Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Remote Sensing Data
2.2. Climate and Soil Data
2.3. Methods
2.3.1. Data Preprocessing
2.3.2. A Satellite-Based Terrestrial Resilience Index
2.3.3. Regression Analysis
3. Results
3.1. Model Determination and Validation
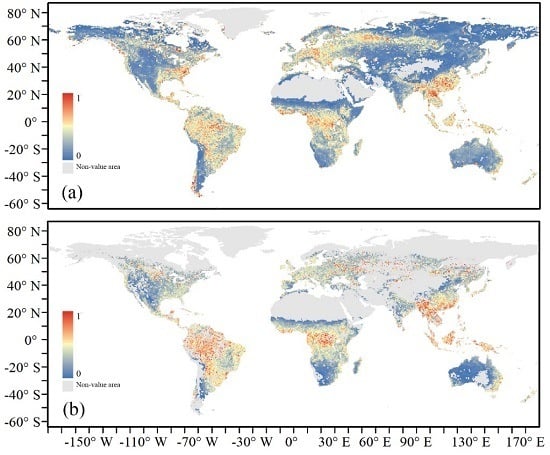
3.2. Spatial Distribution and Temporal Variation of Satellite-Based Terrestrial Resilience
3.3. Driving Factors of Satellite-Based Terrestrial Resilience
4. Discussion
4.1. Satellite-Based Terrestrial Resilience
4.2. Distribution and Variation of Satellite-Based Terrestrial Resilience
4.3. Limitations and Recommendations for Future Research
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tittensor, D.P.; Walpole, M.; Hill, S.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.; Leadley, P.W.; Regan, E.C.; Alkemade, R. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.N.; Newbold, T.; Contu, S.; Hill, S.L.L.; Lysenko, I.; De Palma, A.; Phillips, H.R.P.; Senior, R.A.; Bennett, D.J.; Booth, H.; et al. The predicts database: A global database of how local terrestrial biodiversity responds to human impacts. Ecol. Evol. 2014, 4, 4701–4735. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency (EPA). Climate Change Indicators in the United States; U.S. Environmental Protection Agency (EPA): Washington, DC, USA, 2010. [Google Scholar]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S. Resilience and stability of ecological systems. Ann. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Pimm, S.L. The complexity and stability of ecosystems. Nature 1984, 307, 321–326. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R.; Dakos, V.; Van Nes, E.H. Generic indicators of ecological resilience: Inferring the chance of a critical transition. Ann. Rev. Ecol. Evol. Syst. 2015, 46, 145–167. [Google Scholar] [CrossRef]
- Hirota, M.; Holmgren, M.; Van Nes, E.H.; Scheffer, M. Global resilience of tropical forest and savanna to critical transitions. Science 2011, 334, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.H.; Abel, N.; Anderies, J.M.; Ryan, P. Resilience, adaptability, and transformability in the goulburn-broken catchment, australia. Ecol. Soc. 2009, 14, 12. [Google Scholar] [CrossRef]
- Westman, W.E.; O’Leary, J.F. Measures of resilience: The response of coastal sage scrub to fire. Vegetatio 1986, 65, 179–189. [Google Scholar] [CrossRef]
- Wang, R.; Dearing, J.A.; Langdon, P.G.; Zhang, E.; Yang, X.; Dakos, V.; Scheffer, M. Flickering gives early warning signals of a critical transition to a eutrophic lake state. Nature 2012, 492, 419. [Google Scholar] [CrossRef] [PubMed]
- MacGillivray, C.; Grime, J.; Team, T.I.S.P. Testing predictions of the resistance and resilience of vegetation subjected to extreme events. Funct. Ecol. 1995, 9, 640–649. [Google Scholar] [CrossRef]
- Wu, J.; Liang, S. Developing an integrated remote sensing based biodiversity index for predicting animal species richness. Remote Sens. 2018, 10, 739. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Wang, J.; Yu, L.; Zhao, Y.; Zhao, Y.; Liang, L.; Niu, Z.; Huang, X.; Fu, H.; Liu, S. Finer resolution observation and monitoring of global land cover: First mapping results with landsat tm and etm+ data. Int. J. Remote Sens. 2013, 34, 2607–2654. [Google Scholar] [CrossRef] [Green Version]
- Huete, A. Ecology: Vegetation’s responses to climate variability. Nature 2016, 531, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Bisson, M.; Fornaciai, A.; Coli, A.; Mazzarini, F.; Pareschi, M.T. The vegetation resilience after fire (vraf) index: Development, implementation and an illustration from central italy. Int. J. Appl. Earth Observ. Geoinform. 2008, 10, 312–329. [Google Scholar] [CrossRef]
- Mauro, B.D.; Fava, F.; Busetto, L.; Crosta, G.F.; Colombo, R. Post-fire resilience in the alpine region estimated from modis satellite multispectral data. Int. J. Appl. Earth Observ. Geoinform. 2014, 32, 163–172. [Google Scholar] [CrossRef]
- Díaz-Delgado, R.; Lloret, F.; Pons, X.; Terradas, J. Satellite evidence of decreasing resilience in mediterranean plant communities after recurrent wildfires. Ecology 2002, 83, 2293–2303. [Google Scholar] [CrossRef] [Green Version]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef]
- Van Nes, E.H.; Holmgren, M.; Hirota, M.; Scheffer, M. Response to comment on “global resilience of tropical forest and savanna to critical transitions”. Science 2012, 336, 541–541. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, Z.; Nippert, J.B. Comment on “global resilience of tropical forest and savanna to critical transitions”. Science 2012, 336, 541–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Keersmaecker, W.; Lhermitte, S.; Tits, L.; Honnay, O.; Somers, B.; Coppin, P. A model quantifying global vegetation resistance and resilience to short-term climate anomalies and their relationship with vegetation cover. Global Ecol. Biogeogr. 2015, 24, 539–548. [Google Scholar] [CrossRef]
- Harris, A.; Carr, A.; Dash, J. Remote sensing of vegetation cover dynamics and resilience across southern africa. Int. J. Appl. Earth Observ. Geoinform. 2014, 28, 131–139. [Google Scholar] [CrossRef]
- Seddon, A.W.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbesselt, J.; Umlauf, N.; Hirota, M.; Holmgren, M.; Van Nes, E.H.; Herold, M.; Zeileis, A.; Scheffer, M. Remotely sensed resilience of tropical forests. Nat. Clim. Change 2016, 6, 1028–1031. [Google Scholar] [CrossRef]
- Holling, C.S. The resilience of terrestrial ecosystems: Local surprise and global change. Sustain. Dev. Biosph. 1986, 14, 292–317. [Google Scholar]
- Pimm, S.L. Complexity and stability: Another look at macarthur’s original hypothesis. Oikos 1979, 351–357. [Google Scholar] [CrossRef]
- Costanza, R. Ecosystem health and ecological engineering. Ecol. Eng. 2012, 45, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Liang, S.; Wang, J.; Xiang, Y.; Zhao, X.; Song, J. Long-time-series global land surface satellite leaf area index product derived from modis and avhrr surface reflectance. IEEE Trans. Geosci. Remote Sens. 2016, 54, 5301–5318. [Google Scholar] [CrossRef]
- Liang, S.; Zhao, X.; Liu, S.; Yuan, W.; Cheng, X.; Xiao, Z.; Zhang, X.; Liu, Q.; Cheng, J.; Tang, H. A long-term global land surface satellite (glass) data-set for environmental studies. Int. J. Digit. Earth 2013, 6, 5–33. [Google Scholar] [CrossRef]
- Xiao, Z.; Liang, S.; Jiang, B. Evaluation of four long time-series global leaf area index products. Agricult. Forest Meteorol. 2017, 246, 218–230. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the earth and its drivers. Nat. Clim. Change 2016, 6, 791. [Google Scholar] [CrossRef]
- Liang, S.; Yi, Q.; Liu, J. Vegetation dynamics and responses to recent climate change in xinjiang using leaf area index as an indicator. Ecol. Indic. 2015, 58, 64–76. [Google Scholar]
- Kim, K.; Wang, M.-c.; Ranjitkar, S.; Liu, S.-h.; Xu, J.-c.; Zomer, R.J. Using leaf area index (lai) to assess vegetation response to drought in yunnan province of china. J. Mount. Sci. 2017, 14, 1863–1872. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (lai) 3g and fraction of photosynthetically active radiation (fpar) 3g derived from global inventory modeling and mapping studies (gimms) normalized difference vegetation index (ndvi3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Page, Y.L.; Pereira, J.; Trigo, R.; Camara, C.d.; Oom, D.; Mota, B. Global fire activity patterns (1996–2006) and climatic influence: An analysis using the world fire atlas. Atmosph. Chem. Phys. 2008, 8, 1911–1924. [Google Scholar] [CrossRef] [Green Version]
- Arino, O.; Casadio, S.; Serpe, D. Global night-time fire season timing and fire count trends using the atsr instrument series. Remote Sens. Environ. 2012, 116, 226–238. [Google Scholar] [CrossRef]
- Friedl, M.; Sulla-Menashe, D. MCD12Q1 MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V006. 2019, distributed by NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/MODIS/MCD12Q1.006 (accessed on 4 February 2020).
- Friedl, M.A.; McIver, D.K.; Hodges, J.C.; Zhang, X.; Muchoney, D.; Strahler, A.H.; Woodcock, C.E.; Gopal, S.; Schneider, A.; Cooper, A. Global land cover mapping from modis: Algorithms and early results. Remote Sens. Environ. 2002, 83, 287–302. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag effects of global vegetation responses to climate change. Global Change Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Schrier, G.; Barichivich, J.; Briffa, K.; Jones, P. A scpdsi-based global data set of dry and wet spells for 1901–2009. J. Geophys. Res. Atmosph. 2013, 118, 4025–4048. [Google Scholar] [CrossRef]
- Wells, N.; Goddard, S.; Hayes, M.J. A self-calibrating palmer drought severity index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Change 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.; Osborn, T.; Lister, D. Updated high-resolution grids of monthly climatic observations–the cru ts3. 10 dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- New, M.; Hulme, M.; Jones, P. Representing twentieth-century space–time climate variability. Part ii: Development of 1901–96 monthly grids of terrestrial surface climate. J. Clim. 2000, 13, 2217–2238. [Google Scholar] [CrossRef]
- Mitchell, T.D.; Jones, P.D. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int. J. Climatol. 2005, 25, 693–712. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Ciais, P.; Li, J.; Friedlingstein, P.; Koven, C.; Chen, A. Spring temperature change and its implication in the change of vegetation growth in north america from 1982 to 2006. Proc. Natl. Acad. Sci. USA 2011, 108, 1240–1245. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Lister, D.; Osborn, T.; Harpham, C.; Salmon, M.; Morice, C. Hemispheric and large-scale land-surface air temperature variations: An extensive revision and an update to 2010. J. Geophys. Res. Atmosph. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- Jong, R.; Schaepman, M.E.; Furrer, R.; Bruin, S.; Verburg, P.H. Spatial relationship between climatologies and changes in global vegetation activity. Glob. Change Biol. 2013, 19, 1953–1964. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Penuelas, J.; Zhang, G. Asymmetric effects of daytime and night-time warming on northern hemisphere vegetation. Nature 2013, 501, 88. [Google Scholar] [CrossRef]
- Beinroth, F.H.; Eswaran, H.; Reich, P.F. Global assessment of land quality. Sustain. Global Farm 2001, 569–574. Available online: https://topsoil.nserl.purdue.edu/nserlweb-old/isco99/pdf/ISCOdisc/SustainingTheGlobalFarm/P233-Beinroth.pdf (accessed on 30 January 2020).
- Hild, S.; Chelkowski, S.; Freise, A. Pushing towards the et sensitivity using ‘conventional’ technology. arXiv 2008, preprint. arXiv:0810.0604. [Google Scholar]
- Mageau, M.T.; Costanza, R.; Ulanowicz, R.E. The development and initial testing of a quantitative assessment of ecosystem health. Health 1995, 1, 201–213. [Google Scholar]
- Mageau, M.T. Development and testing of a quantitative assessment of ecosystem health. Ecosyst. Health 1998, 1, 201–213. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with r; Chapman & Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Fahrmeir, L.; Kneib, T.; Lang, S.; Marx, B. Regression: Models, Methods and Applications; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Hastie, T.J.; Tibshirani, R.J. Generalized Additive Models; Chapman & Hall/CRC: Boca Raton, FL, USA, 1990. [Google Scholar]
- Washington-Allen, R.A.; Ramsey, R.; West, N.E.; Norton, B.E. Quantification of the ecological resilience of drylands using digital remote sensing. Ecol. Soc. 2008, 13, 33. [Google Scholar] [CrossRef] [Green Version]
- Pimm, S.L. The Balance of Nature? Ecological Issues in the Conservation of Species and Communities; University of Chicago Press: Chicago, IL, USA, 1991. [Google Scholar]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological resilience, biodiversity, and scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C. Regime shifts, resilience, and biodiversity in ecosystem management. Ann. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Cumming, G.S. Spatial resilience: Integrating landscape ecology, resilience, and sustainability. Landsc. Ecol. 2011, 26, 899–909. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [Green Version]
- Lanfredi, M.; Simoniello, T.; Macchiato, M. Temporal persistence in vegetation cover changes observed from satellite: Development of an estimation procedure in the test site of the mediterranean italy. Remote Sens. Environ. 2004, 93, 565–576. [Google Scholar]
- Baho, D.; Allen, C.; Garmestani, A.; Fried-Petersen, H.; Renes, S.; Gunderson, L.; Angeler, D. A quantitative framework for assessing ecological resilience. Ecol. Soc. 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Standish, R.J.; Hobbs, R.J.; Mayfield, M.M.; Bestelmeyer, B.T.; Suding, K.N.; Battaglia, L.L.; Eviner, V.; Hawkes, C.V.; Temperton, V.M.; Cramer, V.A. Resilience in ecology: Abstraction, distraction, or where the action is? Biol. Conserv. 2014, 177, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Deffuant, G.; Gilbert, N. Viability and Resilience of Complex Systems: Concepts, Methods and Case Studies from Ecology and Society; Springer Science & Business Media: Berlin, Germany, 2011. [Google Scholar]
- Cantarello, E.; Newton, A.C.; Martin, P.A.; Evans, P.M.; Gosal, A.; Lucash, M.S. Quantifying resilience of multiple ecosystem services and biodiversity in a temperate forest landscape. Ecol. Evolut. 2017, 7, 9661–9675. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Li, X.; Wang, J. Advanced Remote Sensing: Terrestrial Information Extraction and Applications; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Keersmaecker, W.; Lhermitte, S.; Honnay, O.; Farifteh, J.; Somers, B.; Coppin, P. How to measure ecosystem stability? An evaluation of the reliability of stability metrics based on remote sensing time series across the major global ecosystems. Glob. Change Biol. 2014, 20, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Rammig, A.; Mahecha, M.D. Ecology: Ecosystem responses to climate extremes. Nature 2015, 527, 315–316. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Liu, S.; Yu, G.; Bonnefond, J.; Chen, J.; Davis, K.J.; Desai, A.R.; Goldstein, A.H.; Gianelle, D.; Rossi, F. Global estimates of evapotranspiration and gross primary production based on modis and global meteorology data. Remote Sens. Environ. 2010, 114, 1416–1431. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, S.; Sun, G. Forest biomass mapping of northeastern china using glas and modis data. Ieee J.-Stars 2014, 7, 140–152. [Google Scholar]
- Zhao, M.; Heinsch, F.A.; Nemani, R.R.; Running, S.W. Improvements of the modis terrestrial gross and net primary production global data set. Remote Sens. Environ. 2005, 95, 164–176. [Google Scholar]
- Gunderson, L.H. Ecological resilience--in theory and application. Ann. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Campos, G.E.; Moran, M.S.; Huete, A.; Zhang, Y.; Bresloff, C.; Huxman, T.E.; Eamus, D.; Bosch, D.D.; Buda, A.R.; Gunter, S.A. Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature 2013, 494, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Côté, I.M.; Darling, E.S. Rethinking ecosystem resilience in the face of climate change. PLoS Biol. 2010, 8, e1000438. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Coops, N.C.; Beck, P.S.; Chambers, J.Q.; Gangodagamage, C.; Hicke, J.A.; Huang, C.-y.; Kennedy, R.; Krofcheck, D.J.; Litvak, M. Global satellite monitoring of climate-induced vegetation disturbances. Trends Plant Sci. 2015, 20, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragão, L.E.; Anderson, L.O.; Myneni, R.B.; Nemani, R. Persistent effects of a severe drought on amazonian forest canopy. Proc. Natl. Acad. Sci. USA 2013, 110, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.A.; Tulloch, D. Rainfall in the wet-dry tropics: Extreme events at darwin and similarities between years during the period 1870–1983 inclusive. Aust. Ecol. 1985, 10, 281–295. [Google Scholar] [CrossRef]
- O’Grady, A.; Eamus, D.; Hutley, L. Transpiration increases during the dry season: Patterns of tree water use in eucalypt open-forests of northern australia. Tree Physiol. 1999, 19, 591–597. [Google Scholar] [CrossRef]
- Dakos, V.; Van Nes, E.H.; D’Odorico, P.; Scheffer, M. Robustness of variance and autocorrelation as indicators of critical slowing down. Ecology 2012, 93, 264–271. [Google Scholar] [CrossRef] [Green Version]
- DeAngelis, D. Dynamics of Nutrient Cycling and Food Webs; Springer Science & Business Media: Berlin, Germany, 2012; Volume 9. [Google Scholar]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.A.; Eigenbrod, F.; Freckleton, R.P.; Hector, A.L.; Orme, C.D.L.; Petchey, O.L. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evolut. 2015, 30, 673. [Google Scholar] [CrossRef] [Green Version]
- Buma, B.; Wessman, C.A. Disturbance interactions can impact resilience mechanisms of forests. Ecosphere 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Adler, R.F.; Gu, G.; Sapiano, M.R.P.; Wang, J.; Huffman, G.J. Global precipitation: Means, variations and trends during the satellite era (1979–2014). Surveys Geophys. 2017, 38, 679–699. [Google Scholar] [CrossRef] [Green Version]
- Gu, G.; Adler, R.F. Precipitation and temperature variations on the interannual time scale: Assessing the impact of enso and volcanic eruptions. J. Clim. 2011, 24, 2258–2270. [Google Scholar] [CrossRef] [Green Version]
- Costanza, R.; Mageau, M. What is a healthy ecosystem? Aquat. Ecol. 1999, 33, 105–115. [Google Scholar] [CrossRef]
- Turner, W. Sensing biodiversity. Science 2014, 346, 301–302. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Source | Base Period | Spatial Resolution | Temporal Resolution |
|---|---|---|---|---|
| Leaf Area Index | GLASS AVHRR LAI | 1982–2016 | 0.05° | 8 days |
| GIMMS AVHRR LAI 3g | 1982–2011 | 1/12° | 15 days | |
| Fire | ATSR World Fire Atlas | 2000–2009 | 0.05° | monthly |
| Drought | sc-PDSI | 2000–2009 | 0.5° | monthly |
| Land Cover | MODIS MCD12Q1 | 2001-2012 | 1/12° | yearly |
| Precipitation | CRU TS4.01 Precipitation | 1982–2011 | 0.5° | monthly |
| Temperature | CRU TS4.01 Mean Temperature | 1982–2011 | 0.5° | monthly |
| Soil moisture | USDA-NRCS | 1997 | 1/30° | yearly |
| Name | Coordinate | Disturbance Time | Disturbance Type |
|---|---|---|---|
| Yellowstone national park | 44°45′N; 110°15′W | 1988, 1994, 2008 | Drought |
| IIha do bananal | 9°45′S; 49°45′W | 1994, 1995 (fire) | Drought, Fire |
| Abumonbazi reserve | 3°15′N; 21°45′E | 1986, 1994 | Drought |
| Three river headwater | 33°45′N; 94°15′E | 1995, 2007 | Drought |
| Khan khentii national park | 46°45′N; 119°45′E | 1992, 1993, 2007 | Drought |
| Southern tanami reserve | 21°15′S; 129°45′E | 1988, 2005, 1995 | Drought |
| Indigenous protected area | 0°45′S; 65°15′W | 1982, 1991, 2005 | Drought |
| Kenozersky national park | 61°45′N; 38°15′E | 1992, 2008 | Drought |
| Wabakimi provincial park | 50°45′N; 89°45′W | 1999, 2004, 2009 | Drought |
| defensores del chaco national park | 20°15′S; 60°15′W | 1993, 2007, | Drought |
| Turgaiskiy national park | 48°45′N; 61°45′E | 1994, 2001, 2006, 2008 | Drought |
| Quetico provinvial park | 48°15′N; 91°15′W | 1988, 2007 | Drought |
| Baishuijiang nature reserve | 32°45′N; 104°45′E | 1989, 2006 | Drought |
| Gemsbok national park | 24°45′S; 21°15′E | 1985, 2007 | Drought |
| Upenba national park | 8°45′S; 26°45′E | 1994, 1999 | Drought |
| Mavinga national park | 16°15′S; 21°15′E | 2005 | Drought |
| Witjira national park | 26°15′S; 135°45′E | 1994 | Drought |
| Lorentz national park | 4°15′S; 137°45′E | 2003 | Drought |
| Angoran protected area | 36°45′N; 47°30′E | 1997, 1999, 2008 | Drought |
| Lower rio grande valley | 26°30′N; 98°45′W | 2011 | Drought |
| Floresta national park | 2°45′N; 61°45′W | 1997, 2003 | Fire |
| Laguna del Tigre national | 17°45′N; 90°45′W | 1998, 2003, 2005 | Fire |
| Okefenokee national wildlife refuge | 30°15′N; 82°15′W | 2007 | Fire |
| Machaquila national wildlife refuge | 16°15′N; 89°45′W | 1998, 2000 | Fire |
| Land-Cover Type | Beta Coefficient (for GLASS LAI) | Beta Coefficient (for GIMMS LAI 3g) |
|---|---|---|
| Barren or sparsely vegetated | 0.4 | 0.35 |
| Cropland and natural vegetation | 0.5 | 0.4 |
| Cropland | 0.45 | 0.4 |
| Permanent wetlands | 0.45 | 0.4 |
| Grassland | 0.35 | 0.3 |
| Savannas | 0.35 | 0.3 |
| Woody savannas | 0.35 | 0.3 |
| Open shrublands | 0.55 | 0.5 |
| Closed shrubland | 0.55 | 0.5 |
| Mixed forest | 0.6 | 0.55 |
| Deciduous broadleaf forest | 0.6 | 0.55 |
| Deciduous needleleaf forest | 0.6 | 0.55 |
| Evergreen broadleaf forest | 0.6 | 0.55 |
| Evergreen needleleaf forest | 0.6 | 0.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Liang, S. Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index. Remote Sens. 2020, 12, 595. https://doi.org/10.3390/rs12040595
Wu J, Liang S. Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index. Remote Sensing. 2020; 12(4):595. https://doi.org/10.3390/rs12040595
Chicago/Turabian StyleWu, Jinhui, and Shunlin Liang. 2020. "Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index" Remote Sensing 12, no. 4: 595. https://doi.org/10.3390/rs12040595
APA StyleWu, J., & Liang, S. (2020). Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index. Remote Sensing, 12(4), 595. https://doi.org/10.3390/rs12040595