
Testing the Height Variation Hypothesis with the R rasterdiv Package for Tree Species Diversity Estimation


 , and
, and 
Abstract
:1. Introduction
- test different LiDAR metrics for the assessment of the HH;
- understand which R rasterdiv index is the most accurate in characterizing the HH;
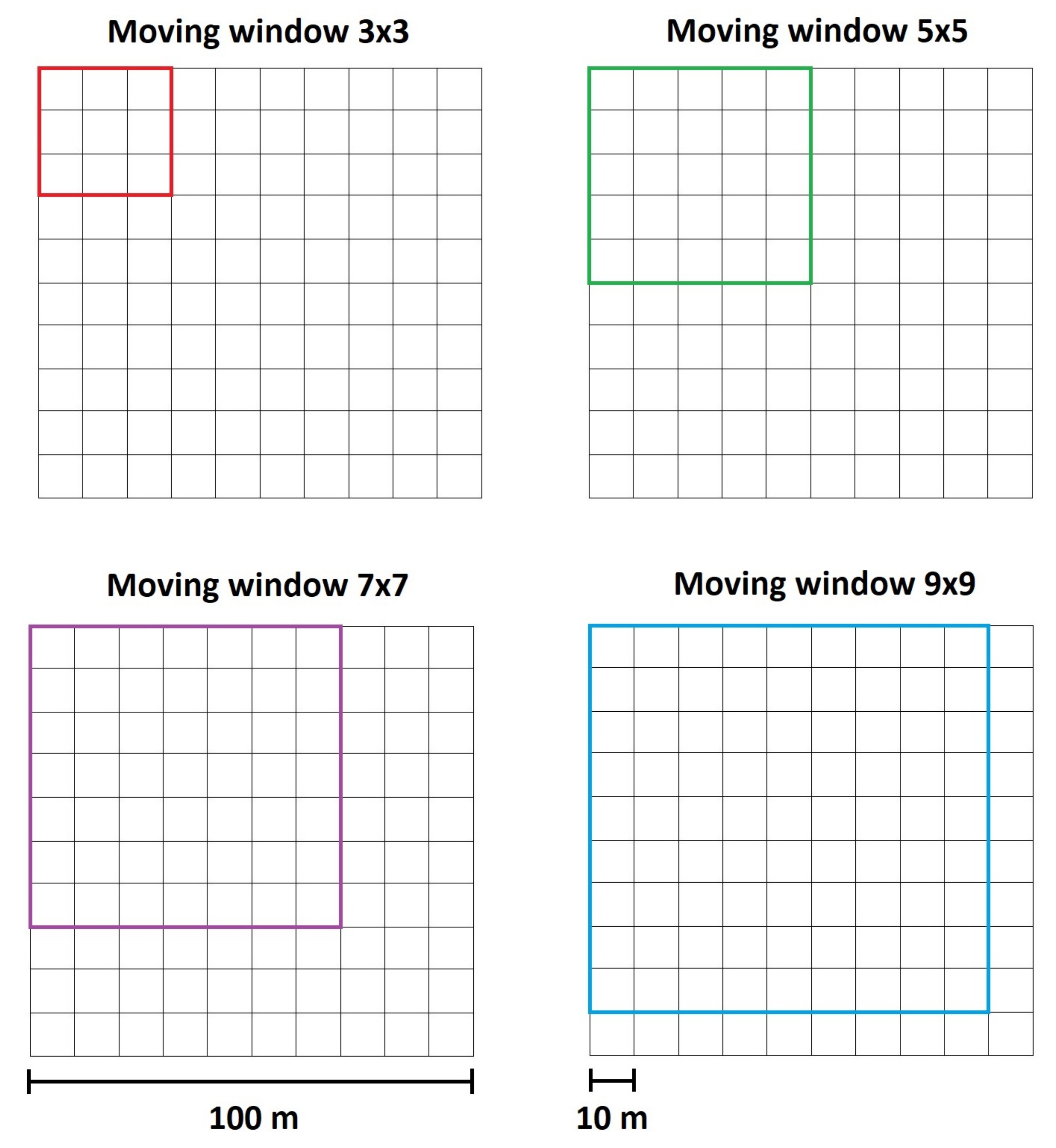
- test the effects of different MW size for each index.
2. Materials and Methods
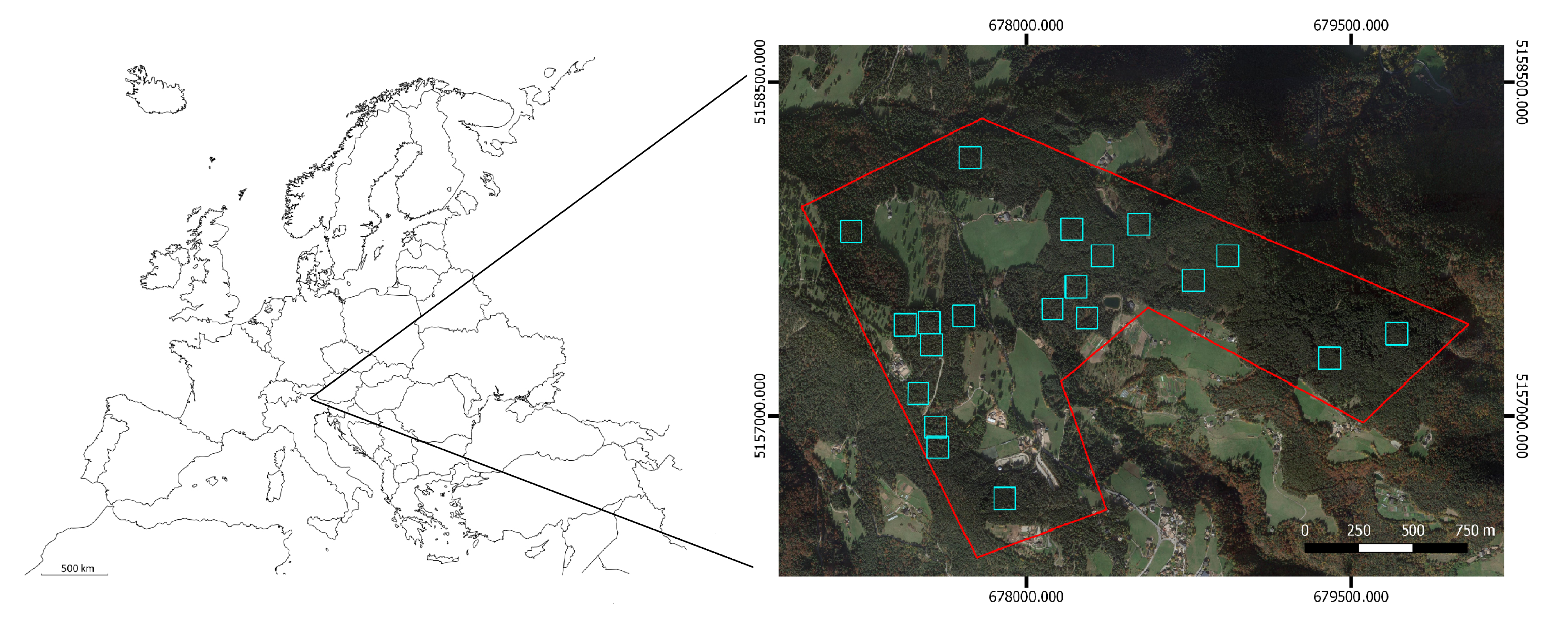
2.1. Study Area and Field Data
- H = Shannon’s H index.
- R = number of species.
- = proportion of species i relative to the total number of species.
2.2. LiDAR Data
- zentropy: entropy of height distribution;
- zmax: maximum height (in meters);
- zsd: standard deviation of height distribution;
- zskew: skewness of height distribution;
- zkurt: kurtosis of height distribution;
- pzabovezmean: percentage of returns above zmean;
- pzabove2: percentage of returns above 2 m;
- zpcumx: cumulative percentage of return in the xth layer according to Woods et al. [51];
- zqx: percentile (quantile) of height distribution.
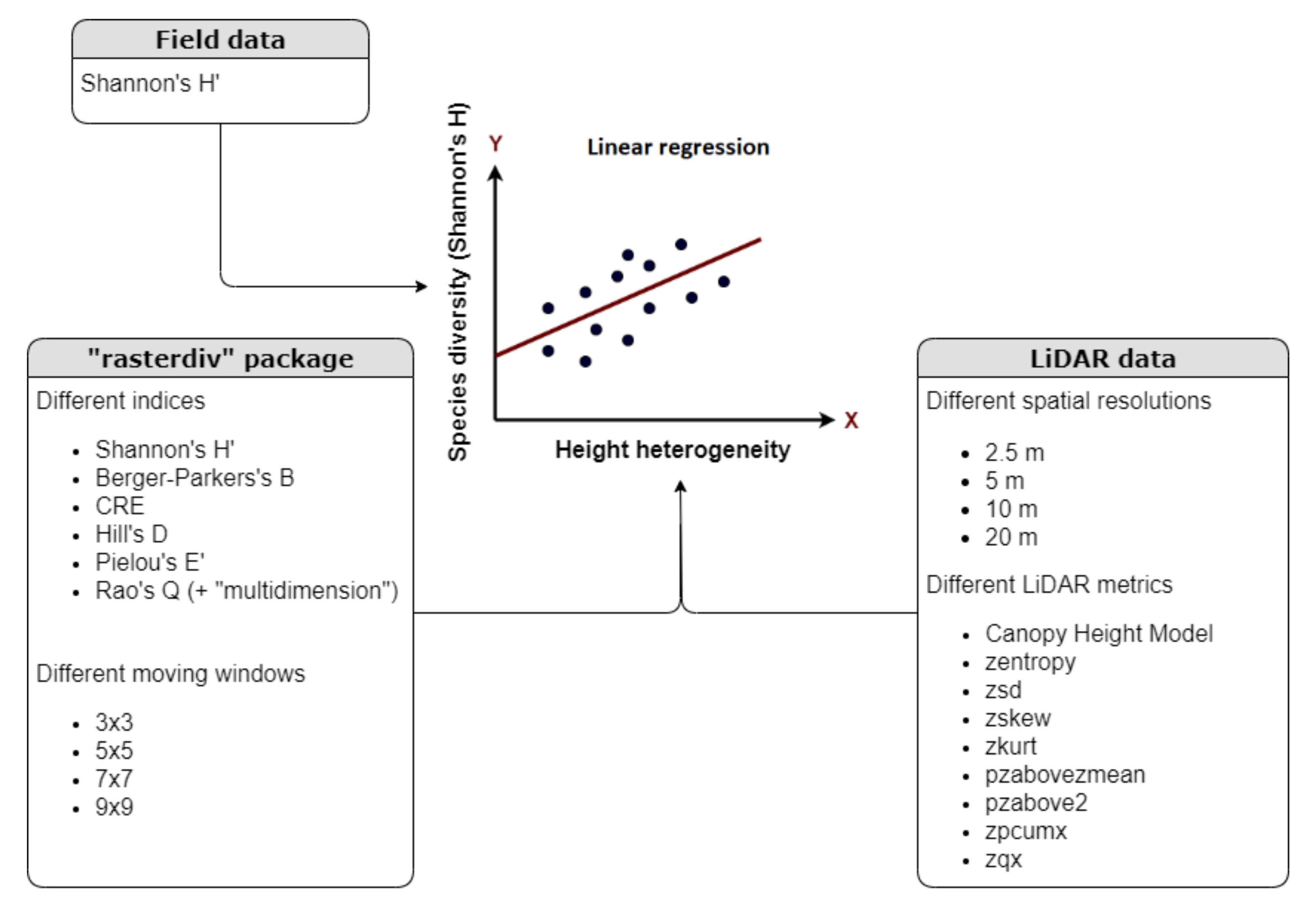
2.3. Height Heterogeneity Assessment and Statistical Analysis
3. Results
3.1. In-Situ Field Data Tree Species Diversity
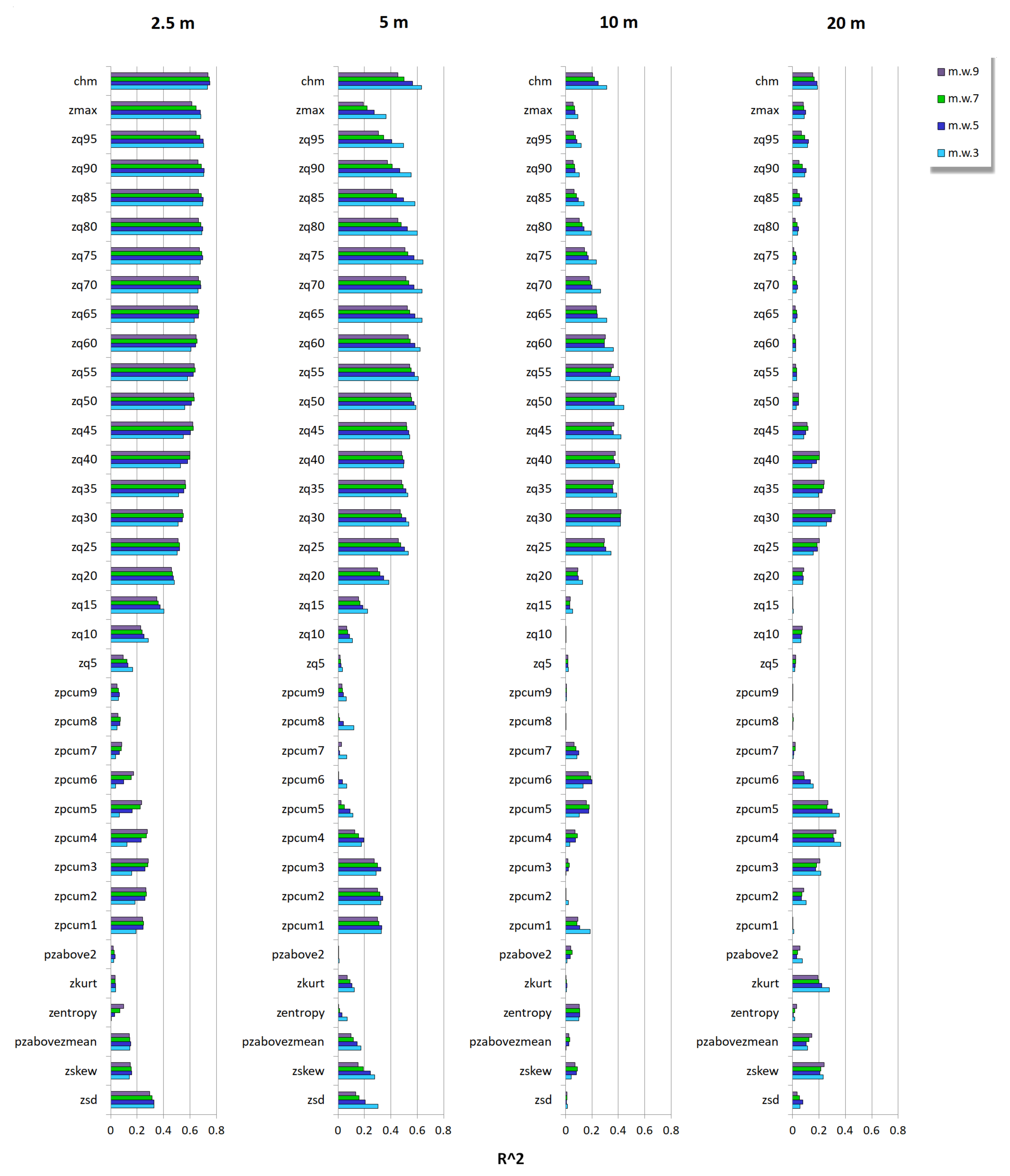
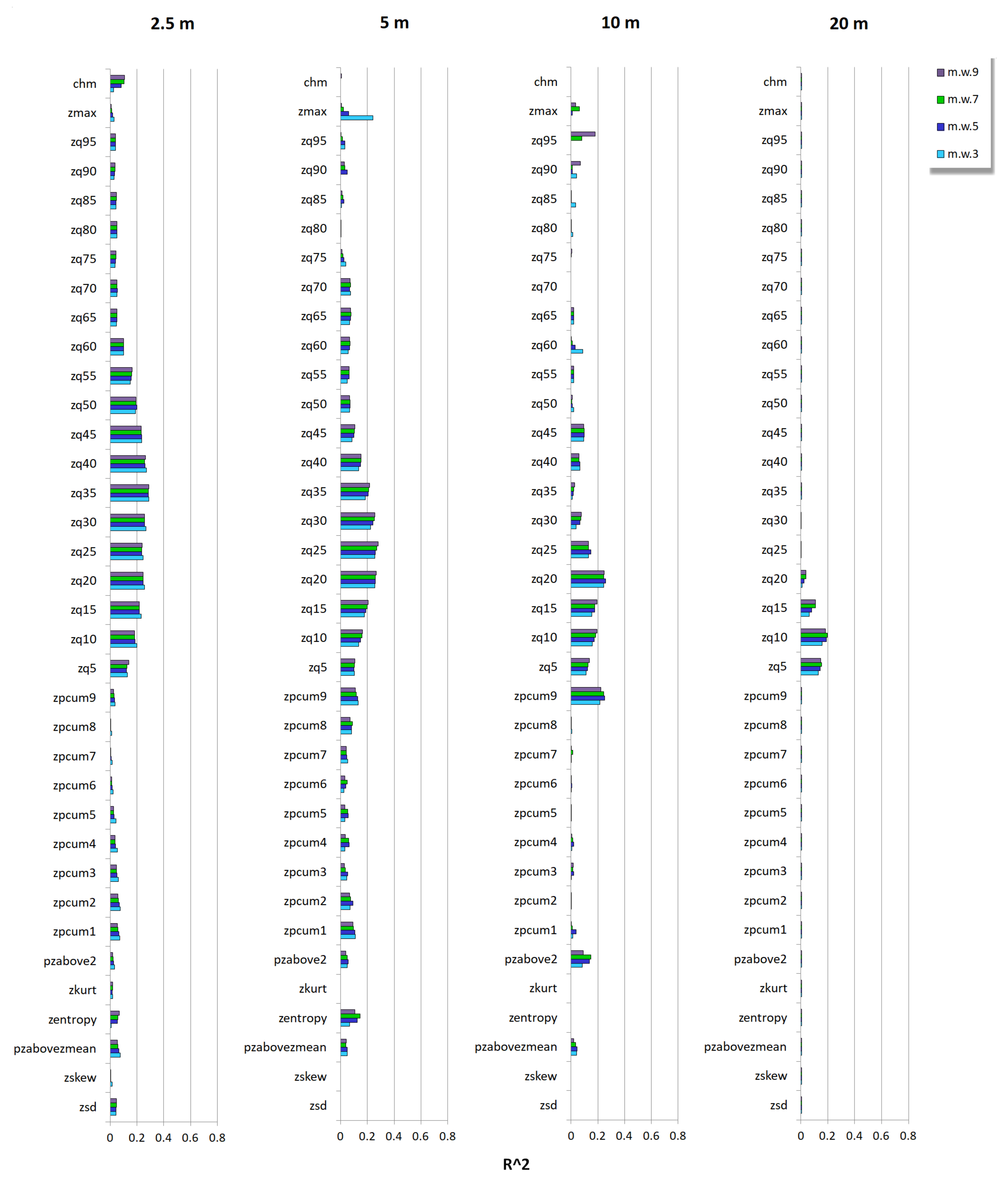
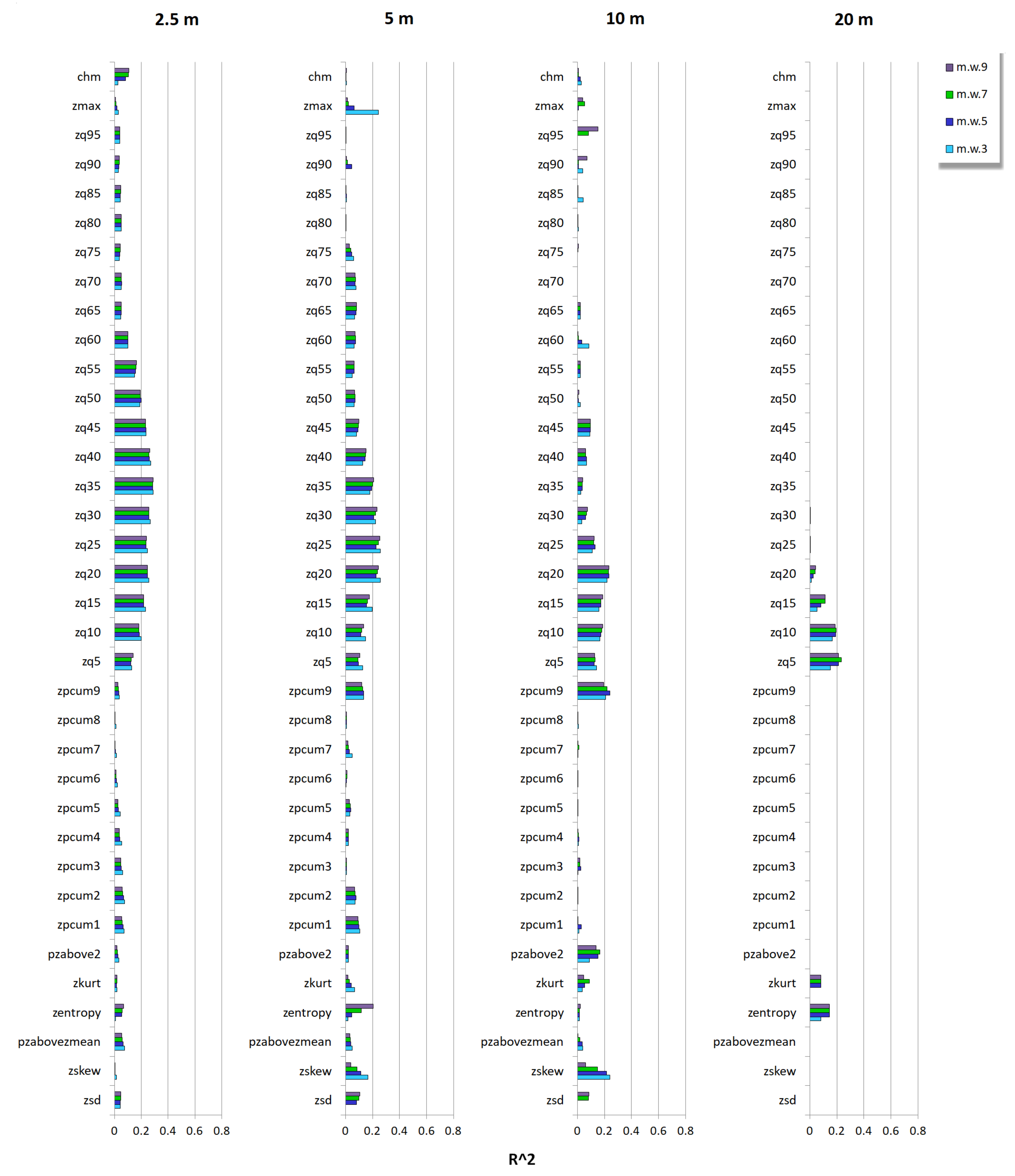
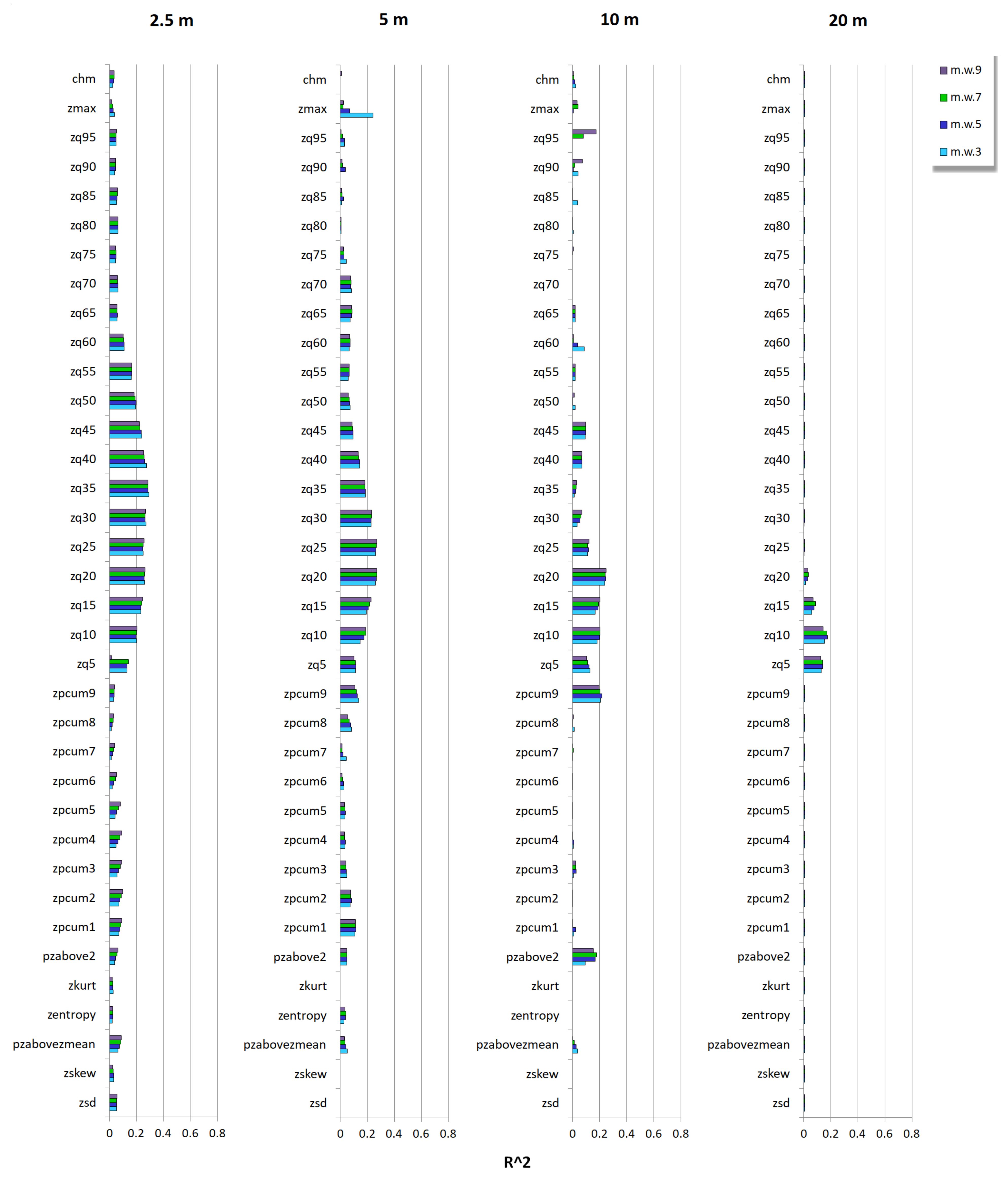
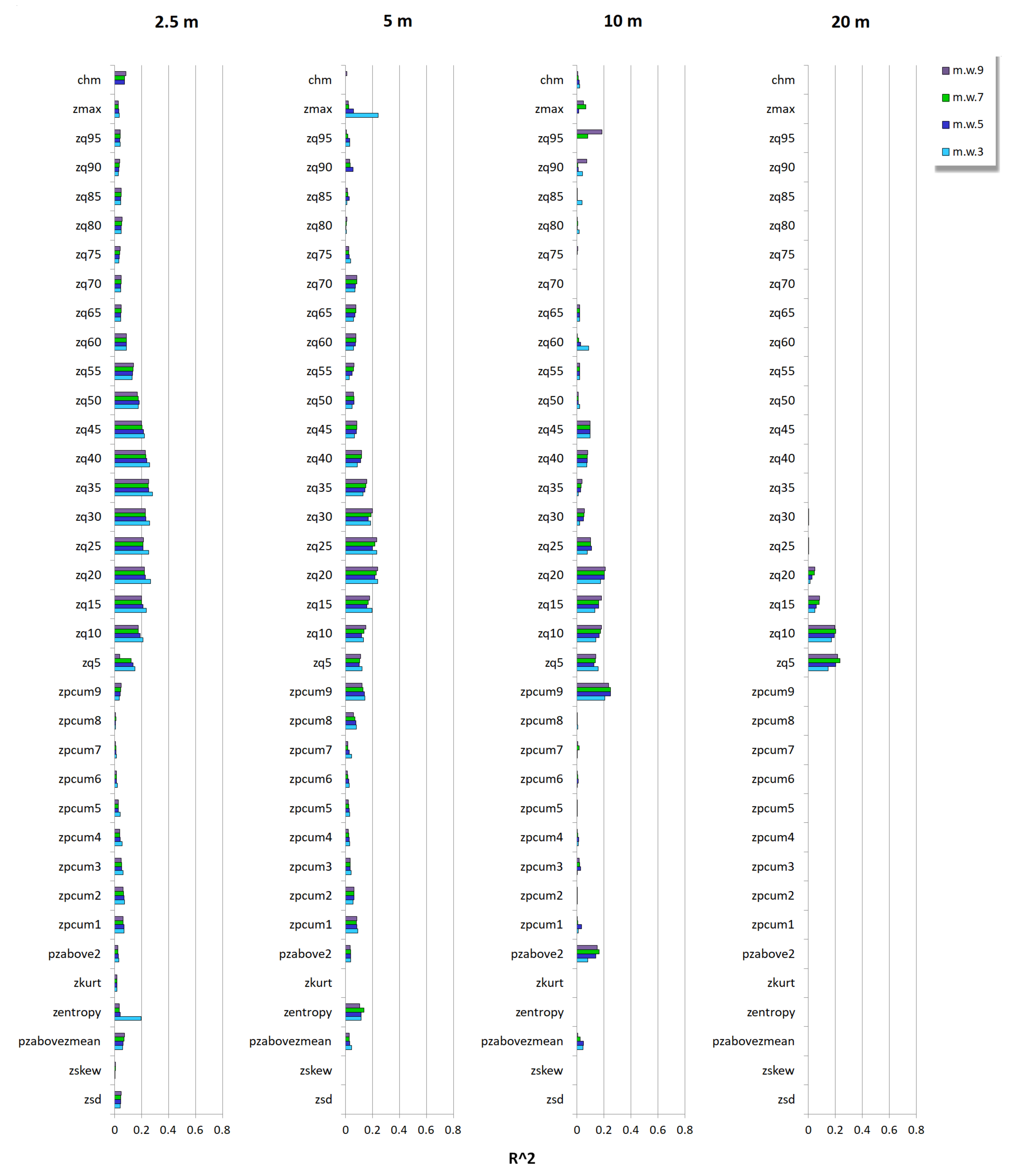
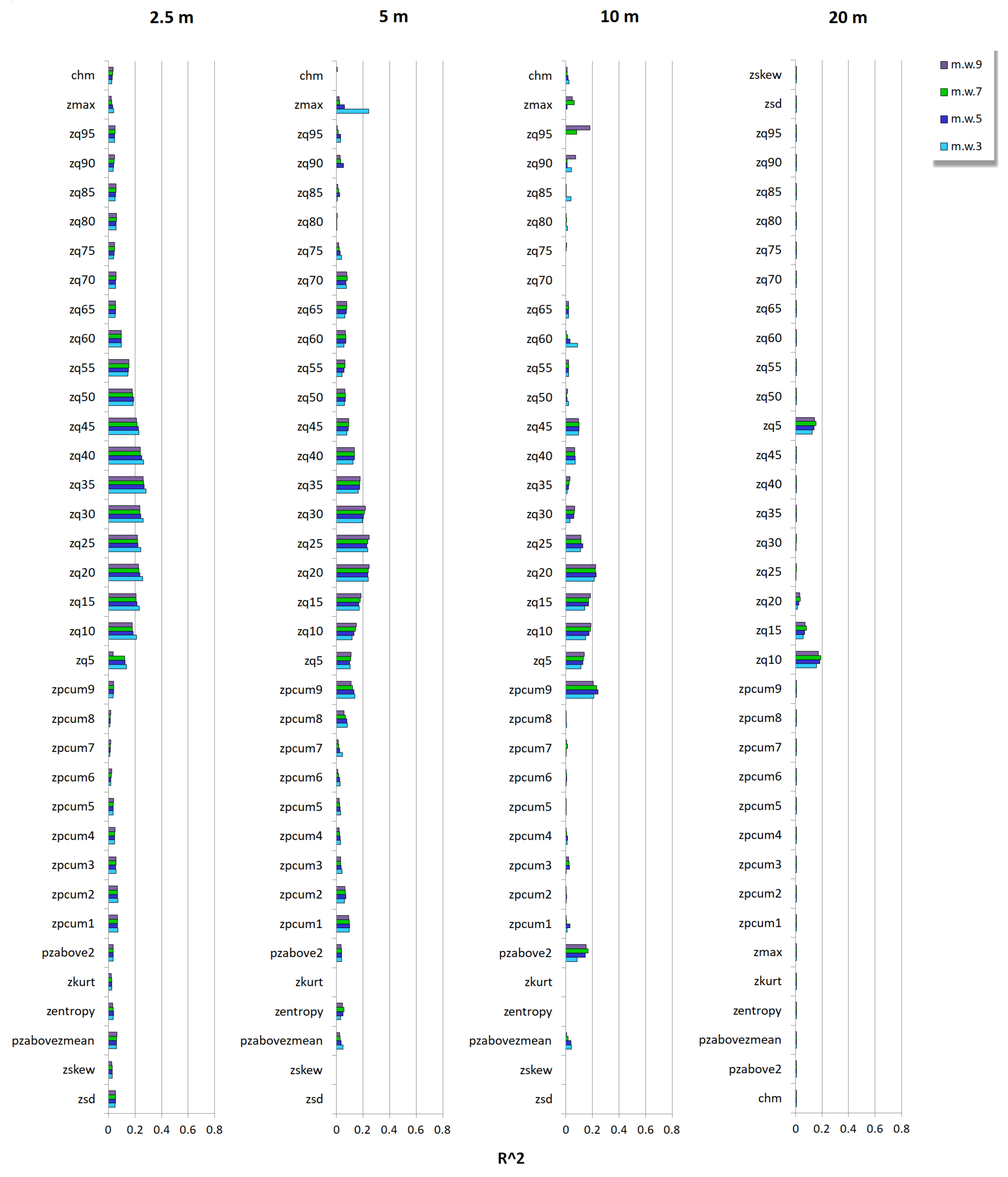
3.2. Relationship between Tree Species Diversity and LiDAR Height Heterogeneity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HH | Height Heterogeneity |
| HVH | Height Variation Hypothesis |
| SVH | Spectral Variation Hypothesis |
| MW | Moving Window |
| CHM | Canopy Height Model |
| DTM | Digital Terrain Model |
| DSM | Digital Surface Model |
| ALS | Airborne Laser Scanning |
| TLS | Terrestrial Laser Scanning |
| TIN | Triangular Irregular Network |
| DBH | Diameter at Breast High |
| zentropy | Entropy of height distribution |
| zmax | Maximum height (in meters) |
| zsd | Standard deviation of height distribution |
| zskew | Skewness of height distribution |
| zkurt | Kurtosis of height distribution |
| pzabovezmean | Percentage of returns above zmean |
| pzabove2 | Percentage of returns above 2 m |
| zpcumx | Cumulative percentage of return in the xth layer according to Woods et al. [51] |
| zqx | Percentile (quantile) of height distribution |
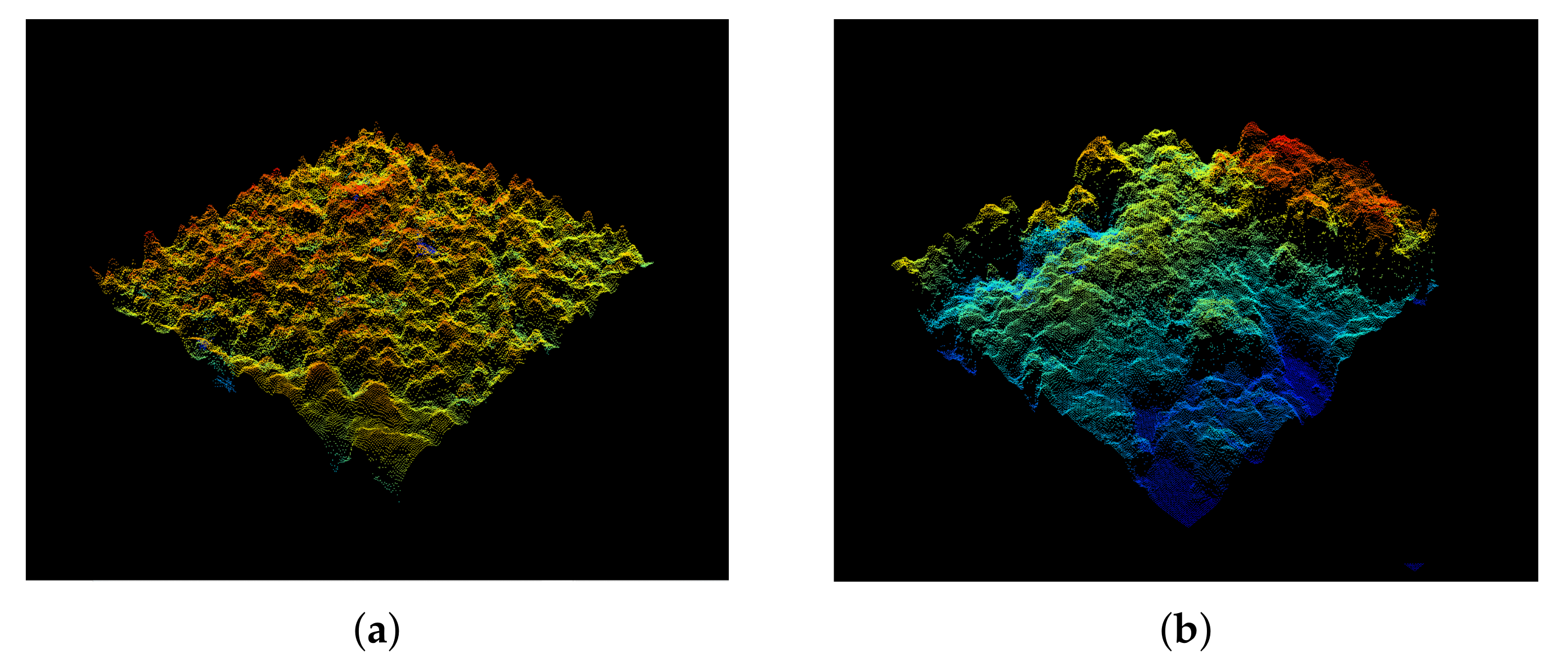
Appendix A. Visual Representation Height Variation Hypothesis

Appendix B. Moving Windows and Spatial Resolution

Appendix C. San Genesio/Jenesien Histograms
Appendix C.1. Berger-Parker

Appendix C.2. CRE

Appendix C.3. Hill

Appendix C.4. Pielou

Appendix C.5. Shannon’s H

Appendix D. Field Data Shannon’s H

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Number | Shannon’s H | Species Richness |
|---|---|---|
| 1 | 0.11 | 5 |
| 2 | 0.98 | 7 |
| 3 | 1.36 | 11 |
| 4 | 0.94 | 9 |
| 5 | 1.05 | 4 |
| 6 | 0.8 | 4 |
| 7 | 0.55 | 4 |
| 8 | 0.96 | 6 |
| 9 | 0.88 | 6 |
| 10 | 0.44 | 7 |
| 11 | 0.65 | 9 |
| 12 | 0.82 | 8 |
| 13 | 0.67 | 7 |
| 14 | 0.32 | 6 |
| 15 | 0.12 | 7 |
| 16 | 0.42 | 4 |
| 17 | 0.44 | 5 |
| 18 | 0.27 | 7 |
| 19 | 0.94 | 6 |
| 20 | 0.7 | 4 |
References
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, D.; Franklin, J.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröbeg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Ball, B.A.; Bradford, M.A.; Coleman, D.C.; Hunter, M.D. Linkages between below and aboveground communities: Decomposer responses to simulated tree species loss are largely additive. Soil Biol. Biochem. 2009, 41, 1155–1163. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Mori, A.S. The potential role of tree diversity in reducing shallow landslide risk. Environ. Manag. 2017, 59, 807–815. [Google Scholar] [CrossRef]
- Castagneyrol, B.; Jactel, H. Unraveling plant–animal diversity relationships: A meta-regression analysis. Ecology 2012, 93, 2115–2124. [Google Scholar] [CrossRef]
- Hanski, I. Habitat loss, the dynamics of biodiversity, and a perspective on conservation. Ambio 2011, 40, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Cayuela, L.; Golicher, D.J.; Benayas, J.M.R.; González-Espinosa, M.; Ramírez, N. Fragmentation, disturbance and tree diversity conservation in tropical montane forests. J. Appl. Ecol. 2006, 43, 1172–1181. [Google Scholar] [CrossRef]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of forest management on species richness: Global meta-analysis and economic trade-offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeely, J.A. The sinking ark: Pollution and the worldwide loss of biodiversity. Biodivers. Conserv. 1992, 1, 2–18. [Google Scholar] [CrossRef]
- Torresani, M.; Rocchini, D.; Sonnenschein, R.; Zebisch, M.; Marcantonio, M.; Ricotta, C.; Tonon, G. Estimating tree species diversity from space in an alpine conifer forest: The Rao’s Q diversity index meets the spectral variation hypothesis. Ecol. Inform. 2019, 52, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Rocchini, D.; Luque, S.; Pettorelli, N.; Bastin, L.; Doktor, D.; Faedi, N.; Feilhauer, H.; Féret, J.B.; Foody, G.M.; Gavish, Y.; et al. Measuring β-diversity by remote sensing: A challeg, for biodiversity monitoring. Methods Ecol. Evol. 2018, 9, 1787–1798. [Google Scholar] [CrossRef] [Green Version]
- Lausch, A.; Bannehr, L.; Beckmann, M.; Boehm, C.; Feilhauer, H.; Hacker, J.; Heurich, M.; Jung, A.; Klenke, R.; Neumann, C.; et al. Linking Earth Observation and taxonomic, structural and functional biodiversity: Local to ecosystem perspectives. Ecol. Indic. 2016, 70, 317–339. [Google Scholar] [CrossRef]
- Rocchini, D.; Balkenhol, N.; Carter, G.A.; Foody, G.M.; Gillespie, T.W.; He, K.S.; Kark, S.; Levin, N.; Lucas, K.; Luoto, M.; et al. Remotely sensed spectral heterogeneity as a proxy of species diversity: Recent advances and open challenges. Ecol. Inform. 2010, 5, 318–329. [Google Scholar] [CrossRef]
- Rocchini, D.; Marcantonio, M.; Da Re, D.; Chirici, G.; Galluzzi, M.; Lenoir, J.; Ricotta, C.; Torresani, M.; Ziv, G. Time-lapsing biodiversity: An open source method for measuring diversity changes by remote sensing. Remote Sens. Environ. 2019, 231, 111192. [Google Scholar] [CrossRef] [Green Version]
- Rocchini, D.; Salvatori, N.; Beierkuhnlein, C.; Chiarucci, A.; de Boissieu, F.; Förster, M.; Garzon-Lopez, C.X.; Gillespie, T.W.; Hauffe, H.C.; He, K.S.; et al. From local spectral species to global spectral communities: A benchmark for ecosystem diversity estimate by remote sensing. Ecol. Inform. 2021, 61, 101195. [Google Scholar] [CrossRef]
- Dalponte, M.; Marzini, S.; Solano-Correa, Y.T.; Tonon, G.; Vescovo, L.; Gianelle, D. Mapping forest windthrows using high spatial resolution multispectral satellite images. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102206. [Google Scholar] [CrossRef]
- Rocchini, D.; Hernández-Stefanoni, J.L.; He, K.S. Advancing species diversity estimate by remotely sensed proxies: A conceptual review. Ecol. Inform. 2015, 25, 22–28. [Google Scholar] [CrossRef]
- Rocchini, D.; Andreo, V.; Förster, M.; Garzon-Lopez, C.X.; Gutierrez, A.P.; Gillespie, T.W.; Hauffe, H.C.; He, K.S.; Kleinschmit, B.; Mairota, P.; et al. Potential of remote sensing to predict species invasions: A modelling perspective. Prog. Phys. Geogr. 2015, 39, 283–309. [Google Scholar] [CrossRef] [Green Version]
- Torresani, M.; Feilhauer, H.; Rocchini, D.; Féret, J.B.; Zebisch, M.; Tonon, G. Which optical traits enable an estimation of tree species diversity based on the Spectral Variation Hypothesis? Appl. Veg. Sci. 2021, 24, e12586. [Google Scholar] [CrossRef]
- Sakowska, K.; MacArthur, A.; Gianelle, D.; Dalponte, M.; Alberti, G.; Gioli, B.; Miglietta, F.; Pitacco, A.; Meggio, F.; Fava, F.; et al. Assessing across-scale optical diversity and productivity relationships in grasslands of the Italian Alps. Remote Sens. 2019, 11, 614. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: A review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Wulder, M.A.; Hall, R.J.; Coops, N.C.; Franklin, S.E. High spatial resolution remotely sensed data for ecosystem characterization. BioScience 2004, 54, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Wüest, R.O.; Bergamini, A.; Bollmann, K.; Baltensweiler, A. LiDAR data as a proxy for light availability improve distribution modelling of woody species. For. Ecol. Manag. 2020, 456, 117644. [Google Scholar] [CrossRef]
- Torresani, M.; Rocchini, D.; Sonnenschein, R.; Zebisch, M.; Hauffe, H.C.; Heym, M.; Pretzsch, H.; Tonon, G. Height variation hypothesis: A new approach for estimating forest species diversity with CHM LiDAR data. Ecol. Indic. 2020, 117, 106520. [Google Scholar] [CrossRef]
- Moudrỳ, V.; Moudrá, L.; Barták, V.; Bejček, V.; Gdulová, K.; Hendrychová, M.; Moravec, D.; Musil, P.; Rocchini, D.; Št’astnỳ, K.; et al. The role of the vegetation structure, primary productivity and senescence derived from airborne LiDAR and hyperspectral data for birds diversity and rarity on a restored site. Landsc. Urban Plan. 2021, 210, 104064. [Google Scholar] [CrossRef]
- Maltamo, M.; Næsset, E.; Vauhkonen, J. Forestry applications of airborne laser scanning. Concepts Case Stud. Manag. Ecosyst. 2014, 27, 2014. [Google Scholar]
- Holopainen, M.; Kankare, V.; Vastaranta, M.; Liang, X.; Lin, Y.; Vaaja, M.; Yu, X.; Hyyppä, J.; Hyyppä, H.; Kaartinen, H.; et al. Tree mapping using airborne, terrestrial and mobile laser scanning–A case study in a heterogeneous urban forest. Urban For. Urban Green. 2013, 12, 546–553. [Google Scholar] [CrossRef]
- Guo, X.; Coops, N.C.; Tompalski, P.; Nielsen, S.E.; Bater, C.W.; Stadt, J.J. Regional mapping of vegetation structure for biodiversity monitoring using airborne lidar data. Ecol. Inform. 2017, 38, 50–61. [Google Scholar] [CrossRef]
- Hakkenbeg, C.R.; Song, C.; Peet, R.K.; White, P.S. Forest structure as a predictor of tree species diversity in the North Carolina Piedmont. J. Veg. Sci. 2016, 27, 1151–1163. [Google Scholar] [CrossRef]
- Bohn, F.J.; Huth, A. The importance of forest structure to biodiversity–productivity relationships. R. Soc. Open Sci. 2017, 4, 160521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverría, C.; Newton, A.C.; Lara, A.; Benayas, J.M.R.; Coomes, D.A. Impacts of forest fragmentation on species composition and forest structure in the temperate landscape of southern Chile. Glob. Ecol. Biogeogr. 2007, 16, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.A.; Stovall, A.E.; Atkins, J.W. Vegetation structural complexity and biodiversity in the Great Smoky Mountains. Ecosphere 2021, 12, e03390. [Google Scholar] [CrossRef]
- Frazer, G.W.; Wulder, M.A.; Niemann, K.O. Simulation and quantification of the fine-scale spatial pattern and heterogeneity of forest canopy structure: A lacunarity-based method designed for analysis of continuous canopy heights. For. Ecol. Manag. 2005, 214, 65–90. [Google Scholar] [CrossRef]
- Ishii, H.T.; Tanabe, S.i.; Hiura, T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperate forest ecosystems. For. Sci. 2004, 50, 342–355. [Google Scholar]
- Alberti, G.; Boscutti, F.; Pirotti, F.; Bertacco, C.; De Simon, G.; Sigura, M.; Cazorzi, F.; Bonfanti, P. A LiDAR-based approach for a multi-purpose characterization of Alpine forests: An Italian case study. Iforest-Biogeosci. For. 2013, 6, 156. [Google Scholar] [CrossRef]
- Huang, Q.; Swatantran, A.; Dubayah, R.; Goetz, S.J. The influence of vegetation height heterogeneity on forest and woodland bird species richness across the United States. PLoS ONE 2014, 9, e103236. [Google Scholar]
- Palmer, M.W.; Earls, P.G.; Hoagland, B.W.; White, P.S.; Wohlgemuth, T. Quantitative tools for perfecting species lists. Environ. Off. J. Int. Environ. Soc. 2002, 13, 121–137. [Google Scholar] [CrossRef]
- Thouverai, E.; Marcantonio, M.; Bacaro, G.; Da Re, D.; Iannacito, M.; Ricotta, C.; Tattoni, C.; Vicario, S.; Rocchini, D. Measuring diversity from space: A global view of the free and open source rasterdiv R package under a coding perspective. Community Ecol. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Rocchini, D.; Thouverai, E.; Marcantonio, M.; Iannacito, M.; Da Re, D.; Torresani, M.; Bacaro, G.; Bazzichetto, M.; Bernardi, A.; Foody, G.M.; et al. rasterdiv—An Information Theory tailored R package for measuring ecosystem heterogeneity from space: To the origin and back. Methods Ecol. Evol. 2021, 12, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhang, J. Tree species diversity of tropical forest vegetation in Xishuangbanna, SW China. Biodivers. Conserv. 1997, 6, 995–1006. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Spellerbeg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Gaudeul, M.; Taberlet, P.; Till-Bottraud, I. Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. Mol. Ecol. 2000, 9, 1625–1637. [Google Scholar] [CrossRef] [Green Version]
- Knoll, C.; Kerschner, H. A glacier inventory for South Tyrol, Italy, based on airborne laser-scanner data. Ann. Glaciol. 2009, 50, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Chen, S.C.; Whitman, D.; Shyu, M.L.; Yan, J.; Zhang, C. A progressive morphological filter for removing nonground measurements from airborne LIDAR data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Woods, M.; Lim, K.; Treitz, P. Predicting forest stand variables from LIDAR data in the Great Lakes St. Lawrence Forest of Ontario. For. Chron. 2008, 84, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Rocchini, D.; Marcantonio, M.; Ricotta, C. Measuring Rao’s Q diversity index from remote sensing: An open source solution. Ecol. Indic. 2017, 72, 234–238. [Google Scholar] [CrossRef]
- Caruso, T.; Pigino, G.; Bernini, F.; Bargagli, R.; Migliorini, M. The Berger–Parker index as an effective tool for monitoring the biodiversity of disturbed soils: A case study on Mediterranean oribatid (Acari: Oribatida) assemblages. In Biodiversity and Conservation in Europe; Springer: Berlin/Heidelbeg, Germany, 2006; pp. 35–43. [Google Scholar]
- Rao, M.; Chen, Y.; Vemuri, B.C.; Wang, F. Cumulative residual entropy: A new measure of information. IEEE Trans. Inf. Theory 2004, 50, 1220–1228. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.H.; Jost, L. Phylogenetic diversity measures based on Hill numbers. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3599–3609. [Google Scholar] [CrossRef]
- Schall, P.; Schulze, E.D.; Fischer, M.; Ayasse, M.; Ammer, C. Relations between forest management, stand structure and productivity across different types of Central European forests. Basic Appl. Ecol. 2018, 32, 39–52. [Google Scholar] [CrossRef]
- Laliberté, E.; Schweiger, A.K.; Legendre, P. Partitioning plant spectral diversity into alpha and beta components. Ecol. Lett. 2020, 23, 370–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Khare, S.; Latifi, H.; Rossi, S. Forest beta-diversity analysis by remote sensing: How scale and sensors affect the Rao’s Q index. Ecol. Indic. 2019, 106, 105520. [Google Scholar] [CrossRef]
- Doxa, A.; Prastacos, P. Using Rao’s quadratic entropy to define environmental heterogeneity priority areas in the European Mediterranean biome. Biol. Conserv. 2020, 241, 108366. [Google Scholar] [CrossRef]
- Ricotta, C. Additive partitioning of Rao’s quadratic diversity: A hierarchical approach. Ecol. Model. 2005, 183, 365–371. [Google Scholar] [CrossRef]
- Ricotta, C.; Szeidl, L. Towards a unifying approach to diversity measures: Bridging the gap between the Shannon entropy and Rao’s quadratic index. Theor. Popul. Biol. 2006, 70, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Ricotta, C.; Pavoine, S.; Bacaro, G.; Acosta, A.T. Functional rarefaction for species abundance data. Methods Ecol. Evol. 2012, 3, 519–525. [Google Scholar] [CrossRef]
- Nagendra, H. Using remote sensing to assess biodiversity. Int. J. Remote Sens. 2001, 22, 2377–2400. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Rocchini, D.; Delucchi, L.; Bacaro, G.; Cavallini, P.; Feilhauer, H.; Foody, G.M.; He, K.S.; Nagendra, H.; Porta, C.; Ricotta, C.; et al. Calculating landscape diversity with information-theory based indices: A GRASS GIS solution. Ecol. Inform. 2013, 17, 82–93. [Google Scholar] [CrossRef]
- Gould, W. Remote sensing of vegetation, plant species richness, and regional biodiversity hotspots. Ecol. Appl. 2000, 10, 1861–1870. [Google Scholar] [CrossRef]
- Rocchini, D.; Chiarucci, A.; Loiselle, S.A. Testing the spectral variation hypothesis by using satellite multispectral images. Acta Oecologica 2004, 26, 117–120. [Google Scholar] [CrossRef]



| TopoSys Falcon II | Optech ALTM 3033 | |
|---|---|---|
| Range | 300–1600 m | 265–3000 m |
| 15 cm < 1200 m | ||
| Elevation accuracy | 5–30 cm depending on satellite constellation | 25 cm < 2000 m |
| 35 cm < 3000 m | ||
| Laser pulse rate | 83 kHz | 33 kHz |
| Scan rate | 653 kHz | Varies with scan angle |
| Laser wavelength | 1560 nm | 1064 nm |
| Index | Symbol | Index Formula | Factors | References |
|---|---|---|---|---|
| Shannon’s H diversity index | H′ | p = relative abundance of a pixel value in a matrix plot (R) | [52] | |
| Berger-Parker’s diversity index | B | p = relative abundance of a pixel value in a matrix plot | [53] | |
| Cumulative Residual Entropy | CRE | N = dimension of the random vector X X = discrete random vector P = the probabilities that the vector of observation is larger or equal to each value of the vector | [54] | |
| Hill’s index of diversity | D | p = relative abundance of a pixel value in a matrix plot (R) q = the ‘order’ of the diversity measure, determines its sensitivity to pixel frequencies | [55] | |
| Pielou’s evenness index | E′ | H = Shannon’s H | ||
| Rao’s Q index of quadratic entropy | Q | p = relative abundance of a pixel value in a matrix plot (R) = distance between the i-th and j-th pixel value = and = 0 i = pixel i j = pixel j | [52] |
| Method ‘Multidimension’ (Rao’s Q Function) | |
|---|---|
| List 1 | chm,pzabove2,pzabovezmean,zentropy,zkurt,zmax,zskew,zsd,zpcum1,zpcum2, zpcum3,zpcum4,zpcum5,zpcum6,zpcum7,zpcum8,zpcum9,zq5,zq10,zq15,zq20, zq25,zq30,zq35,zq40,zq45,zq50,zq55,zq60,zq65,zq70,zq75,zq80,zq85,zq90,zq95 |
| List 2 | pzabove2,pzabovezmean,zentropy,zkurt,zmax,zskew,zsd,zpcum1,zpcum2 zpcum3,zpcum4,zpcum5,zpcum6,zpcum7,zpcum8,zpcum9,zq5,zq10,zq15,zq20, zq25,zq30,zq35,zq40,zq45,zq50,zq55,zq60,zq65,zq70,zq75,zq80,zq85,zq90,zq95 |
| List 3 | chm,pzabove2,pzabovezmean,zentropy,zkurt,zmax,zskew,zsd,zpcum1,zq5 |
| List 4 | chm,pzabove2,pzabovezmean,zentropy,zkurt,zmax,zskew,zsd,zpcum5,zq50 |
| List 5 | chm,pzabove2,pzabovezmean,zmax |
| List 6 | chm,zq95 |
| Shannon’s H Field Data | |
|---|---|
| San Genesio/Jenesien | |
| Number of plots | 20 |
| Mean | 0.67 |
| Standard deviation | 0.33 |
| Min | 0.11 |
| Max | 1.36 |
| Median | 0.68 |
| Spatial Resolution: 2.5 m | ||||
|---|---|---|---|---|
| MW 3 | MW 5 | MW 7 | MW 9 | |
| List 1 | 0.191 | 0.242 | 0.258 | 0.252 |
| List 2 | 0.189 | 0.239 | 0.254 | 0.248 |
| List 3 | 0.214 | 0.248 | 0.248 | 0.232 |
| List 4 | 0.133 | 0.185 | 0.204 | 0.199 |
| List 5 | 0.0762 | 0.0863 | 0.0768 | 0.0623 |
| List 6 | 0.712 | 0.718 | 0.707 | 0.69 |
| Spatial Resolution: 5 m | ||||
| MW 3 | MW 5 | MW 7 | MW 9 | |
| List 1 | 0.279 | 0.22 | 0.166 | 0.141 |
| List 2 | 0.276 | 0.216 | 0.163 | 0.138 |
| List 3 | 0.145 | 0.0998 | 0.0683 | 0.0535 |
| List 4 | 0.0951 | 0.0543 | 0.0273 | 0.0166 |
| List 5 | 0.0623 | 0.027 | 0.0113 | 0.00592 |
| List 6 | 0.577 | 0.494 | 0.433 | 0.386 |
| Spatial Resolution: 10 m | ||||
| MW 3 | MW 5 | MW 7 | MW 9 | |
| List 1 | 0.0002 | 0.01 | 0.01 | 0.01 |
| List 2 | 0.0004 | 0.01 | 0.01 | 0.01 |
| List 3 | 8.19*10 | 0.005 | 0.008 | 0.007 |
| List 4 | 0.04 | 0.07 | 0.08 | 0.08 |
| List 5 | 0.01 | 0.03 | 0.03 | 0.03 |
| List 6 | 0.2 | 0.15 | 0.13 | 0.12 |
| Spatial Resolution: 20 m | ||||
| MW 3 | MW 5 | MW 7 | MW 9 | |
| List 1 | 0.164 | 0.218 | 0.164 | 0.0983 |
| List 2 | 0.168 | 0.222 | 0.168 | 0.101 |
| List 3 | 0.0557 | 0.0906 | 0.121 | 0.109 |
| List 4 | 0.284 | 0.333 | 0.305 | 0.212 |
| List 5 | 0.0784 | 0.125 | 0.155 | 0.132 |
| List 6 | 0.168 | 0.117 | 0.0792 | 0.0471 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamburlin, D.; Torresani, M.; Tomelleri, E.; Tonon, G.; Rocchini, D. Testing the Height Variation Hypothesis with the R rasterdiv Package for Tree Species Diversity Estimation. Remote Sens. 2021, 13, 3569. https://doi.org/10.3390/rs13183569
Tamburlin D, Torresani M, Tomelleri E, Tonon G, Rocchini D. Testing the Height Variation Hypothesis with the R rasterdiv Package for Tree Species Diversity Estimation. Remote Sensing. 2021; 13(18):3569. https://doi.org/10.3390/rs13183569
Chicago/Turabian StyleTamburlin, Daniel, Michele Torresani, Enrico Tomelleri, Giustino Tonon, and Duccio Rocchini. 2021. "Testing the Height Variation Hypothesis with the R rasterdiv Package for Tree Species Diversity Estimation" Remote Sensing 13, no. 18: 3569. https://doi.org/10.3390/rs13183569
APA StyleTamburlin, D., Torresani, M., Tomelleri, E., Tonon, G., & Rocchini, D. (2021). Testing the Height Variation Hypothesis with the R rasterdiv Package for Tree Species Diversity Estimation. Remote Sensing, 13(18), 3569. https://doi.org/10.3390/rs13183569