. "The Response of Vegetation Phenology and Productivity to Drought in Semi-Arid Regions of Northern China" Remote Sensing 10, no. 5: 727. https://doi.org/10.3390/rs10050727
APA StyleKang, W., Wang, T., & Liu, S. (2018). The Response of Vegetation Phenology and Productivity to Drought in Semi-Arid Regions of Northern China. Remote Sensing, 10(5), 727. https://doi.org/10.3390/rs10050727