Improved Remote Sensing Methods to Detect Northern Wild Rice (Zizania palustris L.)




 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
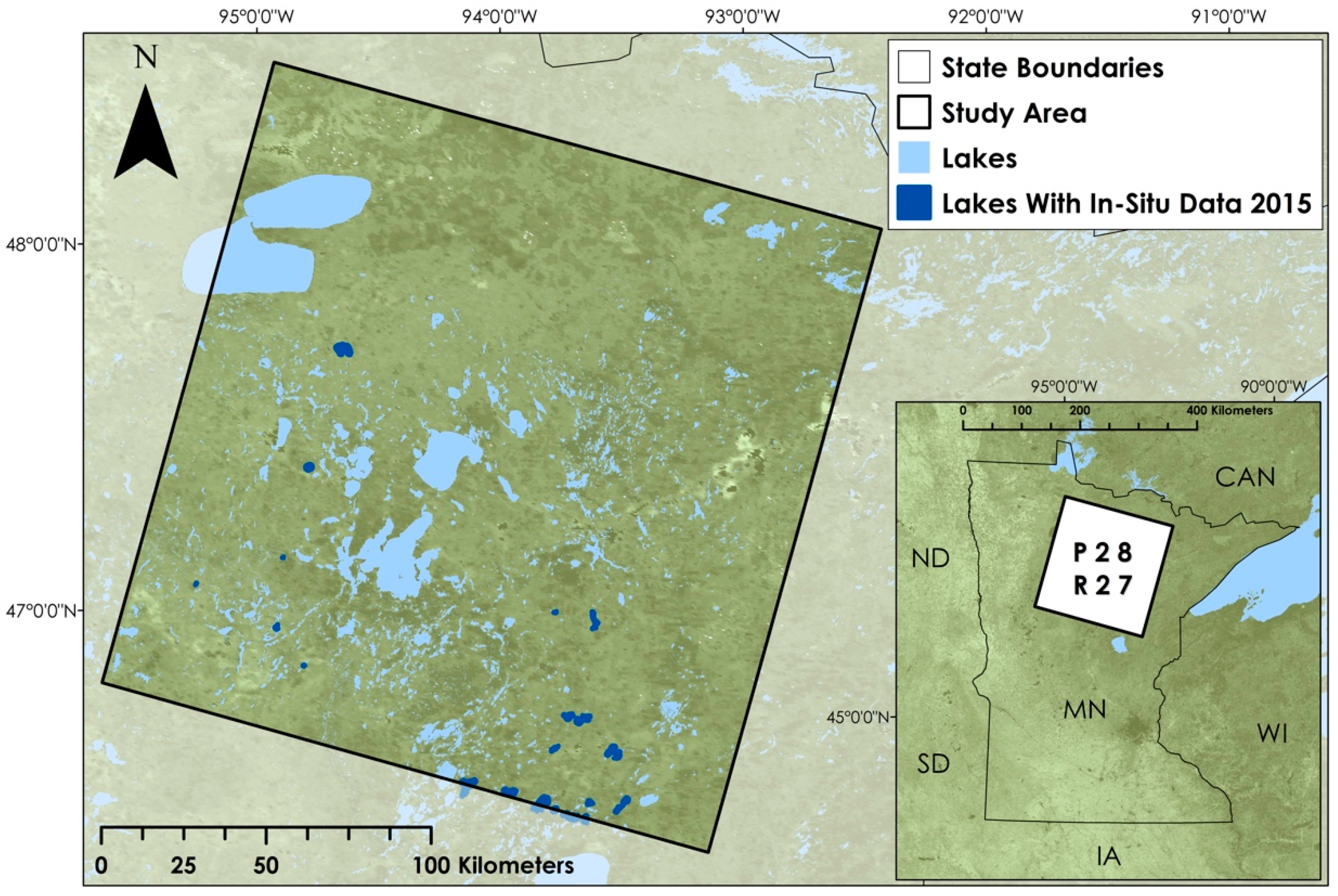
2.1. Study Area and Study Period
2.2. Training and Validation Datasets
2.3. Training and Validation Dataset Modifications
2.4. Satellite-Derived Spectral Data
2.5. Modeling
2.6. Point vs. Area-Based Validation
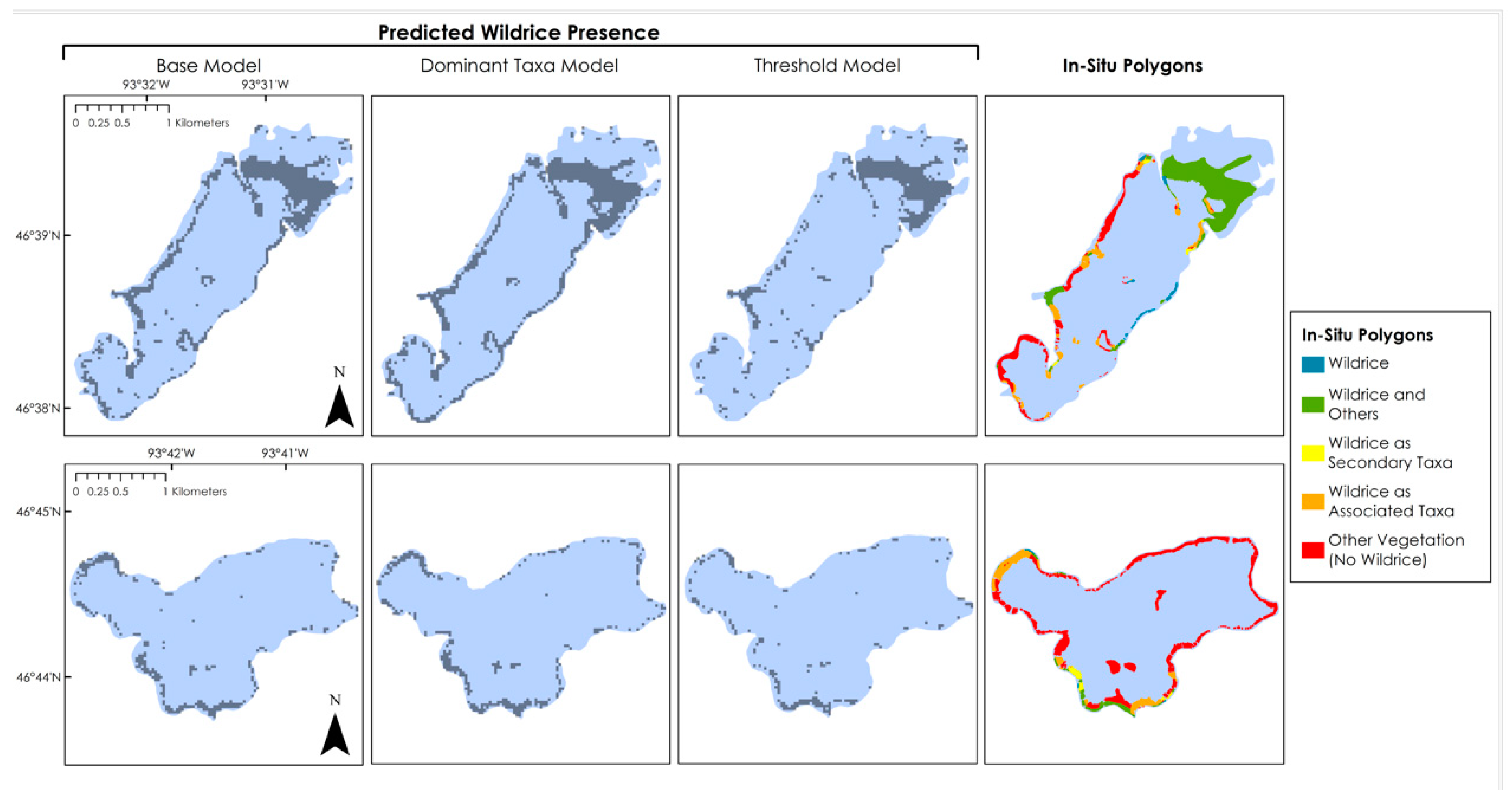
3. Results
3.1. Model Validation
3.2. Predictor Variable Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral Derivative | Equation | Description | Source |
|---|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | NDVI = (NIR − Red)/(NIR + Red) | A remote sensing indicator that can be used to assess vegetation content | [38] |
| Normalized Difference Moisture Index (NDMI) | NDMI = (NIR − SWIR)/(NIR + SWIR) | A remote sensing indicator that can be used to assess water content | [38] |
| Tasseled Cap Wetness | Blue (0.1511) + Green (0.1973) + Red (0.3283) + NIR (0.3407) + SWIR (−0.7117) + SWIR2 (−0.4559) | A linear transformation of spectral bands, used to distinguish wet surfaces and measure water content | [39] |
| Tasseled Cap Greenness | Blue (−0.2941) + Green (−0.243) + Red (−0.5424) + NIR (0.7276) + SWIR (0.0713) + SWIR2 (−0.1608) | A linear transformation of spectral bands, used to distinguish vegetated surfaces and measure vegetation content | [28] |
| Tasseled Cap Brightness | Blue (0.3029) + Green (0.2786) + Red (0.4733) + NIR (0.5599) + SWIR (0.508) + SWIR2 (0.1872) | A linear transformation of spectral bands, used to distinguish bright surfaces and measure brightness content | [28] |
| Radar Range | Radar Median T1 − Radar Median T2 | The range of the radar backscatter signal over the growing season (T1: June–July, T2: August–September) | [17] |
| Radar Variance | The variance of the radar backscatter signal over the growing season | ||
| Radar Cross-Ratio | (Radar VV Median)/Radar (VH) Median or (Radar VH Median)/Radar (VV) Median | The ratio of VV and VH radar polarizations for a certain time period. |
| Model | Validation Set | PCC | Sensitivity (Presence Accuracy) | Specificity (Absence Accuracy) | Confusion Matrix | |||
|---|---|---|---|---|---|---|---|---|
| TP | TN | FP | FN | |||||
| Base | Base | 83.8% | 83.3% | 84.2% | 365 | 523 | 98 | 73 |
| 83.3% | 84.2% | 15.8% | 16.7% | |||||
| Base | Dominant Taxa | 83.5% | 86.0% | 82.1% | 257 | 437 | 95 | 42 |
| 86.0% | 82.1% | 17.9% | 14.0% | |||||
| Base | Threshold | 85.0% | 91.1% | 82.1% | 234 | 437 | 95 | 23 |
| 91.1% | 82.1% | 17.9% | 14.0% | |||||
| Dominant Taxa | Dominant Taxa | 87.5% | 81.6% | 90.8% | 244 | 483 | 49 | 55 |
| 81.6% | 90.8% | 9.2% | 18.4% | |||||
| Dominant Taxa | Base | 79.6% | 64.7% | 90.2% | 282 | 560 | 61 | 154 |
| 64.7% | 90.2% | 9.8% | 35.3% | |||||
| Dominant Taxa | Threshold | 90.0% | 87.9% | 91.0% | 226 | 484 | 48 | 31 |
| 87.9% | 91.1% | 9.0% | 12.1% | |||||
| Threshold | Threshold | 91.1% | 89.5% | 92.0% | 230 | 459 | 40 | 27 |
| 89.5% | 92.0% | 8.0% | 10.5% | |||||
| Threshold | Dominant Taxa | 85.6% | 76.3% | 90.8% | 228 | 483 | 49 | 71 |
| 76.3% | 90.8% | 9.2% | 23.7% | |||||
| Threshold | Base | 79.4% | 62.2% | 91.5% | 271 | 568 | 53 | 165 |
| 62.2% | 91.5% | 8.5% | 37.8% | |||||
| Model Trained | Model Validated | Total False Positives | Rushes | Water Lilies | Rushes and Others | Water lilies and Others | Other | Total Vegetation Absence Points |
|---|---|---|---|---|---|---|---|---|
| Base | Base | 98 | 45 | 15 | 13 | 10 | 15 | 422 |
| Base | Dominant Taxa | 95 | 41 | 14 | 10 | 11 | 19 | 333 |
| Base | Threshold | 95 | 41 | 14 | 10 | 11 | 19 | 300 |
| Dominant Taxa | Base | 61 | 23 | 13 | 10 | 5 | 10 | 422 |
| Dominant Taxa | Dominant Taxa | 49 | 20 | 10 | 7 | 5 | 7 | 333 |
| Dominant Taxa | Threshold | 48 | 20 | 10 | 6 | 5 | 7 | 300 |
| Threshold | Base | 53 | 21 | 11 | 10 | 2 | 9 | 422 |
| Threshold | Dominant Taxa | 49 | 22 | 11 | 6 | 2 | 8 | 333 |
| Threshold | Threshold | 40 | 12 | 10 | 7 | 3 | 8 | 300 |
References
- Pillsbury, R.W.; McGuire, M.A. Factors affecting the distribution of wild rice (Zizania palustris) and the associated macrophyte community. Wetlands 2009, 29, 724–734. [Google Scholar] [CrossRef]
- Drewes, A.D.; Silbernagel, J. Uncovering the spatial dynamics of wild rice lakes, harvesters and management across Great Lakes landscapes for shared regional conservation. Ecol. Model. 2012, 229, 97–107. [Google Scholar] [CrossRef]
- DNR. Natural Wild Rice in Minnesota; Minnesota Department of Natural Resources: St. Paul, MI, USA, 2008; p. 116. Available online: http://files.dnr.state.mn.us/fish_wildlife/wildlife/shallowlakes/natural-wild-rice-in-minnesota.pdf (accessed on 7 August 2019).
- Price, M.W. Spectral Identification of Wild Rice (Zizania Palustris L.) Using Indigenous Knowledge and Landsat Multispectral Data. Master’s Thesis, University of Montana, Missoula, MT, USA, 2012. [Google Scholar]
- Anderson, R.; Kapfer, P.; Ueland, J.; Lawrence, J.; Cordts, S. Comparison of Wild Rice and Waterfowl Surveys. 2011. Available online: https://www.lccmr.leg.mn/projects/2011/finals/2011_04j_2e_rpt_wild-rice-data-waterfowl-surveys.pdf (accessed on 29 June 2020).
- Kennard, W.; Porter, R.; Grombacher, A.; Phillips, R.L. A comparative map of wild rice (Zizania palustris L. 2n = 2x = 30). Theor. Appl. Genet. 1999, 99, 793–799. [Google Scholar] [CrossRef]
- Khoury, C.K.; Greene, S.; Wiersema, J.H.; Maxted, N.; Jarvis, A.; Struik, P.C. An inventory of crop wild relatives of the United States. Crop Sci. 2013, 53, 1496–1508. [Google Scholar] [CrossRef]
- Biesboer, D.D. The ecology and conservation of wild rice, Zizania palustris L., in North America. Acta Limnol. Bras. 2019, 31. [Google Scholar] [CrossRef] [Green Version]
- Rickman, D.L.; Greensky, W.A.; Al-Hamdan, M.Z.; Estes, M.G.; Crosson, W.L.; Estes, S.M. Spatial and Temporal Analyses of Environmental Effects on Zizania palustris and Its Natural Cycles. NASA Technical Reports Server. 2017. Available online: https://ntrs.nasa.gov/search.jsp?R=20170010653 (accessed on 29 June 2020).
- Oelke, A.E.; Teynor, T.M.; Carter, P.R.; Percich, J.A.; Noetzel, D.M.; Bloom, P.R.; Porter, A.R.; Schertz, C.E. Wild rice. In Alternative Field Crops Manual; University of Minnesota: St. Paul, MN, USA, 1997; pp. 315–334. [Google Scholar]
- Rundquist, D.C.; Narumalani, S.; Narayanan, R.M. A review of wetlands remote sensing and defining new considerations. Remote. Sens. Rev. 2001, 20, 207–226. [Google Scholar] [CrossRef]
- Gonoski, J.; Burk, T.E.; Bolstad, P.V.; Balogh, M. Rice Lake National Wildlife Refuge Historic Wild Rice Mapping (1983–2004); College of Natural Resources and Minnesota Agricultural Experiment Station, University of Minnesota: St. Paul, MN, USA, 2005. [Google Scholar]
- Kontgis, C.; Schneider, A.; Ozdogan, M. Mapping rice paddy extent and intensification in the Vietnamese Mekong River Delta with dense time stacks of Landsat data. Remote Sens. Environ. 2015, 169, 255–269. [Google Scholar] [CrossRef]
- Dong, J.; Xiao, X.; Menarguez, M.A.; Zhang, G.; Qin, Y.; Thau, D.; Biradar, C.; Moore, B. Mapping paddy rice planting area in northeastern Asia with Landsat 8 images, phenology-based algorithm and Google Earth Engine. Remote Sens. Environ. 2016, 185, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Torbick, N.; Chowdhury, D.; Salas, W.; Qi, J. Monitoring rice agriculture across Myanmar using time series Sentinel-1 assisted by Landsat-8 and PALSAR-2. Remote Sens. 2017, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Mansaray, L.R.; Huang, W.; Zhang, D.; Huang, J.; Li, J. Mapping rice fields in urban Shanghai, Southeast China, using Sentinel-1A and Landsat 8 datasets. Remote Sens. 2017, 9, 257. [Google Scholar] [CrossRef] [Green Version]
- Gallant, A.L.; Kaya, S.G.; White, L.; Brisco, B.; Roth, M.F.; Sadinski, W.; Rover, J. Detecting emergence, growth, and senescence of wetland vegetation with polarimetric Synthetic Aperture Radar (SAR) data. Water 2014, 6, 694–722. [Google Scholar] [CrossRef] [Green Version]
- Brisco, B.; Li, K.; Tedford, B.; Charbonneau, F.; Yun, S.; Murnaghan, K. Compact polarimetry assessment for rice and wetland mapping. Int. J. Remote Sens. 2012, 34, 1949–1964. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.F.; Son, N.T.; Chen, C.R.; Chang, L.Y.; Chiang, S.H. Rice crop mapping using Sentinel-1A phenological metrics. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, 41, 863–865. [Google Scholar]
- Lasko, K.; Vadrevu, K.P.; Tran, V.T.; Justice, C. Mapping double and single crop paddy rice with Sentinel-1A at varying spatial scales and polarizations in Hanoi, Vietnam. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 498–512. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Costa, M.P.F.; Melack, J.M.; Novo, E.M.L.M. Remote sensing of aquatic vegetation: Theory and applications. Environ. Monit. Assess. 2007, 140, 131–145. [Google Scholar] [CrossRef]
- MNDR. (n.d). Minnesota Climate Trends 1895–2018. Available online: https://arcgis.dnr.state.mn.us/ewr/climatetrends/) (accessed on 29 June 2020).
- Radomski, P.; Woizeschke, K.; Carlson, K.; Perleberg, D. Reproducibility of emergent plant mapping on lakes. N. Am. J. Fish. Manag. 2011, 31, 144–150. [Google Scholar] [CrossRef]
- Perleberg, D.; Radomski, P.; Simon, S.; Carlson, K.; Knopik, J. Minnesota Lake Plant Survey Manual, for Use by MNDNR Fisheries Section and EWR Lake Habitat Program; Minnesota Department of Natural Resources, Ecological and Water Resources Division: Brainerd, MN, USA, 2016; 128p. [Google Scholar]
- United States Department of Agriculture (USDA). Farm Service Agency. National Agriculture Imagery Program (NAIP) [Image collection]. Google Earth Engine API. 2015. Available online: https://code.earthengine.google.com/ (accessed on 20 February 2019).
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- United States Geological Survey (USGS). Earth Resources Observation and Science Center. Landsat 8 OLI Level-2 Surface Reflectance (SR) Science Product. 2014. Available online: https://www.usgs.gov/centers/eros/science/usgs-eros-archive-landsat-archives-landsat-8-olitirs-level-2-data-products?qt-science_center_objects=0#qt-science_center_objects (accessed on 20 February 2019).
- Young, N.E.; Anderson, R.; Chignell, S.M.; Vorster, A.G.; Lawrence, R.; Evangelista, P.H. A survival guide to Landsat preprocessing. Ecology 2017, 98, 920–932. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.H.A.; Zhang, L.; Shuai, T.; Tong, Q. Derivation of a tasselled cap transformation based on Landsat 8 at-satellite reflectance. Remote Sens. Lett. 2014, 5, 423–431. [Google Scholar] [CrossRef]
- Copernicus Sentinel Data (2015). Retrieved from Google Earth Engine [4 February 2019], processed by ESA.
- Breiman, L.; Last, M.; Rice, J. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Genuer, R.; Poggi, J.-M.; Tuleau-Malot, C. VSURF: An R package for variable selection using random forests. R J. 2015, 7, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and regression by randomforest. R News 2002, 2, 18–22. [Google Scholar]
- Shabani, F.; Kumar, L.; Ahmadi, M. Assessing accuracy methods of species distribution models: AUC, specificity, sensitivity and the true skill statistic. Glob. J. Hum. Soc. Sci. B 2018, 18, 7–18. [Google Scholar]
- Kuhn, M. Caret package. J. Stat. Softw. 2008, 28. [Google Scholar]
- Myrbo, A.; Swain, E.B.; Engstrom, D.R.; Wasik, J.C.; Brenner, J.; Shore, M.D.; Peters, E.B.; Blaha, G. Sulfide generated by sulfate reduction is a primary controller of the occurrence of wild rice (Zizania palustris) in shallow aquatic ecosystems. J. Geophys. Res. Biogeosci. 2017, 122, 2736–2753. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Boles, S.; Liu, J.; Zhuang, D.; Frolking, S.; Li, C.; Salas, W.; Moore, B. Mapping paddy rice agriculture in southern China using multi-temporal MODIS images. Remote Sens. Environ. 2005, 95, 480–492. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S.A. Comparison of time series tasseled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]









| Sample Class | Primary Taxa | Secondary Taxa | Associated Taxa | Number of Training Data Points | ||
|---|---|---|---|---|---|---|
| Dominant Vegetation | <Dominant Cover and >30% Cover | <30% Cover | Base Model | Dominant Taxa Model | Threshold Model | |
| Wildrice | • | ◦ | 11 | 11 | 11 | |
| Mixed Wildrice and Others | • | ◦ | ◦ | 322 | 322 | 283 |
| Associated Wildrice | ◦ | •◦ | •◦ | 167 | 0 | 0 |
| Wildrice Absence | ◦ | ◦ | ◦ | 500 | 333 | 300 |
| Open Water | 200 | 200 | 199 | |||
| Model | Validation Set | PCC | Sensitivity (Presence Accuracy) | Specificity (Absence Accuracy) | Confusion Matrices | |||
|---|---|---|---|---|---|---|---|---|
| TP | TN | FP | FN | |||||
| Base | Base | 83.8% | 83.3% | 84.2% | 363 | 523 | 98 | 73 |
| 83.3% | 84.2% | 15.8% | 16.7% | |||||
| Dominant Taxa | Dominant Taxa | 87.5% | 81.6% | 90.8% | 244 | 483 | 49 | 55 |
| 81.6% | 90.8% | 9.2% | 18.4% | |||||
| Threshold | Threshold | 91.1% | 89.5% | 92.0% | 230 | 459 | 40 | 27 |
| 89.5% | 92.0% | 8.0% | 10.5% | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Shea, K.; LaRoe, J.; Vorster, A.; Young, N.; Evangelista, P.; Mayer, T.; Carver, D.; Simonson, E.; Martin, V.; Radomski, P.; et al. Improved Remote Sensing Methods to Detect Northern Wild Rice (Zizania palustris L.). Remote Sens. 2020, 12, 3023. https://doi.org/10.3390/rs12183023
O’Shea K, LaRoe J, Vorster A, Young N, Evangelista P, Mayer T, Carver D, Simonson E, Martin V, Radomski P, et al. Improved Remote Sensing Methods to Detect Northern Wild Rice (Zizania palustris L.). Remote Sensing. 2020; 12(18):3023. https://doi.org/10.3390/rs12183023
Chicago/Turabian StyleO’Shea, Kristen, Jillian LaRoe, Anthony Vorster, Nicholas Young, Paul Evangelista, Timothy Mayer, Daniel Carver, Eli Simonson, Vanesa Martin, Paul Radomski, and et al. 2020. "Improved Remote Sensing Methods to Detect Northern Wild Rice (Zizania palustris L.)" Remote Sensing 12, no. 18: 3023. https://doi.org/10.3390/rs12183023
APA StyleO’Shea, K., LaRoe, J., Vorster, A., Young, N., Evangelista, P., Mayer, T., Carver, D., Simonson, E., Martin, V., Radomski, P., Knopik, J., Kern, A., & Khoury, C. K. (2020). Improved Remote Sensing Methods to Detect Northern Wild Rice (Zizania palustris L.). Remote Sensing, 12(18), 3023. https://doi.org/10.3390/rs12183023