Repeat Oblique Photography Shows Terrain and Fire-Exposure Controls on Century-Scale Canopy Cover Change in the Alpine Treeline Ecotone

Abstract
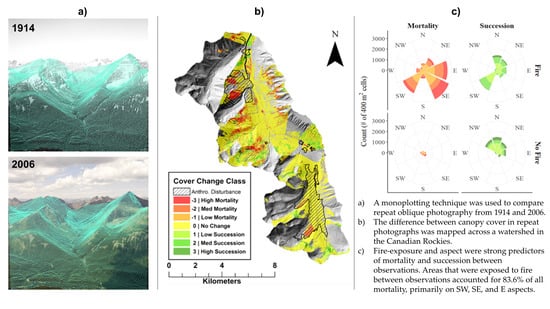
:
1. Introduction
1.1. Fire and Topography in the ATE
1.2. ATE Observation Techniques
1.3. Research Goals
2. Materials and Methods
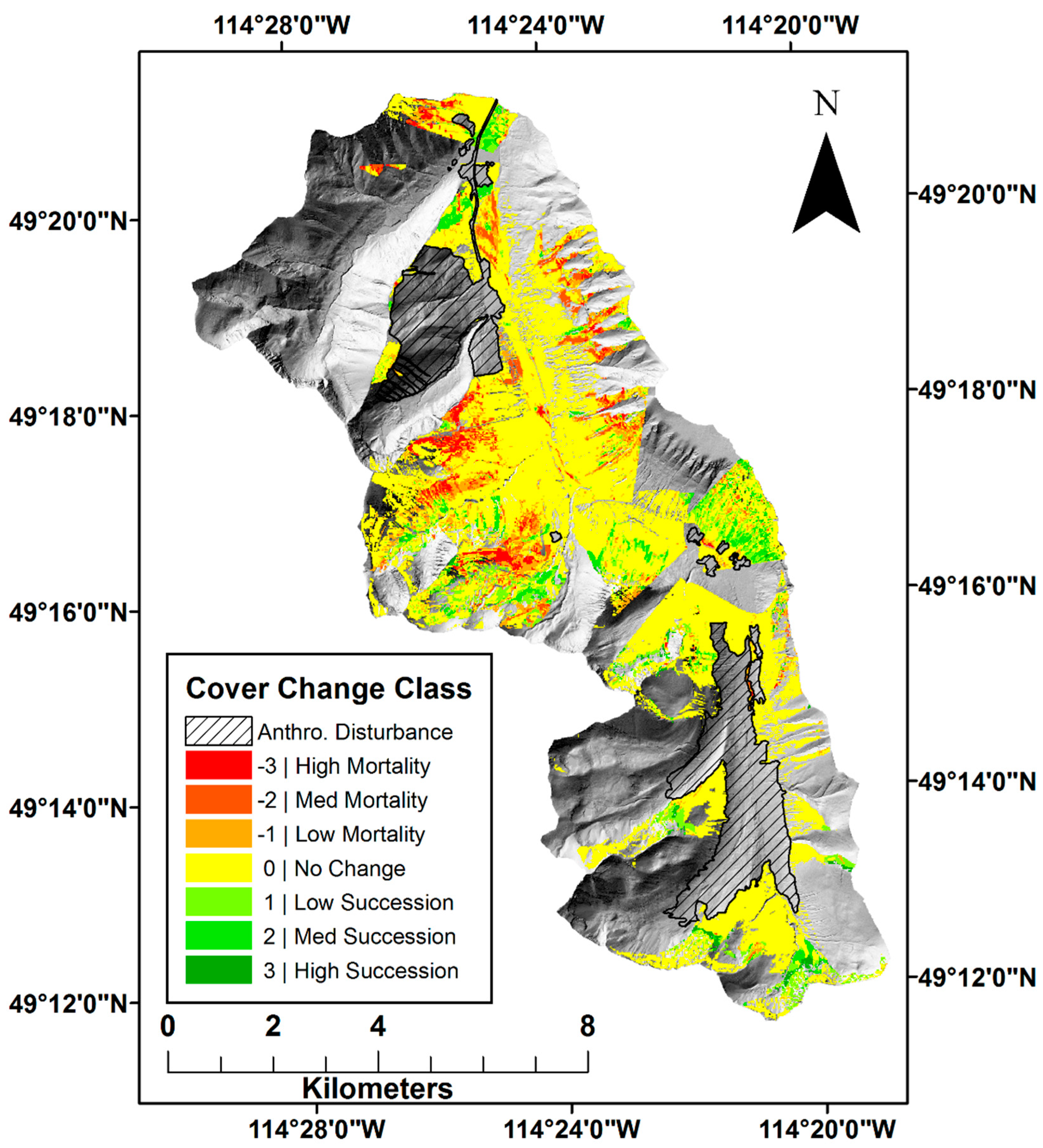
2.1. Study Area
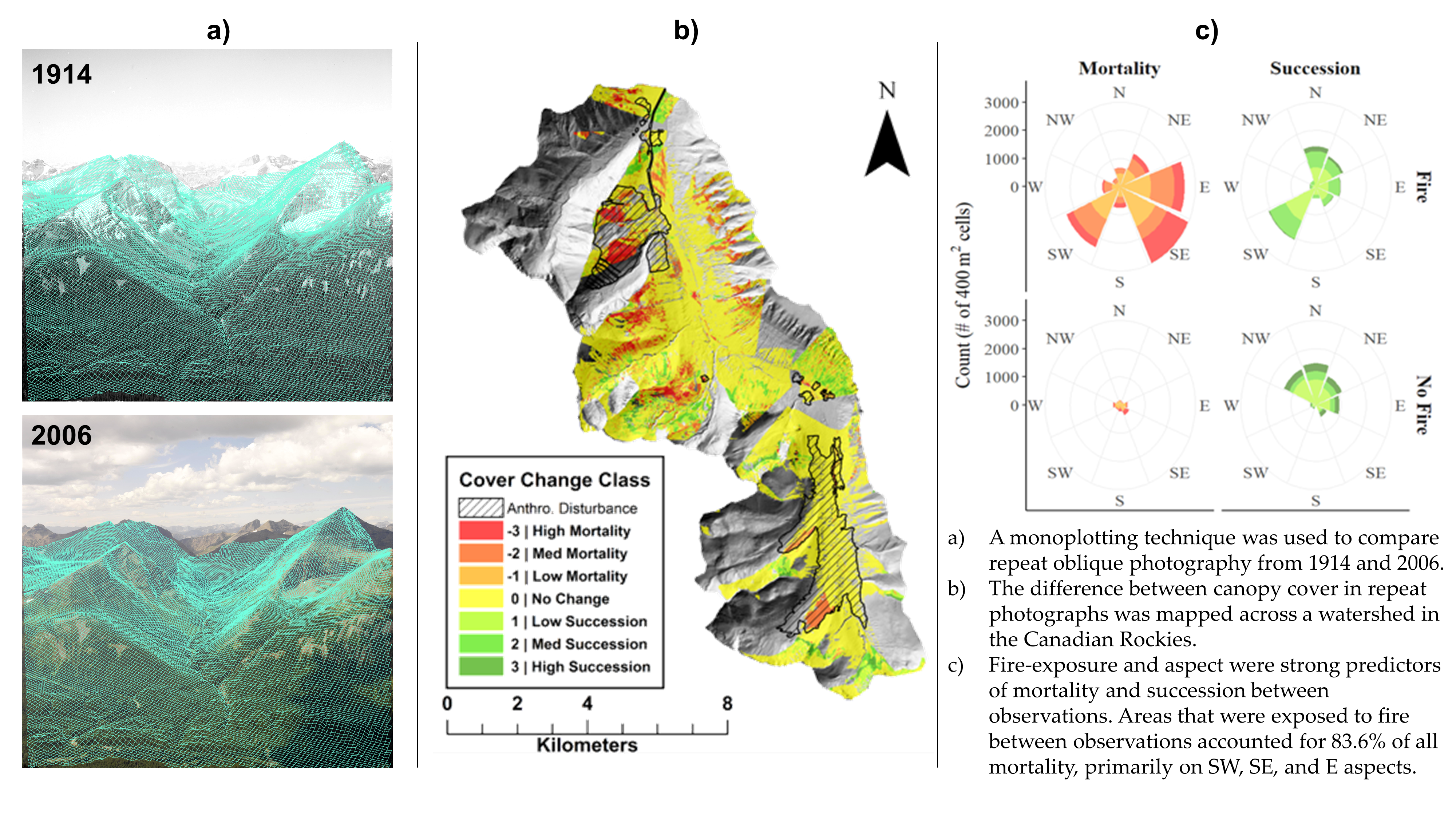
2.2. Repeat Photographs
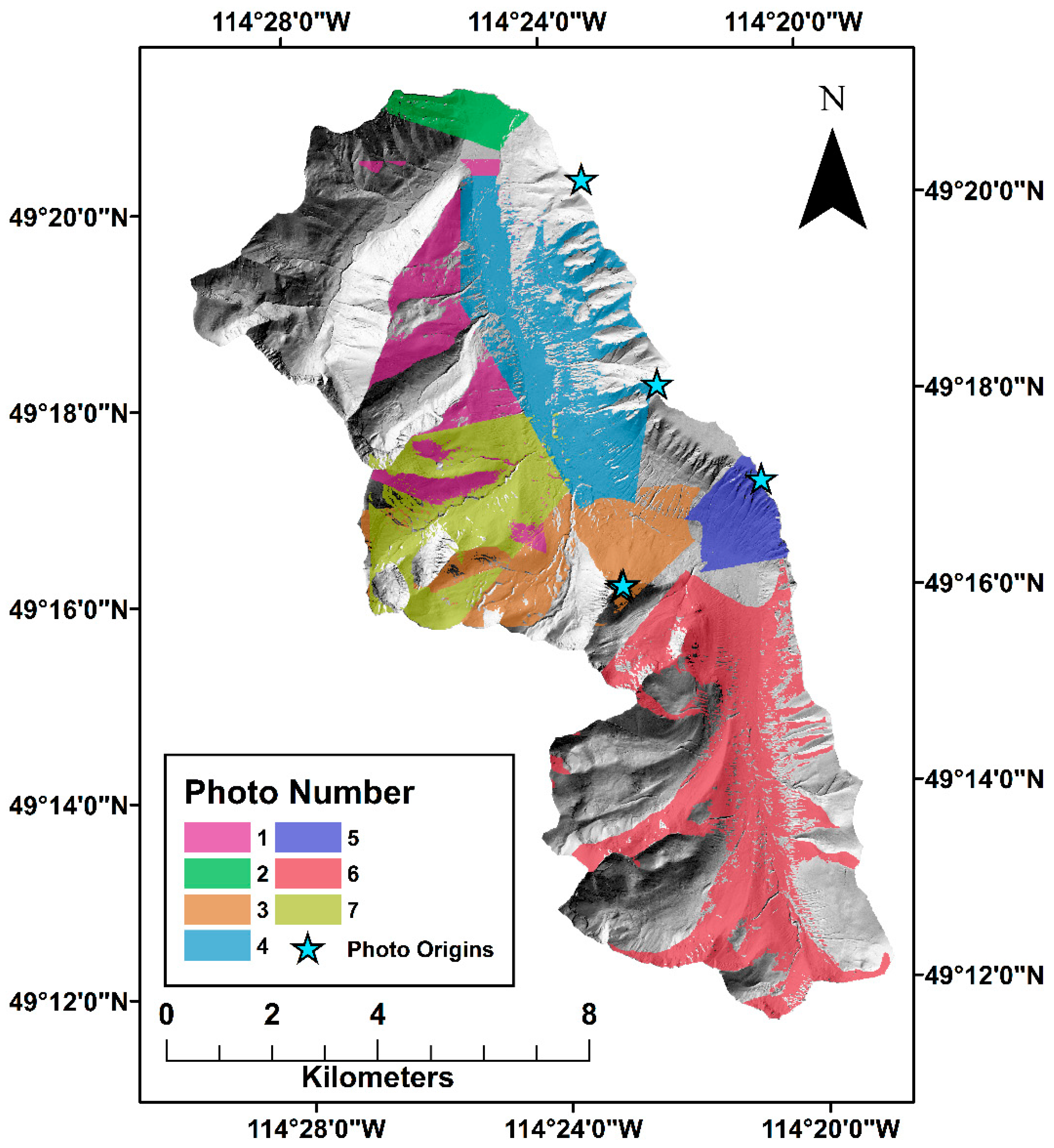
2.3. Canopy Classification
2.4. Anthropogenic Disturbance
2.5. Fire Disturbance
2.6. Topographic Analysis of Change
3. Results
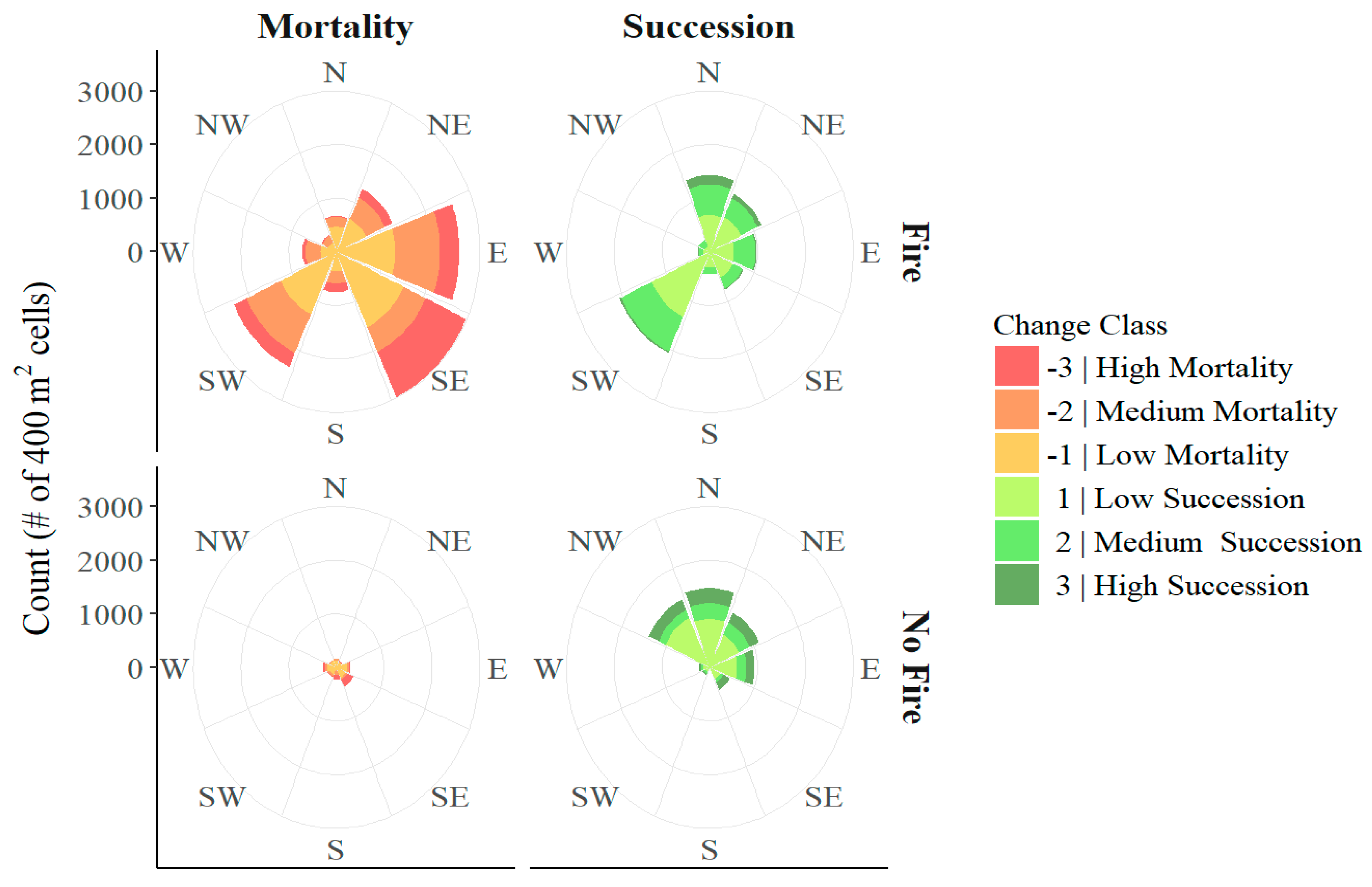
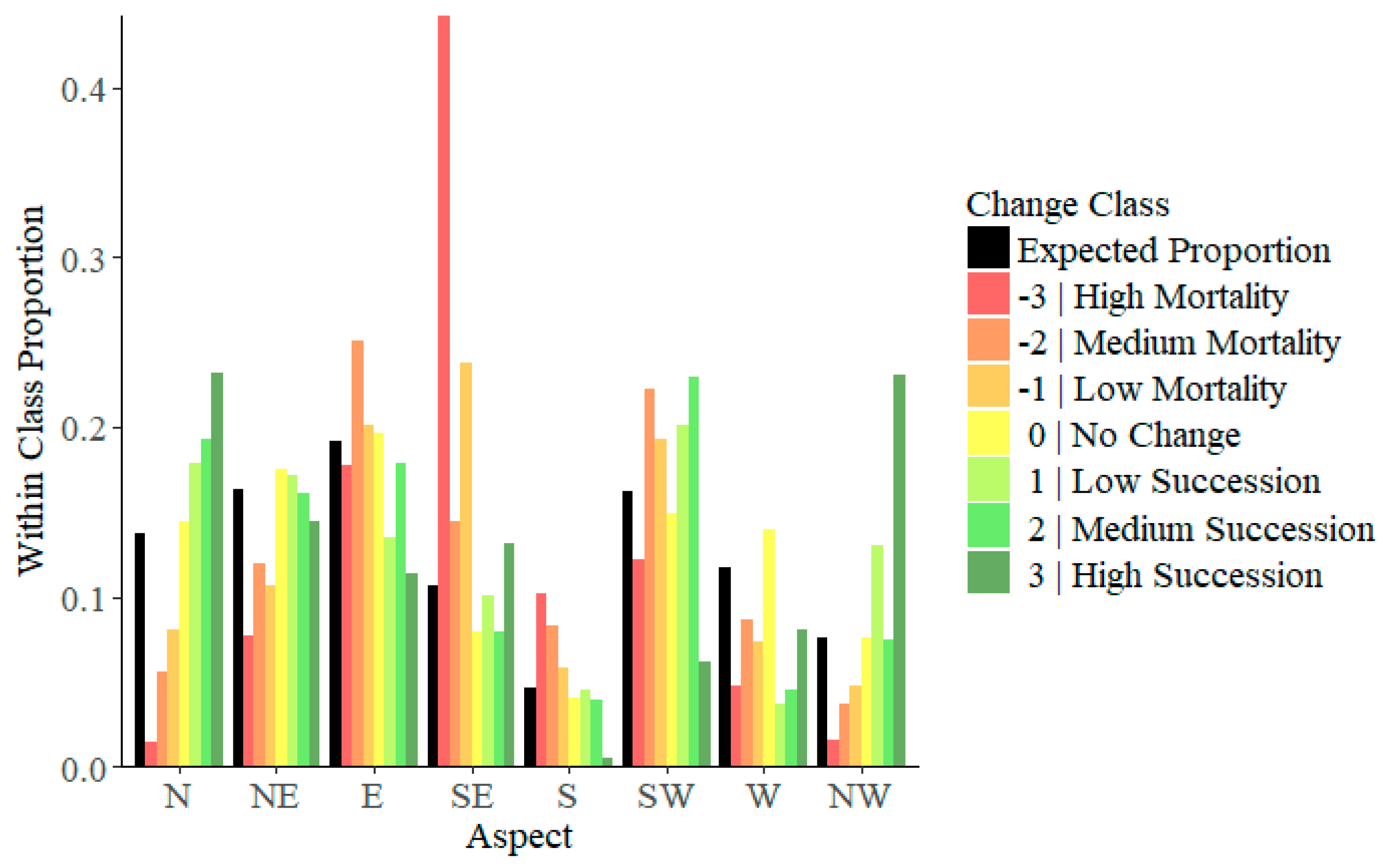
3.1. Aspect
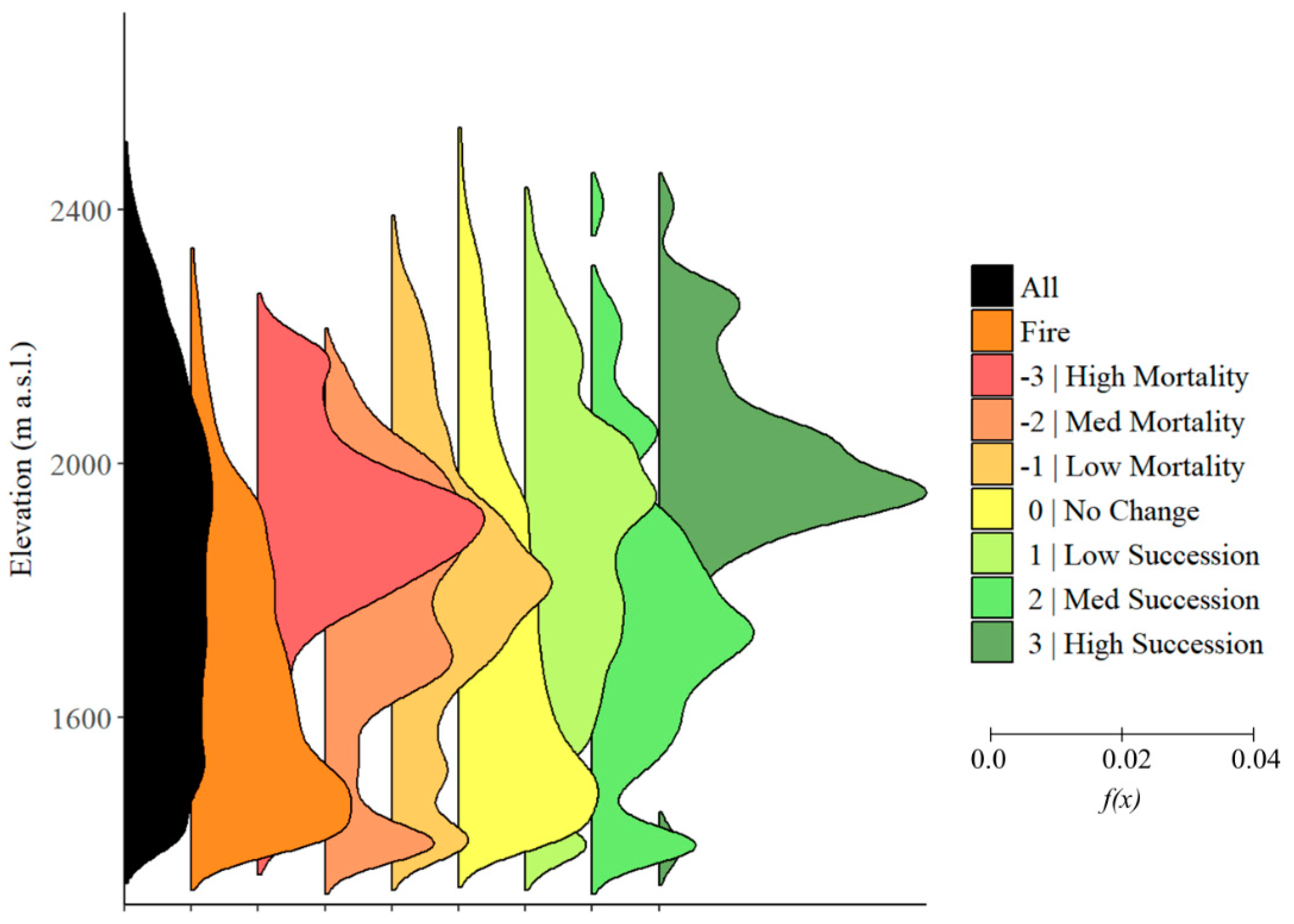
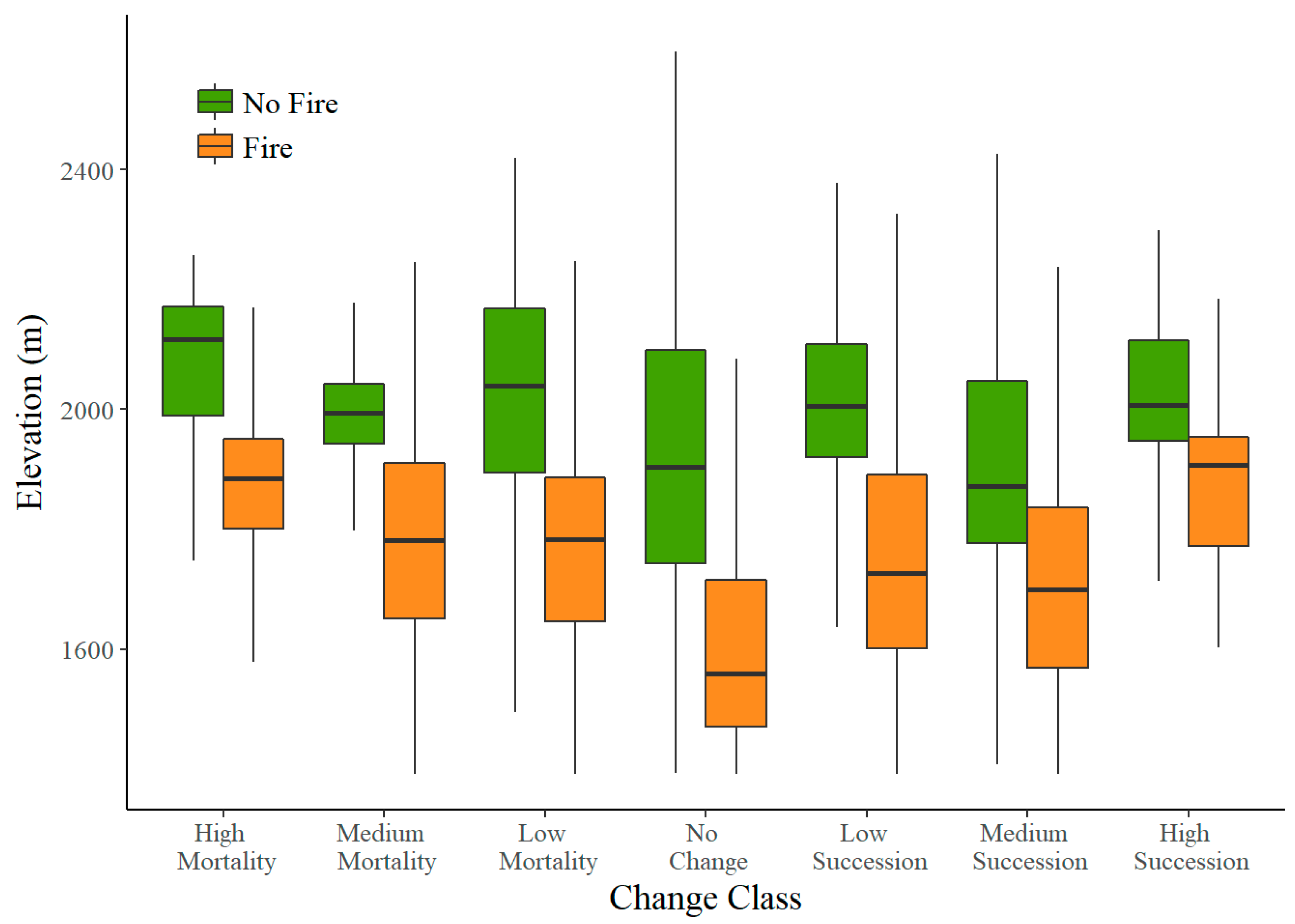
3.2. Elevation
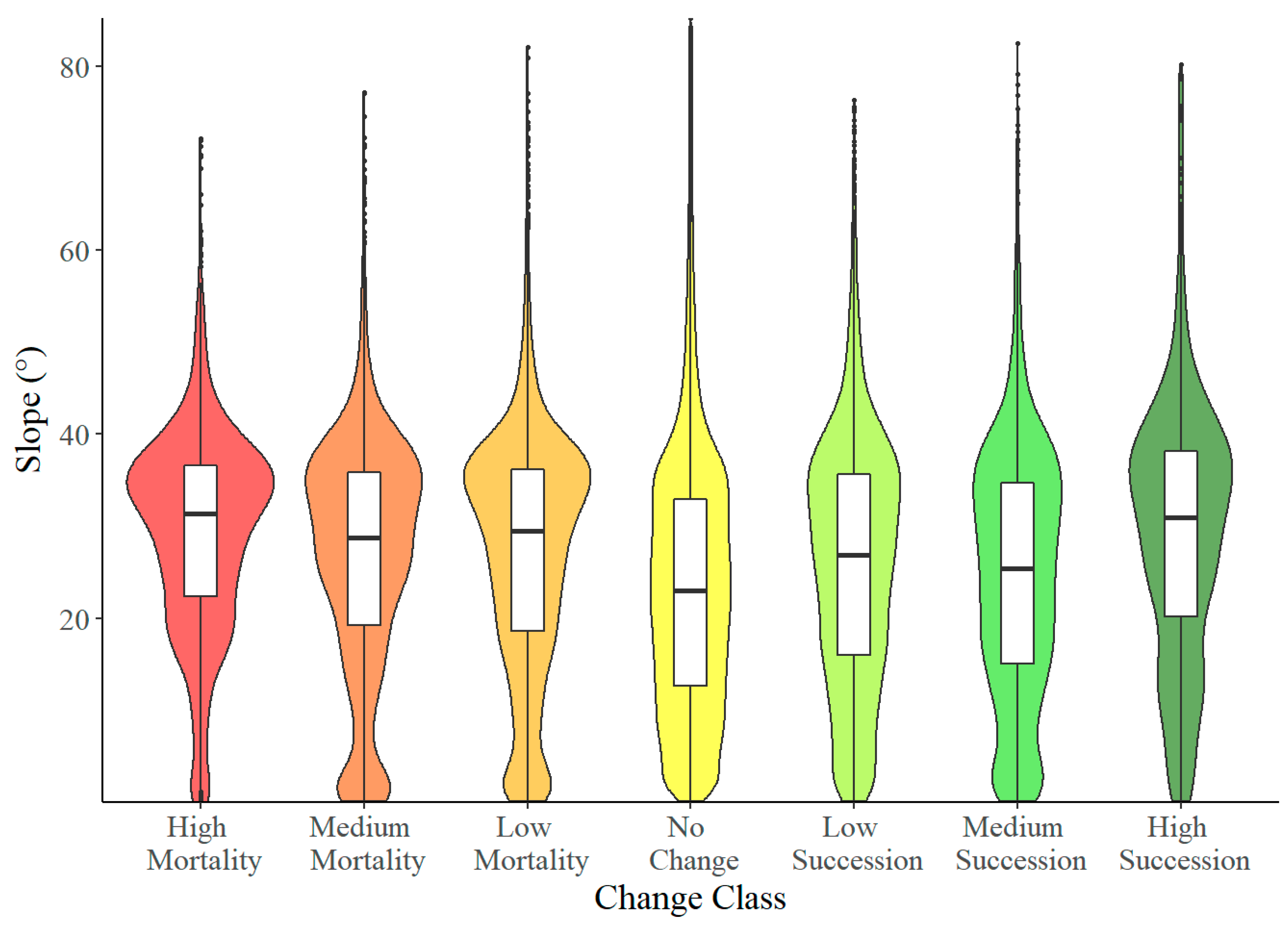
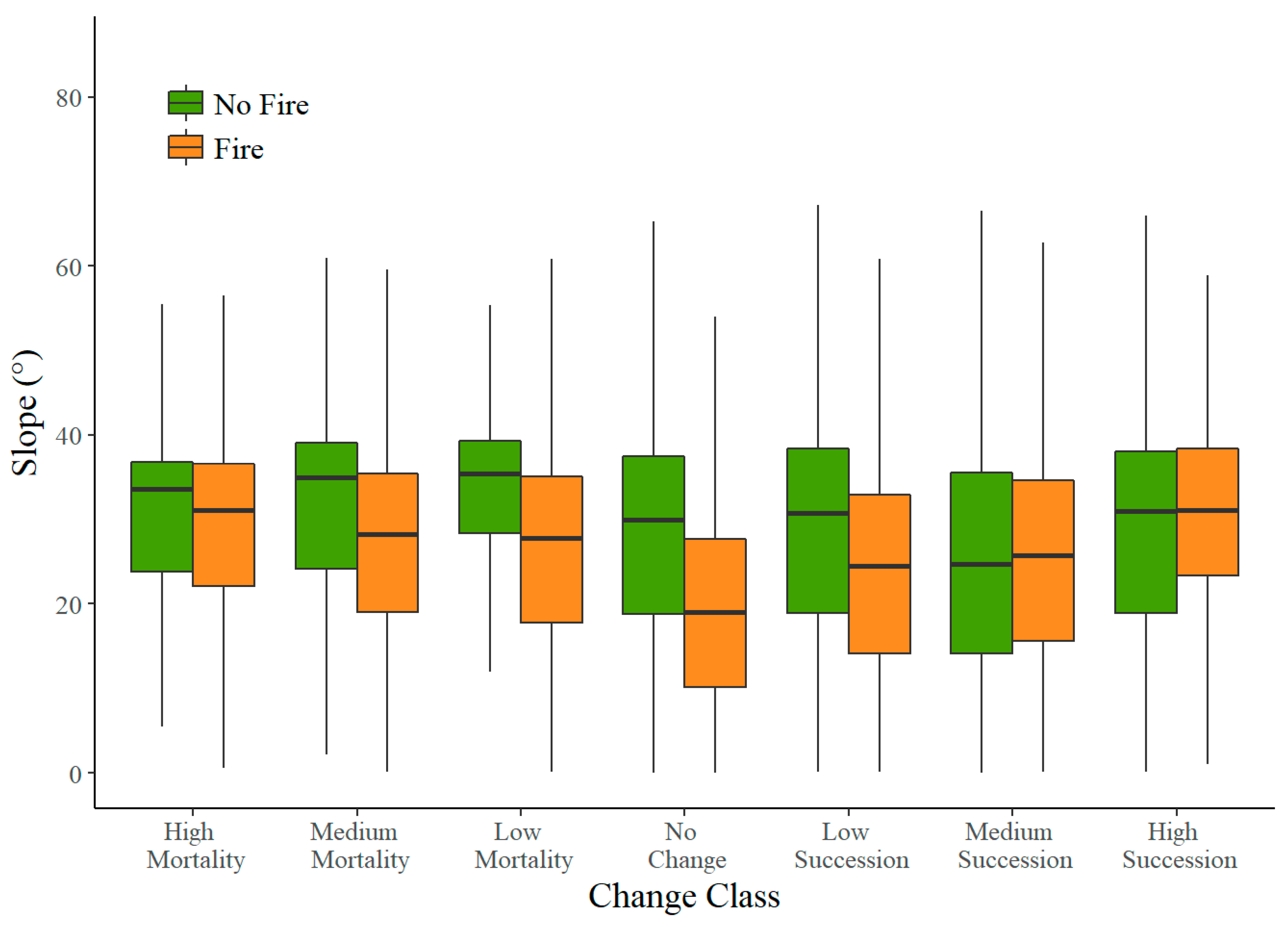
3.3. Slope
3.4. Upslope Advance
4. Discussion
4.1. Aspect
4.2. Elevation
4.3. Slope
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines: Ecology, Patchiness and Dynamics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; Volume 36. [Google Scholar]
- Case, B.; Duncan, R.P. A novel framework for disentangling the scale-dependent influences of abiotic factors on alpine treeline position. Ecography 2014, 37, 838–851. [Google Scholar] [CrossRef]
- Weiss, D.J.; Malanson, G.; Walsh, S. Multiscale Relationships Between Alpine Treeline Elevation and Hypothesized Environmental Controls in the Western United States. Ann. Assoc. Am. Geogr. 2015, 105, 437–453. [Google Scholar] [CrossRef]
- Holtmeier, F.-K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Butler, D.R.; Malanson, G.; Walsh, S.; Fagre, D.B. Influences of Geomorphology and Geology on Alpine Treeline in the American West—More Important than Climatic Influences? Phys. Geogr. 2007, 28, 434–450. [Google Scholar] [CrossRef] [Green Version]
- Resler, L.M. Geomorphic Controls of Spatial Pattern and Process at Alpine Treeline. Prof. Geogr. 2006, 58, 124–138. [Google Scholar] [CrossRef]
- Elliott, G.P.; Cowell, C.M. Slope Aspect Mediates Fine-Scale Tree Establishment Patterns at Upper Treeline during Wet and Dry Periods of the 20th Century. Arct. Antarct. Alp. Res. 2015, 47, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Holtmeier, F.-K.; Broll, G. Wind as an Ecological Agent at Treelines in North America, the Alps, and the European Subarctic. Phys. Geogr. 2010, 31, 203–233. [Google Scholar] [CrossRef]
- Walsh, S.J.; Butler, D.R.; Allen, T.R.; Malanson, G. Influence of snow patterns and snow avalanches on the alpine treeline ecotone. J. Veg. Sci. 1994, 5, 657–672. [Google Scholar] [CrossRef]
- Stine, M.B.; Butler, D.R. Effects of fire on geomorphic factors and seedling site conditions within the alpine treeline ecotone, Glacier National Park, MT. Catena 2015, 132, 37–44. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.L. Fire Ecology in Rocky Mountain Landscapes; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Rogeau, M.-P.; Armstrong, G.W. Quantifying the effect of elevation and aspect on fire return intervals in the Canadian Rocky Mountains. For. Ecol. Manag. 2017, 384, 248–261. [Google Scholar] [CrossRef]
- Colombaroli, D.; Henne, P.D.; Kaltenrieder, P.; Gobet, E.; Tinner, W. Species responses to fire, climate and human impact at tree line in the Alps as evidenced by palaeo-environmental records and a dynamic simulation model. J. Ecol. 2010, 98, 1346–1357. [Google Scholar] [CrossRef]
- Stueve, K.M.; Cerney, D.L.; Rochefort, R.M.; Kurth, L.L. Post-fire tree establishment patterns at the alpine treeline ecotone: Mount Rainier National Park, Washington, USA. J. Veg. Sci. 2009, 20, 107–120. [Google Scholar] [CrossRef]
- Cansler, C.A.; McKenzie, N.; Halpern, C.B. Area burned in alpine treeline ecotones reflects region-wide trends. Int. J. Wildland Fire 2016, 25, 1209. [Google Scholar] [CrossRef] [Green Version]
- Barrows, J. Fire behavior in northern Rocky Mountain forests. In USDA Forest Service Station Paper 29; Northern Rocky Mountain Forest and Range Experiment Station: Missoula, MT, USA, 1951. [Google Scholar]
- Fowler, P.M.; Asleson, D.O. The Location of Lightning-Caused Wildland Fires, Northern Idaho. Phys. Geogr. 1984, 5, 240–252. [Google Scholar] [CrossRef]
- Howe, E.; Baker, W.L. Landscape Heterogeneity and Disturbance Interactions in a Subalpine Watershed in Northern Colorado, USA. Ann. Assoc. Am. Geogr. 2003, 93, 797–813. [Google Scholar] [CrossRef]
- Baker, W.L.; Kipfmueller, K.F. Spatial Ecology of Pre–Euro-American Fires in a Southern Rocky Mountain Subalpine Forest Landscape. Prof. Geogr. 2001, 53, 248–262. [Google Scholar] [CrossRef]
- Buechling, A.; Baker, W.L. A fire history from tree rings in a high-elevation forest of Rocky Mountain National Park. Can. J. For. Res. 2004, 34, 1259–1273. [Google Scholar] [CrossRef]
- Aplet, G.H.; Laven, R.D.; Smith, F.W. Patterns of Community Dynamics in Colorado Engelmann Spruce-Subalpine Fir Forests. Ecology 1988, 69, 312–319. [Google Scholar] [CrossRef]
- Peet, R.K. Forest vegetation of the Colorado Front Range. Vegetatio 1981, 45, 3–75. [Google Scholar] [CrossRef]
- Romme, W.H.; Knight, D.H. Fire Frequency and Subalpine Forest Succession Along a Topographic Gradient in Wyoming. Ecology 1981, 62, 319–326. [Google Scholar] [CrossRef]
- Veblen, T.T. Age and Size Structure of Subalpine Forests in the Colorado Front Range. Bull. Torrey Bot. Club 1986, 113, 225. [Google Scholar] [CrossRef]
- Donnegan, J.A.; Rebertus, A.J. Rates and mechanisms of subalpine forest succession along an environmental gradient. Ecology 1999, 80, 1370–1384. [Google Scholar] [CrossRef]
- Kelsey, K.C.; Redmond, M.D.; Barger, N.N.; Neff, J. Species, Climate and Landscape Physiography Drive Variable Growth Trends in Subalpine Forests. Ecosystems 2017, 21, 125–140. [Google Scholar] [CrossRef]
- Redmond, M.D.; Kelsey, K.C. Topography and overstory mortality interact to control tree regeneration in spruce-fir forests of the southern Rocky Mountains. For. Ecol. Manag. 2018, 427, 106–113. [Google Scholar] [CrossRef]
- Danby, R.K. Monitoring Forest-Tundra Ecotones at Multiple Scales. Geogr. Compass 2011, 5, 623–640. [Google Scholar] [CrossRef]
- Allen, T.R.; Walsh, S.J. Spatial and compositional pattern of alpine treeline, Glacier National Park, Montana. Photogramm. Eng. Remote Sens. 1996, 62, 1261–1268. [Google Scholar]
- Bolton, D.K.; Coops, N.C.; Hermosilla, T.; Wulder, M.A.; White, J.C. Evidence of vegetation greening at alpine treeline ecotones: Three decades of Landsat spectral trends informed by lidar-derived vertical structure. Environ. Res. Lett. 2018, 13, 084022. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Evidence of Recent Treeline Dynamics in Southwest Yukon from Aerial Photographs. Arctic 2009, 60, 60. [Google Scholar] [CrossRef] [Green Version]
- Tinner, W.; Theurillat, J. Institute of Arctic and Alpine Research (INSTAAR) University of Colorado Uppermost Limit, Extent, and Fluctuations of the Timberline and Treeline Ecocline in the Swiss Central Alps during the Past 11,500 Years. Arct. Antarct. Alp. Res. 2003, 35, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Bekker, M.F. Positive Feedback Between Tree Establishment and Patterns of Subalpine Forest Advancement, Glacier National Park, Montana, USA. Arct. Antarct. Alp. Res. 2005, 37, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Mamet, S.D.; Kershaw, G.P. Subarctic and alpine tree line dynamics during the last 400 years in north-western and central Canada. J. Biogeogr. 2011, 39, 855–868. [Google Scholar] [CrossRef]
- Sakulich, J. Reconstruction and spatial analysis of alpine treeline in the Elk Mountains, Colorado, USA. Phys. Geogr. 2015, 36, 471–488. [Google Scholar] [CrossRef]
- Butler, D.R.; DeChano, L.M. Environmental Change in Glacier National Park, Montana: An Assessment through Repeat Photography from Fire Lookouts. Phys. Geogr. 2001, 22, 291–304. [Google Scholar] [CrossRef]
- Butler, D.R.; Malanson, G.P.; Cairns, D. Stability of alpine treeline in Glacier National Park, Montana, USA. Phytocoenologia 1994, 22, 485–500. [Google Scholar] [CrossRef]
- Klasner, F.L.; Fagre, D.B. A Half Century of Change in Alpine Treeline Patterns at Glacier National Park, Montana, USA. Arct. Antarct. Alp. Res. 2002, 34, 49–56. [Google Scholar] [CrossRef]
- Kullman, L.; Öberg, L. Post-Little Ice Age tree line rise and climate warming in the Swedish Scandes: A landscape ecological perspective. J. Ecol. 2009, 97, 415–429. [Google Scholar] [CrossRef]
- Moiseev, P.A.; Shiyatov, S.G. Vegetation Dynamics at the Tree-Line Ecotone in the Ural Highlands, Russia. In Alpine Biodiversity in Europe; Springer Science and Business Media: Berlin, Germany, 2003; Volume 167, pp. 423–435. [Google Scholar]
- Roush, W.; Munroe, J.S.; Fagre, D.B. Development of a spatial analysis metho using ground-based repeat photography to detect changes in the alpine treeline ecotone, Glacier National Park, Montana, USA. Arct. Antarct. Alp. Res. 2007, 39, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Claudio, B.; Conedera, M.; Krebs, P. A New Monoplotting Tool to Extract Georeferenced Vector Data and Orthorectified Raster Data from Oblique Non-Metric Photographs. Int. J. Herit. Digit. Era 2012, 1, 499–518. [Google Scholar] [CrossRef]
- Stockdale, C.A.; Bozzini, C.; Macdonald, S.E.; Higgs, E.; Macdonald, S.E. Extracting ecological information from oblique angle terrestrial landscape photographs: Performance evaluation of the WSL Monoplotting Tool. Appl. Geogr. 2015, 63, 315–325. [Google Scholar] [CrossRef]
- Stockdale, C.A.; Macdonald, S.E.; Higgs, E. Forest closure and encroachment at the grassland interface: A century-scale analysis using oblique repeat photography. Ecosphere 2019, 10, e02774. [Google Scholar] [CrossRef]
- Trant, A.J.; Starzomski, B.M.; Higgs, E. A publically available database for studying ecological change in mountain ecosystems. Front. Ecol. Environ. 2015, 13, 187. [Google Scholar] [CrossRef]
- McCaffrey, D.R.; Hopkinson, C. Assessing Fractional Cover in the Alpine Treeline Ecotone Using the WSL Monoplotting Tool and Airborne Lidar. Can. J. Remote Sens. 2017, 43, 504–512. [Google Scholar] [CrossRef]
- Wildfire Management Branch—Alberta Agriculture and Forestry. Fire History Polygons C5 FMU. 2017. Available online: https://wildfire.alberta.ca/resources/historical-data/spatial-wildfire-data.aspx (accessed on 14 May 2020).
- Alberta Environment and Parks. Alberta Vegetation Inventory; Branch, R.I.M., Ed.; Alberta Environment and Parks: Edmonton, AB, Canada, 2012. [Google Scholar]
- Rogeau, M.-P.; Flannigan, M.; Hawkes, B.C.; Parisien, M.-A.; Arthur, R. Spatial and temporal variations of fire regimes in the Canadian Rocky Mountains and Foothills of southern Alberta. Int. J. Wildland Fire 2016, 25, 1117–1130. [Google Scholar] [CrossRef]
- Rogeau, M.P. Fire History Study Castle River Watershed; Alberta. Technical report prepared for Alberta Environment and Sustainable Resource Development; Forest Protection Branch, Southern Rockies Wildfire Management Area: Calgary, AB, Canada, 2012; 54p. [Google Scholar]
- Lumley, T.; Diehr, P.; Emerson, S.; Chen, L. The Importance of the Normality Assumption in Large Public Health Data Sets. Annu. Rev. Public Heal. 2002, 23, 151–169. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Kueppers, L.M.; Conlisk, E.; Castanha, C.; Moyes, A.; Germino, M.J.; De Valpine, P.; Torn, M.S.; Mitton, J.B. Warming and provenance limit tree recruitment across and beyond the elevation range of subalpine forest. Glob. Chang. Boil. 2016, 23, 2383–2395. [Google Scholar] [CrossRef] [Green Version]
- Moyes, A.; Germino, M.J.; Kueppers, L.M. Moisture rivals temperature in limiting photosynthesis by trees establishing beyond their cold-edge range limit under ambient and warmed conditions. New Phytol. 2015, 207, 1005–1014. [Google Scholar] [CrossRef]
- Silins, U.; Stone, M.; Emelko, M.B.; Bladon, K.D. Sediment production following severe wildfire and post-fire salvage logging in the Rocky Mountain headwaters of the Oldman River Basin, Alberta. Catena 2009, 79, 189–197. [Google Scholar] [CrossRef]
- Hopkinson, C.; Collins, T.; Anderson, A.; Pomeroy, J.; Spooner, I. Spatial Snow Depth Assessment Using LiDAR Transect Samples and Public GIS Data Layers in the Elbow River Watershed, Alberta. Can. Water Resour. J. 2012, 37, 69–87. [Google Scholar] [CrossRef]
- Bader, M.; Ruijten, J.J. A topography-based model of forest cover at the alpine tree line in the tropical Andes. J. Biogeogr. 2008, 35, 711–723. [Google Scholar] [CrossRef]
- Dang, H.; Zhang, Y.; Zhang, Y.; Zhang, K.; Zhang, Q. Variability and rapid response of subalpine fir (Abies fargesii) to climate warming at upper altitudinal limits in north-central China. Trees 2015, 29, 785–795. [Google Scholar] [CrossRef]
- Greenwood, S.; Chen, J.-C.; Chen, C.-T.; Jump, A.S. Strong topographic sheltering effects lead to spatially complex treeline advance and increased forest density in a subtropical mountain region. Glob. Chang. Boil. 2014, 20, 3756–3766. [Google Scholar] [CrossRef] [PubMed]
- Luckman, B.H.; Wilson, R. Summer temperatures in the Canadian Rockies during the last millennium: A revised record. Clim. Dyn. 2005, 24, 131–144. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Tranquillini, W. Physiological Ecology of the Alpine Timberline: Tree Existence at High Altitudes with Special Reference to the European Alps; Springer: Berlin, Germany, 1979. [Google Scholar]
- Butler, D.R.; Walsh, S. Site Characteristics of Debris Flows and their Relationship to Alpine Treeline. Phys. Geogr. 1994, 15, 181–199. [Google Scholar] [CrossRef]
- Shakesby, R.; Doerr, S. Wildfire as a hydrological and geomorphological agent. Earth Sci. Rev. 2006, 74, 269–307. [Google Scholar] [CrossRef]
- Werth, P.A.; Potter, B.E.; Clements, C.; Finney, M.A.; Goodrick, S.L.; Alexander, M.E.; Cruz, M.G.; Forthofer, J.; McAllister, S. Synthesis of Knowledge of Extreme Fire Behavior: Volume I for Fire Managers; USDA: Washington, DC, USA, 2011. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
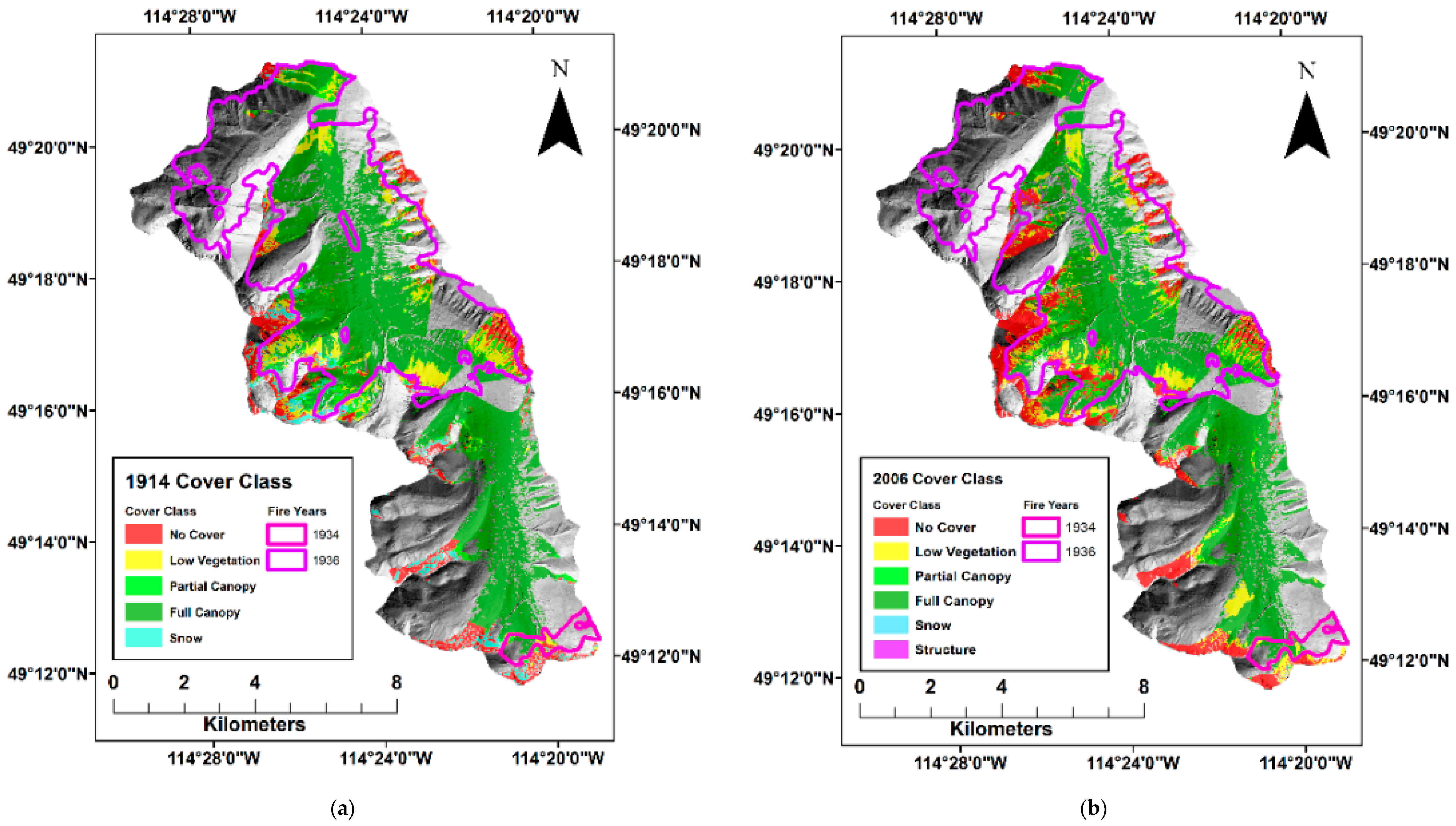{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photograph Number | Elevation (m a.s.l.) | Latitude (N) | Longitude (W) | Photograph Date | MLP Photo ID (2006) |
|---|---|---|---|---|---|
| 1 | 2295 | 49°16′3.62″ | 114°22′58.97″ | 28 July 2006 | B0032P0048 |
| 2 | 2216 | 49°20′14.00″ | 114°23′22.81″ | 29 July 2006 | B0033P0005 |
| 3 | 2390 | 49°17′7.01″ | 114°20′44.21″ | 28 July 2006 | B0032P0073 |
| 4 | 2283 | 49°16′4.94″ | 114°22′56.77″ | 28 July 2006 | B0032P0056 |
| 5 | 2283 | 49°16′4.94″ | 114°22′56.77″ | 28 July 2006 | B0032P0055 |
| 6 | 2390 | 49°17′7.01″ | 114°20′44.21″ | 28 July 2006 | B0032P0075 |
| 7 | 2462 | 49°18′7.26″ | 114°22′18.63″ | 30 July 2006 | B0034P0005 |
| Aspect | Change Class | |||||||
|---|---|---|---|---|---|---|---|---|
| High Mortality | Medium Mortality | Low Mortality | No Change | Low Succession | Medium Succession | High Succession | ||
| N | χ2 | 307.5 | 212.1 | 196.8 | 28.6 | 101.4 | 90.0 | 59.2 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| NE | χ2 | 130.1 | 52.6 | 171.7 | 64.5 | 3.4 | 0.1 | 1.9 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.066 | 0.728 | 0.163 | |
| E | χ2 | 3.3 | 85.9 | 4.4 | 6.1 | 147.0 | 4.2 | 30.7 |
| P | 0.069 | <0.001 * | 0.037 | 0.013 | <0.001 * | 0.041 | <0.001 * | |
| SE | χ2 | 2863.3 | 55.9 | 1327.0 | 530.7 | 2.6 | 27.5 | 5.1 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.109 | <0.001 * | 0.024 | |
| S | χ2 | 177.1 | 121.3 | 26.7 | 34.4 | 0.1 | 3.3 | 29.6 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.702 | 0.070 | <0.001 * | |
| SW | χ2 | 27.8 | 101.1 | 52.0 | 71.2 | 81.5 | 114.1 | 58.0 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| W | χ2 | 114.1 | 34.8 | 136.6 | 311.9 | 434.0 | 169.8 | 10.2 |
| P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| NW | χ2 | 127.0 | 81.2 | 81.9 | 0.0 | 301.4 | 0.1 | 268.0 |
| P | <0.001 * | <0.001 * | <0.001 * | 0.963 | <0.001 * | 0.806 | <0.001 * | |
| Change Class | |||||||
|---|---|---|---|---|---|---|---|
| High Mortality | Medium Mortality | Low Mortality | No Change | Low Succession | Medium Succession | ||
| Medium | t | −24.042 | - | - | - | - | - |
| Mortality | p | <0.001 * | - | - | - | - | - |
| Low | t | −20.153 | 6.643 | - | - | - | - |
| Mortality | p | <0.001 * | <0.001 * | - | - | - | - |
| No | t | −42.218 | −10.400 | −22.928 | - | - | - |
| Change | p | <0.001 * | <0.001 * | <0.001 * | - | - | - |
| Low | t | −7.627 | 18.686 | 13.900 | 39.454 | - | - |
| Succession | p | <0.001 * | <0.001 * | <0.001 * | <0.001 * | - | - |
| Medium | t | −24.636 | −2.525 | −8.754 | 5.790 | −19.643 | - |
| Succession | p | <0.001 * | 0.244 | <0.001 * | <0.001 * | <0.001 * | - |
| High | t | 13.569 | 30.010 | 27.219 | 39.397 | 19.105 | 30.610 |
| Succession | p | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * |
| Change Class | |||||||
|---|---|---|---|---|---|---|---|
| High Mortality | Medium Mortality | Low Mortality | No Change | Low Succession | Medium Succession | ||
| Medium | t | −8.609 | - | - | - | - | - |
| Mortality | p | <0.001 * | - | - | - | - | - |
| Low | t | −8.727 | 1.097 | - | - | - | - |
| Mortality | p | <0.001 * | 1.000 | - | - | - | - |
| No | t | −24.947 | −14.348 | −21.092 | - | - | - |
| Change | p | <0.001 * | <0.001 * | <0.001 * | - | - | - |
| Low | t | −12.396 | −2.926 | −4.793 | 13.790 | - | - |
| Succession | p | <0.001 * | 0.072 | <0.001 * | <0.001 * | - | - |
| Medium Succession | t | −14.204 | −6.250 | −8.061 | 4.742 | −4.204 | - |
| p | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | - | |
| High | t | −0.064 | 4.710 | 4.350 | 11.106 | 6.297 | 8.120 |
| Succession | p | 1.000 | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * |
| Transect | Number of Cells 1914 | Mean Elevation 1914 (m a.s.l.) | Number of Cells 2006 | Mean Elevation 2006 (m a.s.l.) | Elevation Difference (m a.s.l.) |
|---|---|---|---|---|---|
| 1 | 6 | 2046.0 | 8 | 2053.5 | 7.5 |
| 2 | 7 | 1894.6 | 9 | 1897.0 | 2.4 |
| 3 | 35 | 2002.9 | 28 | 2028.5 | 25.6 |
| 4 | 24 | 1945.1 | 24 | 2033.5 | 88.4 |
| 5 | 29 | 1876.1 | 31 | 1954.7 | 78.6 |
| Mean | 20.2 | 1952.9 | 20.0 | 1993.4 | 40.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCaffrey, D.; Hopkinson, C. Repeat Oblique Photography Shows Terrain and Fire-Exposure Controls on Century-Scale Canopy Cover Change in the Alpine Treeline Ecotone. Remote Sens. 2020, 12, 1569. https://doi.org/10.3390/rs12101569
McCaffrey D, Hopkinson C. Repeat Oblique Photography Shows Terrain and Fire-Exposure Controls on Century-Scale Canopy Cover Change in the Alpine Treeline Ecotone. Remote Sensing. 2020; 12(10):1569. https://doi.org/10.3390/rs12101569
Chicago/Turabian StyleMcCaffrey, David, and Chris Hopkinson. 2020. "Repeat Oblique Photography Shows Terrain and Fire-Exposure Controls on Century-Scale Canopy Cover Change in the Alpine Treeline Ecotone" Remote Sensing 12, no. 10: 1569. https://doi.org/10.3390/rs12101569
APA StyleMcCaffrey, D., & Hopkinson, C. (2020). Repeat Oblique Photography Shows Terrain and Fire-Exposure Controls on Century-Scale Canopy Cover Change in the Alpine Treeline Ecotone. Remote Sensing, 12(10), 1569. https://doi.org/10.3390/rs12101569