
Scale and Landscape Features Matter for Understanding Waterbird Habitat Selection



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Satellite Tracking Data
2.3. Environmental Variable
2.4. Univariate Model for Best Scale Selection and Variate Preselection
2.5. Multiscale Model for Habitat Selection
3. Results
3.1. Univariate Model
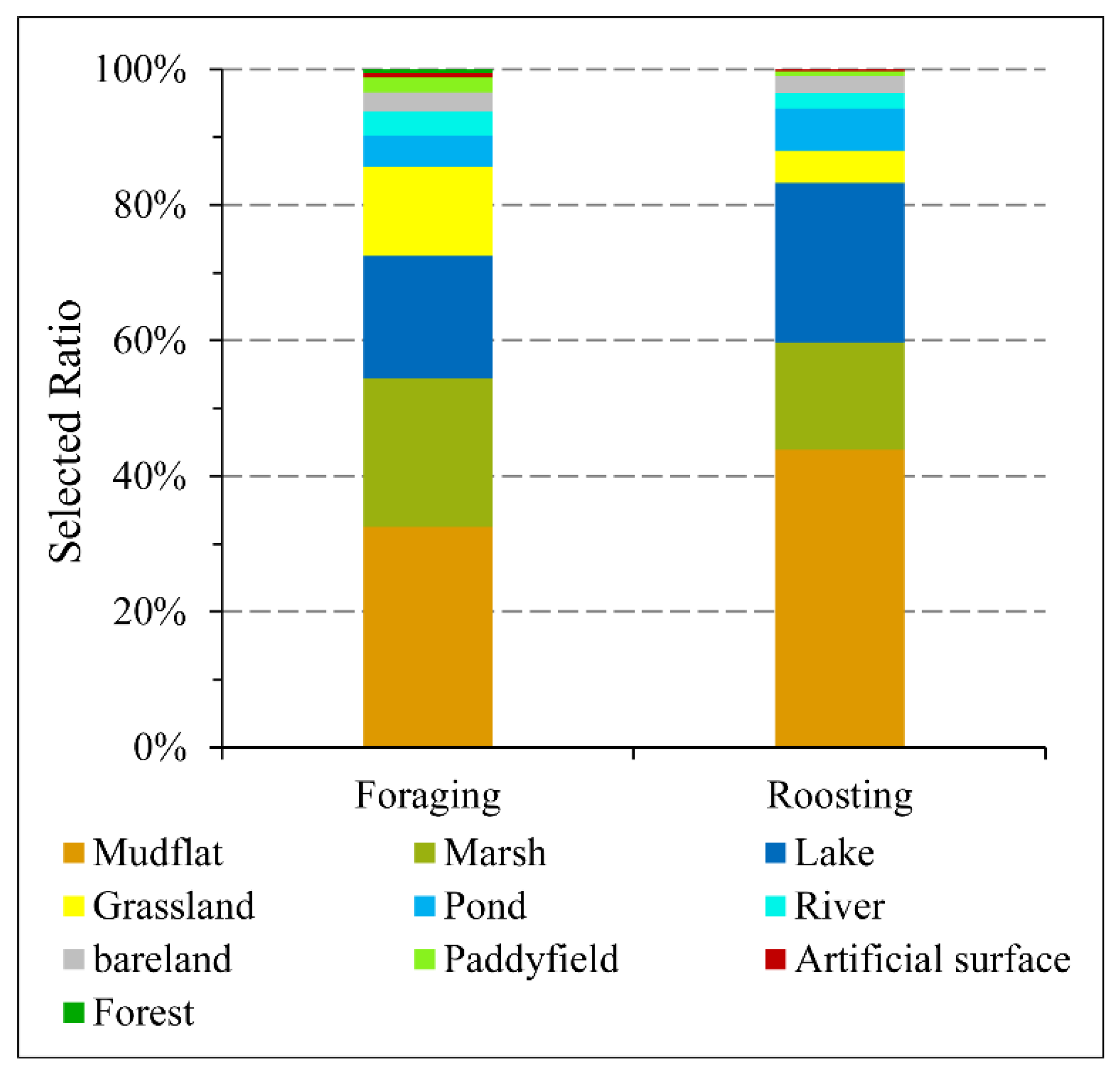
3.2. Multiscale Habitat Selection for Foraging
3.3. Multiscale Habitat Selection for Roosting
4. Discussion
4.1. Landscape Features Are Necessary for Understanding Habitat Selection
4.2. Satellite Tracking Technology Facilitates the Study of the Scale of Effect
4.3. Habitat Selection Research Should Be Detailed According to Habitat Requirements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dzialak, M.R.; Olson, C.V.; Webb, S.L.; Harju, S.M.; Winstead, J.B. Incorporating within- and between-patch resource selection in identification of critical habitat for brood-rearing greater sage-grouse. Ecol. Process. 2015, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, N.; Gao, S.; An, S. Response of wintering waterbird diversity to reclamation history and post-reclamation habitat along Yellow Sea coast. Integr. Zool. 2020, 15, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Frans, V.F.; Augé, A.A.; Edelhoff, H.; Erasmi, S.; Balkenhol, N.; Engler, J.O. Quantifying apart what belongs together: A multi-state species distribution modelling framework for species using distinct habitats. Methods Ecol. Evol. 2018, 9, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Amirkhiz, R.G.; Dixon, M.D.; Palmer, J.S.; Swanson, D.L. Investigating niches and distribution of a rare species in a hierarchical framework: Virginia’s Warbler (Leiothlypis virginiae) at its northeastern range limit. Landsc. Ecol. 2021, 36, 1039–1054. [Google Scholar] [CrossRef]
- Duan, R.; Kong, X.; Huang, M.; Wu, G.; Wang, Z. SDM vs pecies: A software for creating virtual species for species distribution modelling. Ecography 2015, 38, 108–110. [Google Scholar] [CrossRef] [Green Version]
- Engler, J.O.; Stiels, D.; Schidelko, K.; Strubbe, D.; Quillfeldt, P.; Brambilla, M. Avian SDMs: Current state, challenges, and opportunities. J. Avian Biol. 2017, 48, 1483–1504. [Google Scholar] [CrossRef] [Green Version]
- Miguet, P.; Jackson, H.B.; Jackson, N.D.; Martin, A.E.; Fahrig, L. What determines the spatial extent of landscape effects on species? Landsc. Ecol. 2016, 31, 1177–1194. [Google Scholar] [CrossRef]
- Anderson, R.P.; Martínez-Meyer, E. Modeling species’ geographic distributions for preliminary conservation assessments: An implementation with the spiny pocket mice (Heteromys) of Ecuador. Biol. Conserv. 2004, 116, 167–179. [Google Scholar] [CrossRef]
- Graham, C.H.; Elith, J.; Hijmans, R.J.; Guisan, A.; Peterson, A.T.; Loiselle, B.A. The influence of spatial errors in species occurrence data used in distribution models. J. Appl. Ecol. 2007, 45, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Gschweng, M.; Kalko, E.K.V.; Berthold, P.; Fiedler, W.; Fahr, J. Multi-temporal distribution modelling with satellite tracking data: Predicting responses of a long-distance migrant to changing environmental conditions. J. Appl. Ecol. 2012, 49, 803–813. [Google Scholar] [CrossRef]
- Li, X.; Si, Y.; Ji, L.; Gong, P. Dynamic response of East Asian Greater White-fronted Geese to changes of environment during migration: Use of multi-temporal species distribution model. Ecol. Model. 2017, 360, 70–79. [Google Scholar] [CrossRef]
- Blakey, R.V.; Siegel, R.B.; Webb, L.; Dillingham, C.P.; Johnson, M.; Kesler, D.C. Multi-scale habitat selection by Northern Goshawks (Accipiter gentilis) in a fire-prone forest. Biol. Conserv. 2020, 241, 108348. [Google Scholar] [CrossRef]
- Brambilla, M.; Saporetti, F. Modelling distribution of habitats required for different uses by the same species: Implications for conservation at the regional scale. Biol. Conserv. 2014, 174, 39–46. [Google Scholar] [CrossRef]
- Zhang, W.; Li, X.; Yu, L.; Si, Y. Multi-scale habitat selection by two declining East Asian waterfowl species at their core spring stopover area. Ecol. Indic. 2018, 87, 127–135. [Google Scholar] [CrossRef]
- Zuckerberg, B.; Fink, D.; La Sorte, F.A.; Hochachka, W.M.; Kelling, S. Novel seasonal land cover associations for eastern North American forest birds identified through dynamic species distribution modelling. Divers. Distrib. 2016, 22, 717–730. [Google Scholar] [CrossRef] [Green Version]
- Taboada, A.; von Wehrden, H.; Assmann, T.; Srygley, R.B. Integrating life stages into ecological niche models: A case study on tiger beetles. PLoS ONE 2013, 8, e70038. [Google Scholar] [CrossRef] [Green Version]
- Ashrafzadeh, M.R.; Khosravi, R.; Adibi, M.A.; Taktehrani, A.; Wan, H.Y.; Cushman, S.A. A multi-scale, multi-species approach for assessing effectiveness of habitat and connectivity conservation for endangered felids. Biol. Conserv. 2020, 245, 108523. [Google Scholar] [CrossRef]
- Wevers, J.; Beenaerts, N.; Casaer, J.; Zimmermann, F.; Artois, T.; Fattebert, J. Modelling species distribution from camera trap by-catch using a scale-optimized occupancy approach. Remote Sens. Ecol. Conserv. 2021, 7, 534–549. [Google Scholar] [CrossRef]
- Bellamy, C.; Scott, C.; Altringham, J. Multiscale, presence-only habitat suitability models: Fine-resolution maps for eight bat species. J. Appl. Ecol. 2013, 50, 892–901. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, M.C.M.; Cushman, S.A.; Saura, S. Scale dependence in habitat selection: The case of the endangered brown bear (Ursus arctos) in the Cantabrian Range (NW Spain). Int. J. Geogr. Inf. Sci. 2013, 28, 1531–1546. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. Are ecologists conducting research at the optimal scale? Glob. Ecol. Biogeogr. 2015, 24, 52–63. [Google Scholar] [CrossRef]
- McGarigal, K.; Wan, H.Y.; Zeller, K.A.; Timm, B.C.; Cushman, S.A. Multi-scale habitat selection modeling: A review and outlook. Landsc. Ecol. 2016, 31, 1161–1175. [Google Scholar] [CrossRef]
- Martínez-Ruiz, M.; Arroyo-Rodríguez, V.; Franch-Pardo, I.; Renton, K. Patterns and drivers of the scale of effect of landscape structure on diurnal raptors in a fragmented tropical dry forest. Landsc. Ecol. 2020, 35, 1309–1322. [Google Scholar] [CrossRef]
- Wu, J. Effects of changing scale on landscape pattern analysis: Scaling relations. Landsc. Ecol. 2004, 19, 125–138. [Google Scholar] [CrossRef]
- Carrara, E.; Arroyo-Rodríguez, V.; Rivera, J.H.V.; Schondube, J.E.; de Freitas, S.M.; Fahrig, L. Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biol. Conserv. 2015, 184, 117–126. [Google Scholar] [CrossRef]
- Quesnelle, P.E.; Fahrig, L.; Lindsay, K. Effects of habitat loss, habitat configuration and matrix composition on declining wetland species. Biol. Conserv. 2013, 160, 200–208. [Google Scholar] [CrossRef]
- Mukherjee, T.; Sharma, V.; Sharma, L.K.; Thakur, M.; Joshi, B.D.; Sharief, A.; Thapa, A.; Dutta, R.; Dolker, S.; Tripathy, B.; et al. Landscape-level habitat management plan through geometric reserve design for critically endangered Hangul (Cervus hanglu hanglu). Sci. Total Environ. 2021, 777, 146031. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.M.; McGarigal, K. The influence of research scale on bald eagle habitat selection along the lower Hudson River, New York (USA). Landsc. Ecol. 2002, 17, 569–586. [Google Scholar] [CrossRef]
- Baldan, D.; Piniewski, M.; Funk, A.; Gumpinger, C.; Flödl, P.; Höfer, S.; Hauer, C.; Hein, T. A multi-scale, integrative modeling framework for setting conservation priorities at the catchment scale for the Freshwater Pearl Mussel Margaritifera margaritifera. Sci. Total Environ. 2020, 718, 137369. [Google Scholar] [CrossRef] [PubMed]
- IUCN. Ciconia Boyciana. BirdLife International: The IUCN Red List of Threatened Species 2018. 2018. Available online: https://www.iucnredlist.org/ (accessed on 30 August 2021).
- Li, Y.; Zhang, Q.; Cai, Y.; Tan, Z.; Wu, H.; Liu, X.; Yao, J. Hydrodynamic investigation of surface hydrological connectivity and its effects on the water quality of seasonal lakes: Insights from a complex floodplain setting (Poyang Lake, China). Sci. Total Environ. 2019, 660, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Huai, W.; Yang, Z.; Li, D.; Wang, Y. Assessing habitat suitability and habitat fragmentation for endangered Siberian cranes in Poyang Lake region, China. Ecol. Indic. 2021, 125, 107594. [Google Scholar] [CrossRef]
- Li, Y.; Qian, F.; Silbernagel, J.; Larson, H. Community structure, abundance variation and population trends of waterbirds in relation to water level fluctuation in Poyang Lake. J. Great Lakes Res. 2019, 45, 976–985. [Google Scholar] [CrossRef]
- Si, Y.; Xu, Y.; Xu, F.; Li, X.; Zhang, W.; Wielstra, B.; Wei, J.; Liu, G.; Luo, H.; Takekawa, J.; et al. Spring migration patterns, habitat use, and stopover site protection status for two declining waterfowl species wintering in China as revealed by satellite tracking. Ecol. Evol. 2018, 8, 6280–6289. [Google Scholar] [CrossRef]
- Kennerley, R.; Nicoll, M.; Butler, S.; Young, R.; Nuñez-Miño, J.; Brocca, J.; Turvey, S. Home range and habitat data for Hispaniolan mammals challenge assumptions for conservation management. Glob. Ecol. Conserv. 2019, 18, 640. [Google Scholar] [CrossRef]
- Wilson, J.; Sexton, J.O.; Jobe, R.T.; Haddad, N.M. The relative contribution of terrain, land cover, and vegetation structure indices to species distribution models. Biol. Conserv. 2013, 164, 170–176. [Google Scholar] [CrossRef]
- Evans, J.S.; Oakleaf, J.; Cushman, S.A.; Theobald, D. An ArcGIS Toolbox for Surface Gradient and Geomorphometric Modeling, Version 2.0-0; The Nature Conservancy and University of Wyoming: Laramie, WY, USA, 2014; Available online: http://evansmurphy.wix.com/evansspatial (accessed on 30 August 2021).
- Pekel, J.-F.; Cottam, A.; Gorelick, N.; Belward, A.S. High-resolution mapping of global surface water and its long-term changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hill, N.J.; Tobin, A.J.; Reside, A.E.; Pepperell, J.G.; Bridge, T.C.L. Dynamic habitat suitability modelling reveals rapid poleward distribution shift in a mobile apex predator. Glob. Chang. Biol. 2016, 22, 1086–1096. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Clemente, P.; Calvache, M.; Antunes, P.; Santos, R.; Cerdeira, J.O.; Martins, M.J. Combining social media photographs and species distribution models to map cultural ecosystem services: The case of a Natural Park in Portugal. Ecol. Indic. 2019, 96, 59–68. [Google Scholar] [CrossRef]
- Gross, N.; Le Bagousse-Pinguet, Y.; Liancourt, P.; Berdugo, M.; Gotelli, N.J.; Maestre, F.T. Functional trait diversity maximizes ecosystem multifunctionality. Nat. Ecol. Evol. 2017, 1, 0132. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, C.; Guo, Z.; Dai, L.; Wu, Y.; Liu, H.; Li, Y.; Chen, H.; Zhang, Y.; Zhao, Y.; et al. A multiscale approach to identifying spatiotemporal pattern of habitat selection for red-crowned cranes. Sci. Total Environ. 2020, 739, 139980. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ruan, L.; Huang, P.; Luo, H.; Chen, S. Habitat Selection and Protection of Ciconia boyciana during Overwintering Stage in Poyang Lake. J. Anhui Agric. Sci. 2010, 38, 7376–7378. [Google Scholar]
- Xie, P. Habitat Comparison of Oriental White Stork (Ciconia Boyciana) in Different Periods. Master’s Thesis, Northeast Forestry University, Harbin, China, 2018. [Google Scholar]
- Tehrani, N.A.; Naimi, B.; Jaboyedoff, M. Toward community predictions: Multi-scale modelling of mountain breeding birds’ habitat suitability, landscape preferences, and environmental drivers. Ecol. Evol. 2020, 10, 5544–5557. [Google Scholar] [CrossRef]
- Hebblewhite, M.; Haydon, D. Distinguishing technology from biology: A critical review of the use of GPS telemetry data in ecology. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2303–2312. [Google Scholar] [CrossRef]
- Orians, G.H.; Pearson, N.E. On the Theory of Central Place Foraging. In Analysis of Ecological Systems; Ohio State University Press: Columbus, OH, USA, 1979; pp. 155–177. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Variable | Abbreviation(unit) | Description |
|---|---|---|---|
| Landscape Composition | Percentage of focal Land cover type | PLAND (%) | Percentage the landscape comprised of the corresponding patch type. |
| Landscape Configuration | Mean path size | AM(ha) | Mean patch area of focal land cover type. |
| Largest patch index | LPI (%) | Percentage of total landscape area comprised by the largest patch. | |
| Edge density | ED (m/ha) | Edge length on a per unit area basis. | |
| Mean patch shape index | SHAPE_MN (-) | Mean perimeter-area ratio of focal land cover type. | |
| Aggregation index | AI (%) | The number of like adjacencies involving the corresponding class, divided by the maximum possible number of like adjacencies involving the corresponding class. | |
| Interspersion and juxtaposition index | IJI (%) | The observed interspersion over the maximum possible interspersion for the given number of patch types. | |
| Patch cohesion index | COHESION (-) | Physical connectedness of the corresponding patch type. | |
| Shannon’s diversity index | SHDI (-) | Diversity in a landscape. | |
| Patch density | PD (n/100 ha) | The number of patches in the landscape, divided by total landscape area. | |
| Distance to particular land cover types | Eucd_marsh, Eucd_mudflat, Eucd_lake, etc. (m) | Euclidean distance to mudflat, lake, river, marsh, grassland, paddy field, forest or pond. | |
| Natural features | Focal mean of Elevation, Aspect, Slope and Roughness | Elevation (m), Aspect (°), Slope (°) and Roughness (-) | Derived from the DEM using Evans et al. 2014 ArcGIS Geomorphometric & Gradient Metrics Toolbox. |
| Water seasonality | Seasonality (-) | Intra-annual behaviour of water surfaces for a single year (2018). It is the number of months water was present [38]. | |
| Water recurrence | Recurrence (%) | The frequency with which water returns from years to year expressed as a percentage. It is a measurement of the degree of inter-annual variability in the presence of water [38]. | |
| Human disturbance | Road density | Roaddensity (m/m2) | Derived from road layer that is created from Open Street Map. |
| Waterway density | Waterwaydensity (m/m2) | Derived from waterway layer that is created from Open Street Map. | |
| Distance to road, water way and other artificial surfaces | Eucd_road, Eucd_waterway, and Eucd_artificial (m) | Euclidean distance to road, water way and other artificial surfaces. | |
| Landscape composition and configuration of artificial surfaces | PLAND_artificial, AREA_MN_artificial, AI_artificial, etc. | Landscape metrics that describe the characteristic of artificial surface. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhang, Y.; Zhao, L.; Deng, W.; Qian, F.; Ma, K. Scale and Landscape Features Matter for Understanding Waterbird Habitat Selection. Remote Sens. 2021, 13, 4397. https://doi.org/10.3390/rs13214397
Li J, Zhang Y, Zhao L, Deng W, Qian F, Ma K. Scale and Landscape Features Matter for Understanding Waterbird Habitat Selection. Remote Sensing. 2021; 13(21):4397. https://doi.org/10.3390/rs13214397
Chicago/Turabian StyleLi, Jinya, Yang Zhang, Lina Zhao, Wanquan Deng, Fawen Qian, and Keming Ma. 2021. "Scale and Landscape Features Matter for Understanding Waterbird Habitat Selection" Remote Sensing 13, no. 21: 4397. https://doi.org/10.3390/rs13214397
APA StyleLi, J., Zhang, Y., Zhao, L., Deng, W., Qian, F., & Ma, K. (2021). Scale and Landscape Features Matter for Understanding Waterbird Habitat Selection. Remote Sensing, 13(21), 4397. https://doi.org/10.3390/rs13214397