Ocean Color Quality Control Masks Contain the High Phytoplankton Fraction of Coastal Ocean Observations


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Selection:
2.2. Atmospheric Dataset:
2.3. Biological Field Data
2.4. Satellite Data
2.5. Remote Estimation of Phytoplankton Biomass:
2.6. Match-up Procedure, Derivation of Climatological Averages and Error Statistics
3. Results
3.1. Association Between Red Band Difference and Phytoplankton Biomarkers
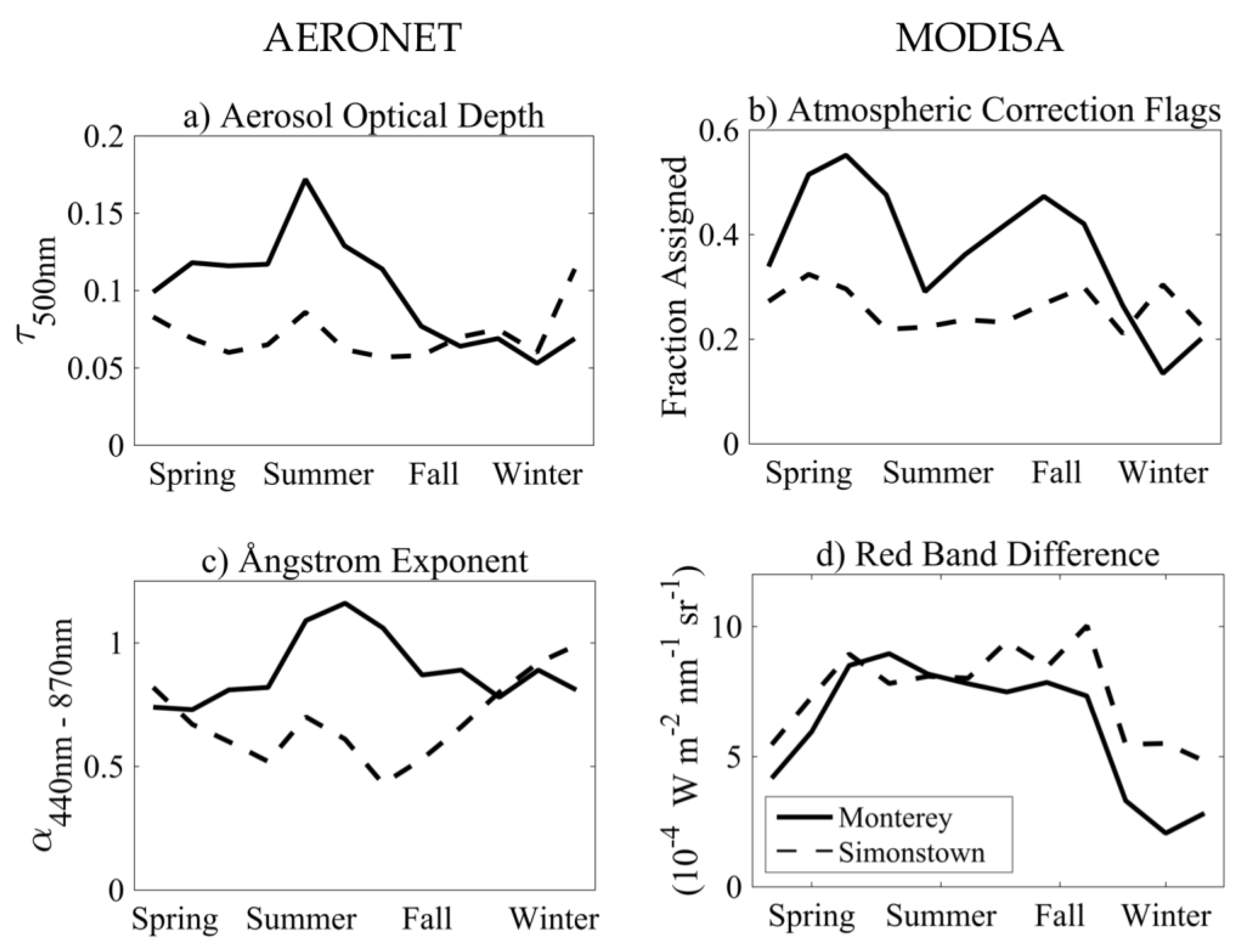
3.2. Climatology of Atmospheric Correction Flags
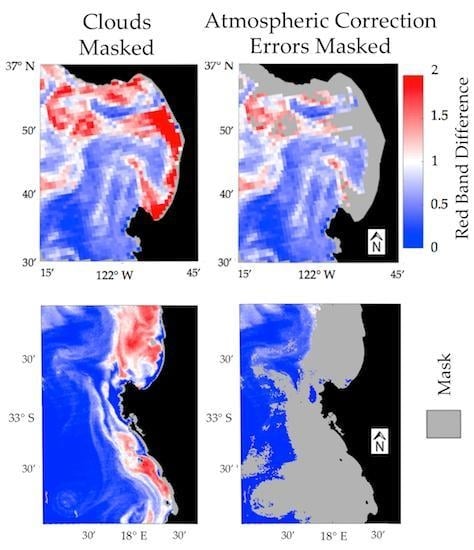
3.3. Impact of Atmospheric Correction Masks on Level 2 RBD datasets
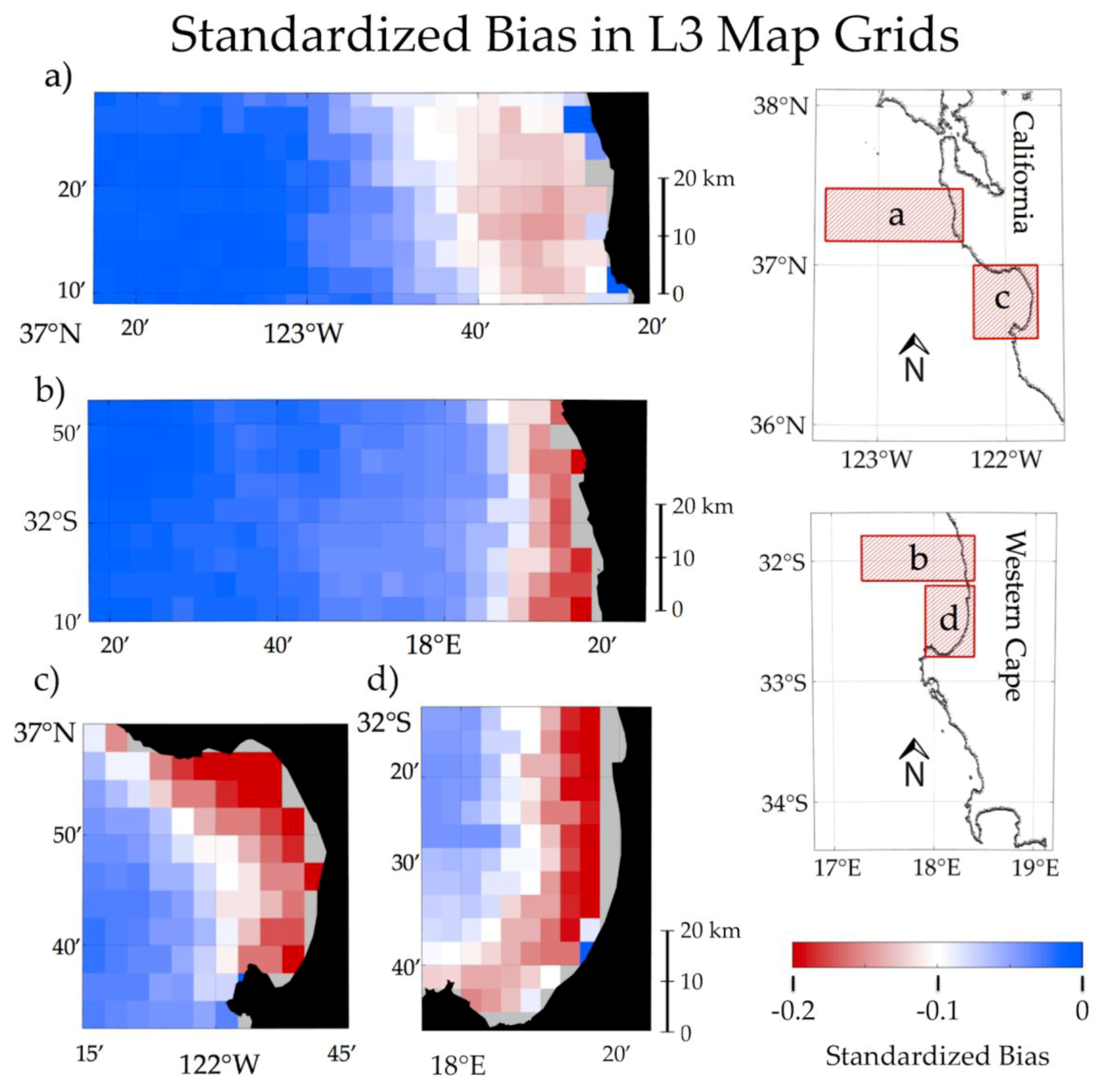
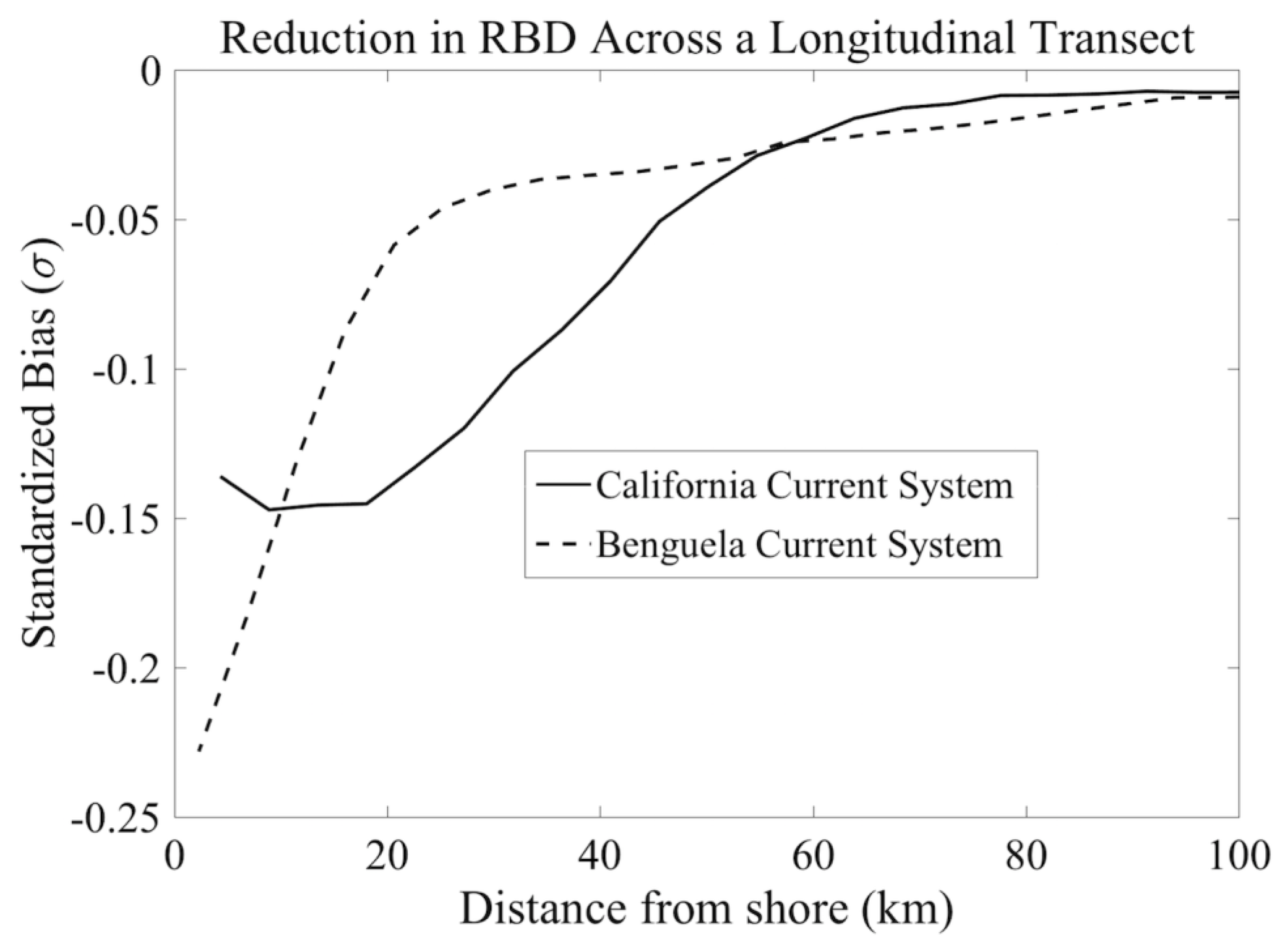
3.4. Impact of Atmospheric Correction Masks on Level 3 RBD Datasets
4. Discussion
4.1. Performance of Satellite Products at Match-up Sites
4.2. Variability of Atmospheric Constituents and Efficacy of Ocean Color AC Flags
4.3. Potential for User Evaluation of L2 and L3 Datasets
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gregg, W.W.; Conkright, M.E. Decadal changes in global ocean chlorophyll. Geophys. Res. Lett. 2002, 29, 1–4. [Google Scholar] [CrossRef]
- McClain, C.R. A Decade of Satellite Ocean Color Observations. Annu. Rev. Mar. Sci. 2009, 1, 19–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierssen, H.M. Perspectives on empirical approaches for ocean color remote sensing of chlorophyll in a changing climate. Proc. Natl. Acad. Sci. USA 2010, 107, 17073–17078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, D.A.; Wang, M.; Maritorena, S.; Robinson, W. Atmospheric correction of satellite imagery: The black pixel assumption. Appl. Opt. 2000, 39, 3582–3591. [Google Scholar] [CrossRef] [PubMed]
- Sathyendranath, S.; Bukata, R.P.; Arnone, R.; Dowell, M.D.; Davis, C.O.; Babin, M.; Berthon, J.F.; Kopelevich, J.; Cmpbell, J.W. Color of Case 2 Waters. In Remote Sensing of Ocean Colour in Coastal and Other Optically-Complex, Waters; Sathyendranath, S., Ed.; Reports of the International Ocean-Color Coordinating Group: Dartmouth, NS, Canada, 2010; Volume 3, pp. 23–46. [Google Scholar]
- Kahru, M.; Mitchell, B.G. Ocean color reveals increased blooms in various parts of the world. EOS Trans. Am. Geophys. Union 2008, 89, 170–172. [Google Scholar] [CrossRef]
- Jessup, D.A.; Miller, M.A.; Ryan, J.P.; Nevins, H.M.; Kerkering, H.A.; Mekebri, A.; Crane, D.B.; Johnson, T.A.; Kudela, R.M. Mass stranding of marine birds caused by a surfactant-producing red tide. PLoS ONE 2009, 4, e4550. [Google Scholar] [CrossRef]
- Lewitus, A.J.; Horner, R.A.; Carn, D.A.; Garcia-Mendoza, E.; Hickey, B.M.; Hunter, M.; Huppert, D.D.; Kudela, R.M.; Langlois, G.W.; Largier, J.L.; et al. Harmful algal blooms along the North American west coast region: History, trends, causes and impacts. Harmful Algae 2012, 19, 133–159. [Google Scholar] [CrossRef]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Figueiras, F.G.; Kudela, R.M.; Moita, T.; Reguera, B.; Ruiz-Villareal, M. Key questions and recent research advances on harmful algal blooms in eastern boundary upwelling systems. In Global Ecology and Oceanography of Harmful Algal Blooms, 1st ed.; Gilbert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer: Cham, Switzerland, 2018; Volume 232, pp. 205–227. [Google Scholar]
- Bracher, A.; Bouman, H.A.; Brewin, R.J.; Bricaud, A.; Brotas, V.; Ciotti, A.M.; Clementson, L.; Devred, E.; Di Cicco, A.; Dutkiewicz, S.; et al. Obtaining phytoplankton diversity from ocean color: A scientific roadmap for future development. Front. Mar. Sci. 2017, 4, 1–15. [Google Scholar] [CrossRef]
- Frouin, R.J.; Franz, B.A.; Ibrahim, A.; Knobelspiesse, K.; Ahmad, Z.; Cairns, B.; Chowdhary, J.; Dierssen, H.M.; Tan, J.; Dubovik, O.; et al. Atmospheric correction of satellite ocean-color imagery during the PACE era. Front. Earth Sci. 2019, 7, 1–43. [Google Scholar] [CrossRef]
- Barnes, R.A.; Clark, D.K.; Esaias, W.E.; Fargion, G.S.; Feldman, C.R.; McClain, C.R. Development of a consistent multi-sensor global ocean colour time series. Int. J. Remote Sens. 2003, 24, 4047–4064. [Google Scholar] [CrossRef]
- Gregg, W.W.; Casey, N.W. Improving the consistency of ocean color data: A step toward climate data records. Geophys. Res. Lett. 2010, 37, 1–5. [Google Scholar] [CrossRef]
- Kahru, M.; Kudela, R.M.; Manzano-Sarabia, M.; Mitchell, B.G. Trends in the surface chlorophyll of the California Current: Merging data from multiple ocean color satellites. Deep Sea Res. Part II Top. Stud. Oceanogr. 2012, 77, 89–98. [Google Scholar] [CrossRef]
- Gordon, H.R.; Wang, M. Retrieval of water-leaving radiance and aerosol optical thickness over the oceans with SeaWiFS: A preliminary algorithm. Appl. Opt. 1994, 33, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Mobley, C.D.; Werdell, J.; Franz, B.; Ahmad, Z.; Bailey, S. Atmospheric correction for satellite ocean color radiometry. NASA Tech. Rep. Serv. 2016, 1, 1–85. [Google Scholar]
- Wang, M.; Antoine, D.; Fruoin, R.; Gordon, H.R.; Fukushima, H.; Morel, A.; Nicolas, J.; Deschamps, P. Comparison Results. In Atmospheric Correction for Remotely-Sensed Ocean-Colour Products; Wang, A., Ed.; Reports of the International Ocean-Color Coordinating Group: Dartmouth, NS, Canada, 2010; Volume 10, pp. 23–38. [Google Scholar]
- Wang, M. Remote sensing of the ocean contributions from ultraviolet to near-infrared using the shortwave infrared bands: Simulations. Appl. Opt. 2007, 46, 1535–1547. [Google Scholar] [CrossRef] [PubMed]
- Ruddick, K.G.; Ovidio, F.; Rijkeboer, M. Atmospheric correction of SeaWiFS imagery for turbid coastal and inland waters. Appl. Opt. 2000, 39, 897–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockmann, C.; Doerffer, R.; Peters, M.; Stelzer, K.; Embacher, S.; Ruescas, A. Evolution of the C2RCC neural network for Sentinel 2 and 3 for the retrieval of ocean colour products in normal and extreme optically complex waters. In Proceedings of the Living Planet Symposium, Prague, Czech Republic, 9–13 May 2016; Volume 740, p. 54. [Google Scholar]
- Mograne, M.A.; Jamet, C.; Loisel, H.; Vantrepotte, V.; Mériaux, X.; Cauvin, A. Evaluation of five atmospheric correction algorithms over French optically-complex waters for the Sentinel-3A OLCI ocean color sensor. Remote Sens. 2019, 11, 668. [Google Scholar] [CrossRef]
- Campbell, J.W.; Blaisdell, J.M.; Darzi, M. Level-3 Sea WiFS data products: Spatial and temporal binning algorithms. Oceano. Lit. Rev. 1996, 9, 952. [Google Scholar]
- Scott, J.P.; Werdell, P.J. Comparing level-2 and level-3 satellite ocean color retrieval validation methodologies. Accepted Optics Express. 2019. [Google Scholar] [CrossRef]
- Ryan, J.P.; Fischer, A.M.; Kudela, R.M.; Gower, J.F.R.; King, S.A.; Marin III, R.; Chavez, F.P. Influences of upwelling and downwelling winds on red tide bloom dynamics in Monterey Bay, California. Cont. Shelf Res. 2009, 29, 785–795. [Google Scholar] [CrossRef]
- Pennington, J.T.; Chavez, F.P. Seasonal fluctuations of temperature, salinity, nitrate, chlorophyll and primary production at station H3/M1 over 1989-1996 in Monterey Bay, California. Deep Sea Res. Part II 2016, 47, 947–973. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Brown, P.C.; Mitchell-Innes, B.A. Spatio-temporal variability of phytoplankton in the southern Benguela upwelling system. S. Afr. J. Mar. Sci. 1992, 12, 439–456. [Google Scholar] [CrossRef]
- Barlow, R.; Sessions, H.; Balarin, M.; Weeks, S.; Whittle, C.; Hutchings, L. Seasonal variation in phytoplankton in the southern Benguela: Pigment indices and ocean colour. Afr. J. Mar. Sci. 2005, 27, 275–287. [Google Scholar] [CrossRef]
- Fawcett, A.; Pitcher, G.C.; Bernard, S.; Cembella, A.D.; Kudela, R.M. Contrasting wind patterns and toxigenic phytoplankton in the southern Benguela upwelling system. Mar. Ecol. Prog. Ser. 2007, 348, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Schumann, E.H.; Martin, J.A. Climatological aspects of the coastal wind field at Cape Town, Port Elizabeth and Durban. S. Afr. Geogr. J. 1991, 73, 48–51. [Google Scholar] [CrossRef]
- Patt, F.S.; Barnes, R.A.; Eplee, R.E., Jr.; Franz, B.A.; Robinson, W.D.; Feldman, G.C.; Bailey, S.W.; Gales, J.; Werdell, P.J.; Wang, M.; et al. Algorithm updates for the fourth SeaWiFS Data Reprocessing. In SeaWiFS Postlaunch Technical Report Series; Hooker, S.B., Firestone, E.R., Eds.; NASA Technical Reports; NASA: Greenbelt, MD, USA, 2002; Volume 22, pp. 34–40. [Google Scholar]
- Neville, R.A.; Gower, J.F.R. Passive remote sensing of phytoplankton via chlorophyll a fluorescence. J. Geophys. Res. 1977, 82, 3487–3493. [Google Scholar] [CrossRef]
- Gordon, H.R. Diffuse reflectance of the ocean: The theory of its augmentation by chlorophyll a fluorescence at 685 nm. Appl. Opt. 1979, 18, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Letelier, R.M.; Abbott, M.R. An analysis of chlorophyll fluorescence algorithms for the Moderate Resolution Imaging Spectrometer (MODIS). Remote Sens. Environ. 1996, 58, 215–223. [Google Scholar] [CrossRef]
- Gower, J.F.R. On the use of satellite-measured chlorophyll fluorescence for monitoring coastal waters. Int. J. Remote Sens. 2016, 37, 2077–2086. [Google Scholar] [CrossRef]
- Amin, R.; Zhou, J.; Gilerson, A.; Gross, B.; Moshary, F.; Ahmed, S. Novel optical techniques for detecting and classifying toxic dinoflagellate Karenia brevis blooms using satellite imagery. Opt. Express 2009, 17, 9126–9144. [Google Scholar] [CrossRef] [PubMed]
- Roesler, C.S.; Perry, M.J. In situ phytoplankton absorption, fluorescence emission and particulate backscattering spectra determined from reflectance. J. Geophys. Res. 1995, 100, 13279–13294. [Google Scholar] [CrossRef]
- Gilerson, A.; Zhou, J.; Hlaing, S.; Ioannou, I.; Schalles, J.; Gross, B.; Moshary, F.; Ahmed, S. Fluorescence component in the reflectance spectra from coastal waters. Dependence on water composition. Opt. Express 2007, 15, 15702–15721. [Google Scholar] [CrossRef] [PubMed]
- Gower, J.F.R.; Borstad, G.A. On the potential of MODIS and MERIS for imaging chlorophyll fluorescence from space. Int. J. Remote Sens. 2004, 25, 1459–1464. [Google Scholar] [CrossRef]
- Ryan, J.P.; Davis, C.O.; Tufillaro, N.B.; Kudela, R.M.; Gao, B.C. Application of the Hyperspectral Imager for the Coastal Ocean to phytoplankton ecology studies in Monterey Bay, CA, USA. Remote Sens. 2014, 6, 1007–1025. [Google Scholar] [CrossRef]
- Ryan, J.P.; Gower, J.F.R.; King, S.A.; Bissett, W.P.; Fischer, A.M.; Kudela, R.M.; Kolber, Z.; Mazzillo, F.; Rienecker, E.V.; Chavez, F.P. A coastal ocean extreme bloom incubator. Geophys. Res. Lett. 2008, 35, 1–5. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Product | |||
|---|---|---|---|---|
| Monterey Bay, California M1 Buoy | Red Band Difference Blue-Green Chla Algorithm1 Neural Network Chla Algorithm2 | 1012 773 840 | 0.4190 0.3344 0.2179 | 14.4% 14.8% 17.0% |
| Monterey Bay, California Santa Cruz Wharf | Red Band Difference Blue-Green Chla Algorithm1 Neural Network Chla Algorithm2 | 361 8 132 | 0.2055 0.0359 0.0488 | 14.1% 28.8% 17.3% |
| St. Helena Bay, South Africa Various Locations | Red Band Difference Blue-Green Chla Algorithm1 Neural Network Chla Algorithm2 | 90 74 75 | 0.5493 0.4283 0.3973 | 19.2% 20.8% 19.9% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houskeeper, H.F.; Kudela, R.M. Ocean Color Quality Control Masks Contain the High Phytoplankton Fraction of Coastal Ocean Observations. Remote Sens. 2019, 11, 2167. https://doi.org/10.3390/rs11182167
Houskeeper HF, Kudela RM. Ocean Color Quality Control Masks Contain the High Phytoplankton Fraction of Coastal Ocean Observations. Remote Sensing. 2019; 11(18):2167. https://doi.org/10.3390/rs11182167
Chicago/Turabian StyleHouskeeper, Henry F., and Raphael M. Kudela. 2019. "Ocean Color Quality Control Masks Contain the High Phytoplankton Fraction of Coastal Ocean Observations" Remote Sensing 11, no. 18: 2167. https://doi.org/10.3390/rs11182167
APA StyleHouskeeper, H. F., & Kudela, R. M. (2019). Ocean Color Quality Control Masks Contain the High Phytoplankton Fraction of Coastal Ocean Observations. Remote Sensing, 11(18), 2167. https://doi.org/10.3390/rs11182167