
New Biomass Estimates for Chaparral-Dominated Southern California Landscapes


Abstract
:1. Introduction
2. Materials and Methods
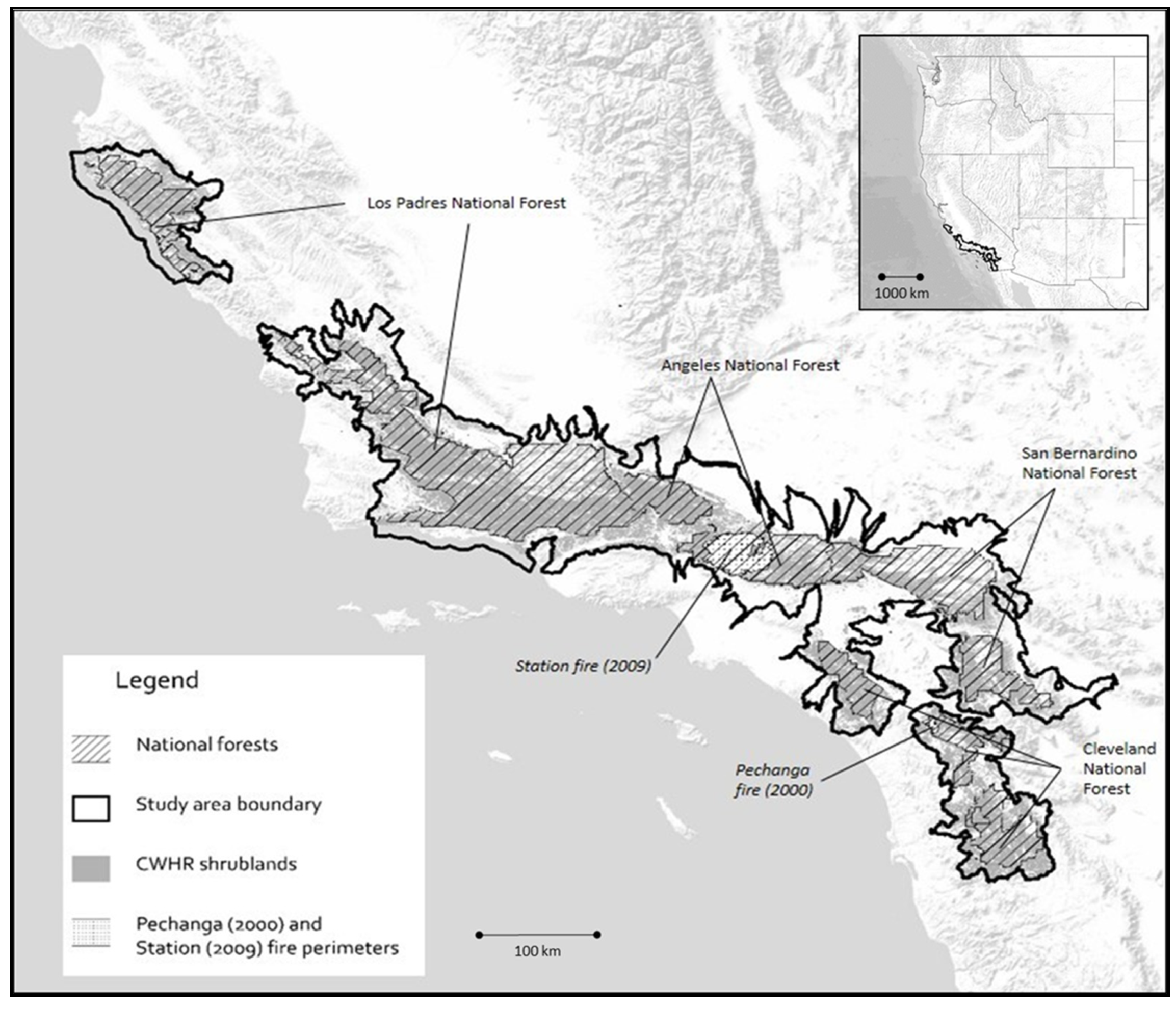
2.1. Study Area
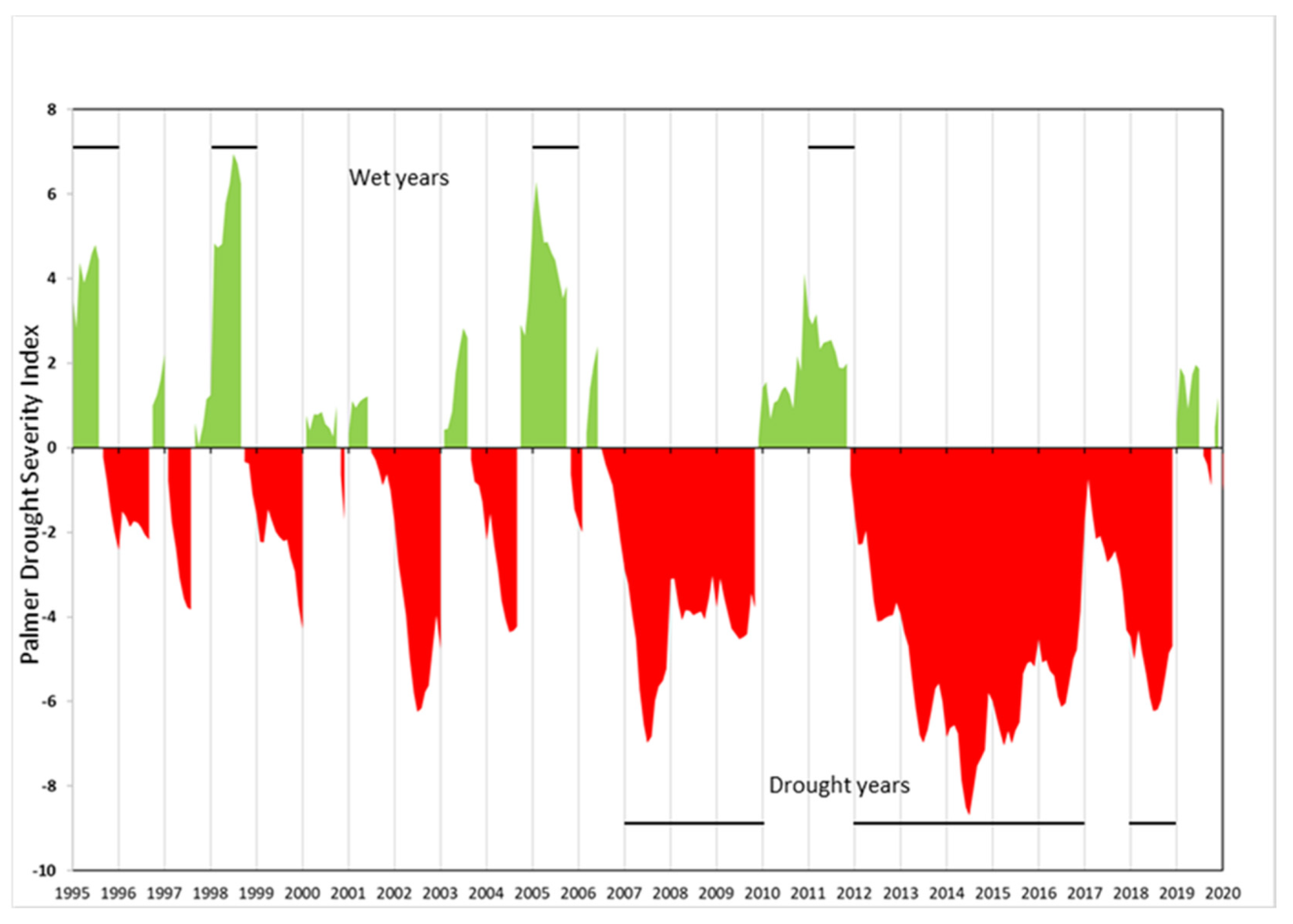
2.2. Environmental Data Layers
2.3. Landsat TM Data and Vegetation Indices
2.4. Plot Data for Biomass Estimates
2.4.1. USFS Forest Inventory and Analysis Data
2.4.2. LANDFIRE Reference Database
2.4.3. Additional Field Plot Data
2.4.4. Assigning Predictor Values to Plots
2.5. Estimating AGLBM with Random Forest
2.6. Comparison of WETAC-UCD Estimates with Other Global and Statewide Biomass Datasets
2.7. Comparison of WETAC-UCD Estimates with Field Estimated Biomass
3. Results
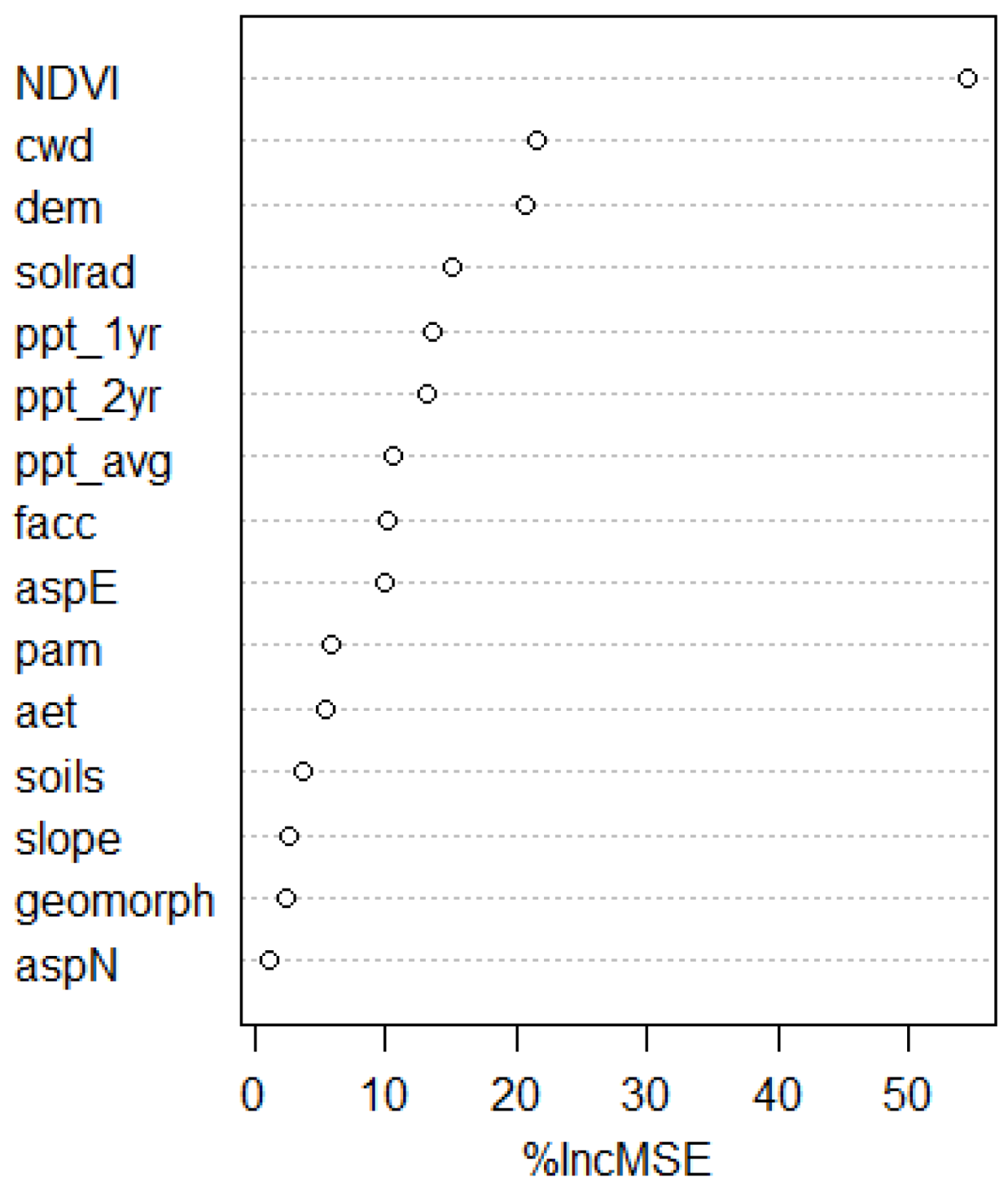
3.1. Estimating AGLBM with Random Forest
3.1.1. Plot Data
3.1.2. RF Model
3.2. Validation by CWHR Vegetation Class
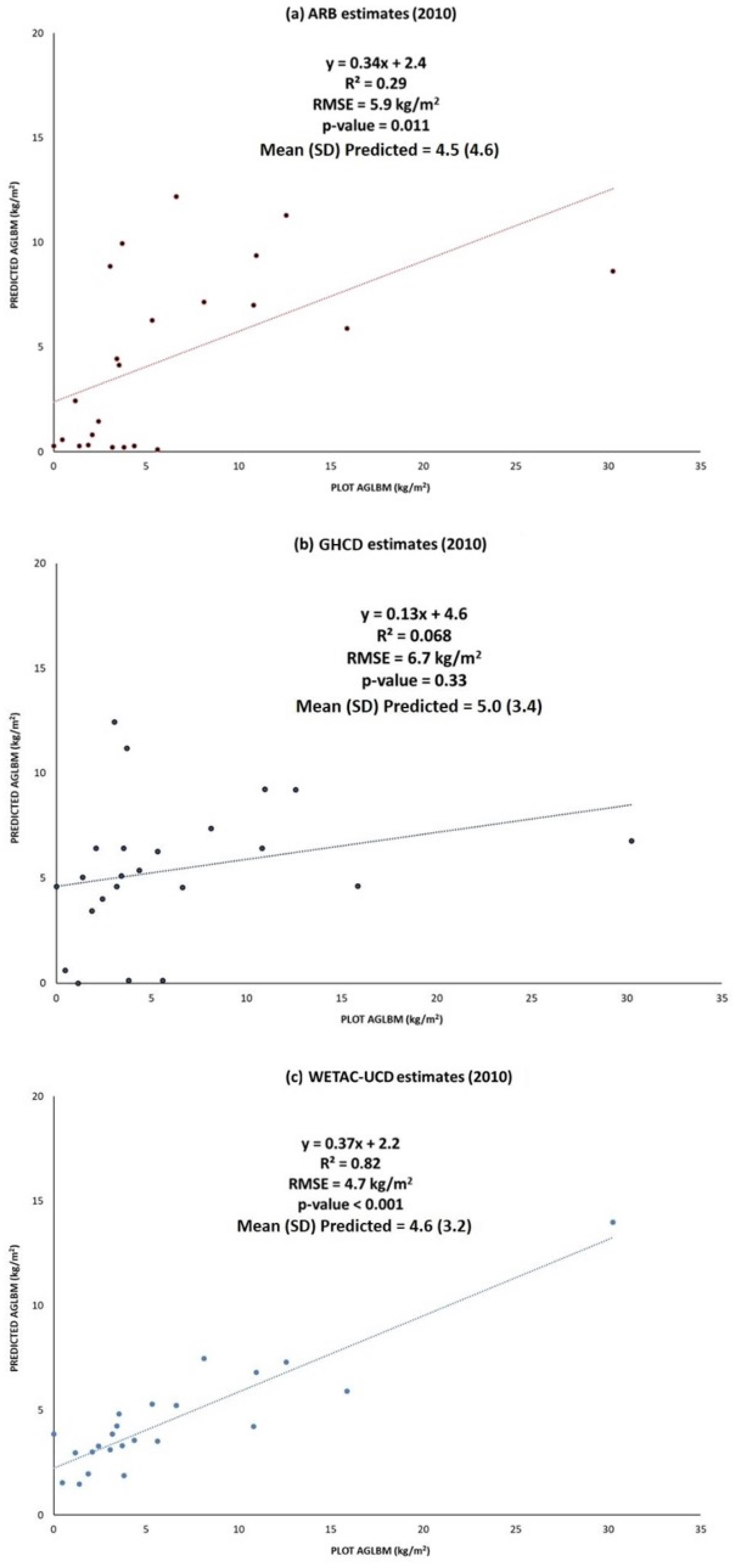
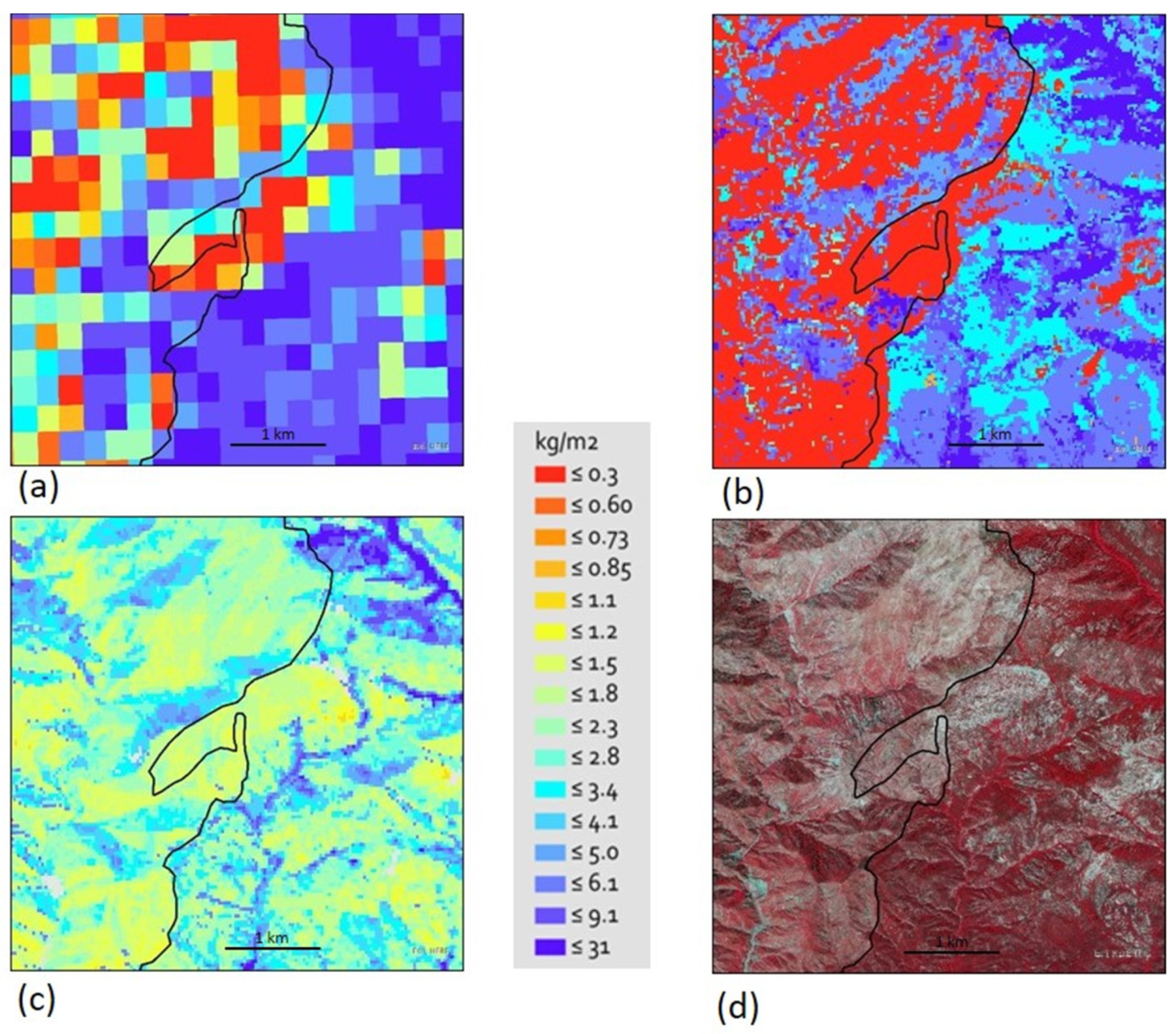
3.3. Comparison of WETAC-UCD Estimates with Other Global and Statewide Biomass Datasets
3.4. Comparison of WETAC-UCD Estimates with Field Measured Biomass
4. Discussion
4.1. Comparison of WETAC-UCD Estimates with Other Global and Statewide Biomass Datasets
4.2. Comparison of WETAC-UCD Estimates with Field Estimates
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shrub Species | R2 | SE | Slope | Intercept | p-Value | N |
|---|---|---|---|---|---|---|
| Adenostoma fasciculatum | 0.63 | 0.02 | 1.2 | 0.053 | <0.001 | 380 |
| Arctostaphylos glandulosa | 0.49 | 0.012 | 0.39 | 0.056 | <0.001 | 223 |
| Eriogonum fasciculatum | 0.66 | 0.0020 | 0.24 | 0.0037 | <0.001 | 24 |
| Quercus berberidifolia | 0.52 | 0.13 | 2.6 | 0.17 | <0.001 | 42 |
| Ribes spp. | 0.95 | 0.0077 | 0.68 | 0.018 | <0.001 | 10 |
References
- Rundel, P. California chaparral and its global significance. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–27. ISBN 978-3-319-68302-7. [Google Scholar]
- Jennings, M.K. Faunal diversity in chaparral ecosystems. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 53–77. ISBN 978-3-319-68302-7. [Google Scholar]
- Underwood, E.C.; Safford, H.D.; Molinari, N.A.; Keeley, J.E. Valuing Chaparral: Ecological, Socio-Economic, and Management Perspectives; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-68303-4. [Google Scholar]
- Jenerette, D.; Park, I.; Andrews, H.; Eberwein, J. Biogeochemical cycling of carbon and Nitrogen in chaparral dominated ecosystems. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 141–169. ISBN 978-3-319-68302-7. [Google Scholar]
- Luo, Y. Terrestrial Carbon–Cycle Feedback to Climate Warming. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 683–712. [Google Scholar] [CrossRef] [Green Version]
- Keeley, J.E.; Davis, F.W. Chaparral. In Terrestrial Vegetation of California, 3rd ed.; University of California Press: Oakland, CA, USA, 2007; pp. 339–366. ISBN 978-0-520-24955-4. [Google Scholar]
- Riggan, P.J.; Goode, S.; Jacks, P.M.; Lockwood, R.N. Interaction of Fire and Community Development in Chaparral of Southern California. Ecol. Monogr. 1988, 58, 155–176. [Google Scholar] [CrossRef]
- Keeley, J.E.; Syphard, A.E. Climate Change and Future Fire Regimes: Examples from California. Geosciences 2016, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Safford, H.D. Man and Fire in Southern California: Doing the Math. Fremontia 2007, 35, 25–29. [Google Scholar]
- Safford, H.D.; Van de Water, K.M. Using Fire Return Interval Departure (FRID) Analysis to Map Spatial and Temporal Changes in Fire Frequency on National Forest Lands in California; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2014. [Google Scholar]
- Jacobsen, A.L.; Pratt, R.B. Extensive Drought-associated Plant Mortality as an Agent of Type-conversion in Chaparral Shrublands. New Phytol. 2018, 7, 498–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmon, M.E.; Marks, B. Effects of Silvicultural Practices on Carbon Stores in Douglas-Fir Western Hemlock Forests in the Pacific Northwest, U.S.A.: Results from a Simulation Model. Can. J. For. Res. 2002, 32, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.G.; Harmon, M.E.; Birdsey, R.A.; Giardina, C.P.; Heath, L.S.; Houghton, R.A.; Jackson, R.B.; McKinley, D.C.; Morrison, J.F.; Murray, B.C.; et al. A Synthesis of the Science on Forests and Carbon for U.S. Forests; Issues in Ecology; Ecological Society of America: Washington, DC, USA, 2010; pp. 1–16. [Google Scholar]
- Keeley, J.E.; Safford, H.D. Fire as an ecosystem process: Chapter 3. In Ecosystems of California; Mooney, H.A., Zavaleta, E.S., Eds.; University of California Press: Oakland, CA, USA, 2016. [Google Scholar]
- Van de Water, K.M.; Safford, H.D. A Summary of Fire Frequency Estimates for California Vegetation before Euro-American Settlement. Fire Ecol. 2011, 7, 26–58. [Google Scholar] [CrossRef]
- Black, C.H. Biomass, nitrogen, and phosphorus accumulation over a southern California fire cycle chronosequence. In Plant Response to Stress. NATO ASI Series; Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; SeriesG: Ecological Sciences; Springer: Berlin/Heidelberg, Germany, 1987; Volume 15. [Google Scholar]
- Bohlman, G.N.; Underwood, E.C.; Safford, H.D. Estimating Biomass in California’s Chaparral and Coastal Sage Scrub Shrublands. Madroño 2018, 65, 28–46. [Google Scholar] [CrossRef]
- Allen, E.B.; Williams, K.; Beyers, J.L.; Phillips, M.; Ma, S.; D’Antonio, C.M. Chaparral Restoration. In Valuing Chaparral: Ecological, Socio-Economic, and Management Perspectives; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 347–384. ISBN 978-3-319-68303-4. [Google Scholar]
- Pratt, R.B.; Jacobsen, A.L.; Ramirez, A.R.; Helms, A.M.; Traugh, C.A.; Tobin, M.F.; Heffner, M.S.; Davis, S.D. Mortality of Resprouting Chaparral Shrubs after a Fire and during a Record Drought: Physiological Mechanisms and Demographic Consequences. Glob. Chang. Biol. 2014, 20, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Syphard, A.D.; Brennan, T.J.; Keeley, J.E. Extent and Drivers of Vegetation Type Conversion in Southern California Chaparral. Ecosphere 2019, 10, e02796. [Google Scholar] [CrossRef]
- Blackard, J.; Finco, M.; Helmer, E.; Holden, G.; Hoppus, M.; Jacobs, D.; Lister, A.; Moisen, G.; Nelson, M.; Riemann, R.; et al. Forest Biomass Using Nationwide Forest Inventory Data and Moderate Resolution Information. Remote Sens. Environ. 2008, 112, 1658–1677. [Google Scholar] [CrossRef]
- Ohmann, J.L.; Gregory, M.J.; Roberts, H.M. Scale Considerations for Integrating Forest Inventory Plot Data and Satellite Image Data for Regional Forest Mapping. Remote Sens. Environ. 2014, 151, 3–15. [Google Scholar] [CrossRef]
- Bolsinger, C.L. Shrubs of California’s Chaparral, Timberland, and Woodland. USDA For. Serv. Pac. Northwest Res. Stn. Portland OR 1989, 50. [Google Scholar] [CrossRef] [Green Version]
- Padgett, P.E.; Allen, E.B. Differential Responses to Nitrogen Fertilization in Native Shrubs and Exotic Annuals Common to Mediterranean Coastal Sage Scrub of California. Plant Ecol. 1999, 144, 93–101. [Google Scholar] [CrossRef]
- Pratt, R.B.; Jacobsen, A.L.; Hernandez, J.; Ewers, F.W.; North, G.B.; Davis, S.D. Allocation Tradeoffs among Chaparral Shrub Seedlings with Different Life History Types (Rhamnaceae). Am. J. Bot. 2012, 99, 1464–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, A.M.; Hogue, T.S. Spatial and Temporal Controls on Post-Fire Hydrologic Recovery in Southern California Watersheds. CATENA 2011, 87, 240–252. [Google Scholar] [CrossRef]
- Keeley, J.E.; Brennan, T.J. Fire-Driven Alien Invasion in a Fire-Adapted Ecosystem. Oecologia 2012, 169, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, P.M.; Lilley, K.A. Sediment delivery, flood control, and physical ecosystem services in southern California chaparral landscapes. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 181–205. ISBN 978-3-319-68302-7. [Google Scholar]
- Galidaki, G.; Zianis, D.; Gitas, I.; Radoglou, K.; Karathanassi, V.; Tsakiri–Strati, M.; Woodhouse, I.; Mallinis, G. Vegetation Biomass Estimation with Remote Sensing: Focus on Forest and Other Wooded Land over the Mediterranean Ecosystem. Int. J. Remote Sens. 2017, 38, 1940–1966. [Google Scholar] [CrossRef] [Green Version]
- Masek, J.G.; Goward, S.N.; Kennedy, R.E.; Cohen, W.B.; Moisen, G.G.; Schleeweis, K.; Huang, C. United States Forest Disturbance Trends Observed Using Landsat Time Series. Ecosystems 2013, 16, 1087–1104. [Google Scholar] [CrossRef] [Green Version]
- Shoshany, M. Satellite Remote Sensing of Natural Mediterranean Vegetation: A Review within an Ecological Context. Prog. Phys. Geogr. 2000, 24, 153–178. [Google Scholar] [CrossRef]
- Filella, I. Reflectance Assessment of Seasonal and Annual Changes in Biomass and CO2 Uptake of a Mediterranean Shrubland Submitted to Experimental Warming and Drought. Remote Sens. Environ. 2004, 90, 308–318. [Google Scholar] [CrossRef]
- Calvão, T.; Palmeirim, J.M. Mapping Mediterranean Scrub with Satellite Imagery: Biomass Estimation and Spectral Behaviour. Int. J. Remote Sens. 2004, 25, 3113–3126. [Google Scholar] [CrossRef]
- Shoshany, M.; Karnibad, L. Mapping Shrubland Biomass along Mediterranean Climatic Gradients: The Synergy of Rainfall-Based and NDVI-Based Models. Int. J. Remote Sens. 2011, 32, 9497–9508. [Google Scholar] [CrossRef]
- Wittenberg, L.; Malkinson, D.; Beeri, O.; Halutzy, A.; Tesler, N. Spatial and Temporal Patterns of Vegetation Recovery Following Sequences of Forest Fires in a Mediterranean Landscape, Mt. Carmel Israel. CATENA 2007, 71, 76–83. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C.J. Impact of past, present and future fire regimes on North American mediterranean shrublands. In Fire, Chaparral, and Survival in Southern California; Sunbelt Publishing: San Diego, CA, USA, 2005; pp. 218–262. ISBN 0-932653-69-3. [Google Scholar]
- Stephenson, N. Actual Evapotranspiration and Deficit: Biologically Meaningful Correlates of Vegetation Distribution across Spatial Scales. J. Biogeogr. 1998, 25, 855–870. [Google Scholar] [CrossRef]
- Mayer, K.E.; Laudenslayer, W.F. A Guide to Wildlife Habitats of California; California Department of Forestry and Fire Protection: Sacramento, CA, USA, 1988. [Google Scholar]
- Kaufman, L.; Rousseeuw, P.J. Clustering by Means of Medoids. In Statistical Data Analysis, Based on the L1 Norm; Dodge, Y., Ed.; Elsevier/North Holland: Amsterdam, The Netherlands, 1987; pp. 405–416. ISBN 978-3-0348-9472-2. [Google Scholar]
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-Scale Hydrologic Modeling for Regional Landscape Applications: The California Basin Characterization Model Development and Performance. Ecol. Process. 2013, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Uyeda, K.A.; Stow, D.A.; O’Leary, J.F.; Tague, C.; Riggan, P.J. Chaparral Growth-Ring Analysis as an Indicator of Stand Biomass Development. Int. J. Wildland Fire 2016, 25, 1086–1092. [Google Scholar] [CrossRef]
- Uyeda, K.A.; Stow, D.A.; Riggan, P.J. Tracking MODIS NDVI Time Series to Estimate Fuel Accumulation. Remote Sens. Lett. 2015, 6, 587–596. [Google Scholar] [CrossRef]
- Keeley, J.E.; Keely, S.C. Chaparral. In North American Terrestrial Vegetation; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: Cambridge, MA, USA, 1988; pp. 165–207. ISBN 0-521-55027-0. [Google Scholar]
- CalFire CalFire Stats and Events. Available online: https://www.fire.ca.gov/stats-events/ (accessed on 1 April 2021).
- Daly, C.; Gibson, W.; Taylor, G.; Johnson, G.; Pasteris, P. A Knowledge-Based Approach to the Statistical Mapping of Climate. Clim. Res. 2002, 22, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally Downscaled and Spatially Customizable Climate Data for Historical and Future Periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Big Remote. Sensed Data Tools Appl. Exp. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Storey, E.A.; Stow, D.A.; O’Leary, J.F. Assessing Postfire Recovery of Chamise Chaparral Using Multi-Temporal Spectral Vegetation Index Trajectories Derived from Landsat Imagery. Remote Sens. Environ. 2016, 183, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Burham, B. Forest Inventory and Analysis Sampling and Plot Design; FIA Fact Sheet Series; USDA Forest Service Forest Inventory and Analysis National Program; USDA Forest Service: Washington DC, USA, 2005. Available online: https://www.fia.fs.fed.us/library/fact-sheets/data-collections/Sampling%20and%20Plot%20Design.pdf (accessed on 4 April 2019).
- McGinnis, T.W.; Shook, C.D.; Keeley, J.E. Estimating Aboveground Biomass for Broadleaf Woody Plants and Young Conifers in Sierra Nevada, California, Forests. West. J. Appl. For. 2010, 25, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, R.H. Responses of Southern California Brushland Vegetation to Fuel Manipulation; University of California: Berkeley, CA, USA, 1978. [Google Scholar]
- Lutes, D.; Keane, R.; Caratti, J.; Key, C.; Benson, N.; Sutherland, S.; Gangi, L. FIREMON: Fire Effects Monitoring and Inventory System; Rocky Mountain Research Station: Fort Collins, CO, USA, 2006. [Google Scholar]
- Jenkins, J.; Chojnacky, D.C.; Heath, L.; Birdsey, R.A. National Scale Biomass Estimators for United States Tree Species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Woodall, C.; Heath, L.; Domke, G.; Nichols, M. Methods and Equations for Estimating Volume, Biomass, and Carbon for Trees in the U.S. Forest Inventory, 2010; Gen. Tech. Rep. NRS-88; Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2010; 30p. [Google Scholar]
- Thompson, M.T.; Toone, M.K. Estimating Root Collar Diameter Growth for Multi-Stem Western Woodland Tree Species on Remeasured Forest Inventory and Analysis Plots. In Moving from Status to Trends: Forest Inventory and Analysis (FIA) Symposium 2012; Morin, R.S., Liknes, G.C., Eds.; Department of Agriculture, Forest Service, Northern Research Station. [CD-ROM]: Newtown Square, PA, USA, 2012; pp. 334–337. Available online: https://www.nrs.fs.fed.us/pubs/gtr/gtr-nrs-p-105papers/53thompson-p-105.pdf (accessed on 3 March 2018).
- Vourlitis, G.L. Aboveground Net Primary Production Response of Semi-Arid Shrublands to Chronic Experimental Dry-Season N Input. Ecosphere 2012, 3, art22. [Google Scholar] [CrossRef]
- Hoaglin, D.C.; John, W. Tukey and Data Analysis. Stat. Sci. 2003, 18, 311–318. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random Forest in Remote Sensing: A Review of Applications and Future Directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Spawn, S.A.; Gibbs, H.K. Global Aboveground and Belowground Biomass Carbon Density Maps for the Year 2010; ORNL Distributed Active Archive Center: Oak Ridge, TN, USA, 2020. [Google Scholar]
- Battles, J.; Gonzalez, P.; Robards, T.; Collins, B.; Saah, D. California Forest and Rangeland Greenhouse Gas Inventory Development FINAL REPORT; California Air Resources Board: Sacramento, CA, USA, 2014; p. 46. [Google Scholar]
- Gonzalez, P.; Battles, J.; Collins, B.; Robards, T.; Saah, D. Aboveground Live Carbon Stock Changes of California Wildland Ecosystems, 2001–2010. For. Ecol. Manag. 2015, 348. [Google Scholar] [CrossRef]
- USDA Forest Service Rapid Assessment of Vegetation Condition after Wildfire (RAVG). Available online: https://fsapps.nwcg.gov/ravg/content/home (accessed on 5 July 2020).
- Dai, A. Dai Global Palmer Drought Severity Index (PDSI). Res. Data Arch. Natl. Cent. Atmos. Res. Comput. Inf. Syst. Lab. 2017. [Google Scholar] [CrossRef]
- Battles, J.J.; Gonzalez, P.; Collins, B.; Robards, T.; Saah, D. California Forest and Rangeland Greenhouse Gas Inventory Development; California Air Resources Board: Sacramento, CA, USA, 2014. [Google Scholar]
- Catchpole, W.R.; Wheeler, C.J. Estimating Plant Biomass: A Review of Techniques. Austral. Ecol. 1992, 17, 121–131. [Google Scholar] [CrossRef]
- Brown, J.K. Bulk Densities of Nonuniform Surface Fuels and Their Application to Fire Modeling. For. Sci. 1981, 27, 667–683. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mahalingam, S.; Weise, D. Chaparral Shrub Bulk Density and Fire Behavior. Available online: https://www.fs.usda.gov/rds/archive/Catalog/RDS-2016-0031 (accessed on 6 June 2020).
- Lutes, D.; (USDA Forest Service, Washington, DC, USA). Personal Communication (email), 29 December 2020.
- Butler, B. Calculating Accurate Aboveground Dry Weight Biomass of Herbaceous Vegetation in the Great Plains: A Comparison of Three Calculations to Determine the Least Resource Intensive and Most Accurate Method. In The Fire Environment—Innovations, Management, and Policy, Proceedings of the RMRS-P-46CD, Destin , FL, USA, 26–30 March 2007; Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ft Collins CO: Fort Collins, CO, USA, 2007. [Google Scholar]
- Pereira, J.; Oliveira, T.; Uva, J. Satellite-Based Estimation of Mediterranean Shrubland Structural Parameters. EARSeL Adv. Remote Sens. 1995, 4, 14–20. [Google Scholar]
- Storey, E.A.; Stow, D.A.; Roberts, D.A. Evaluating Uncertainty in Landsat-Derived Postfire Recovery Metrics Due to Terrain, Soil, and Shrub Type Variations in Southern California. GIScience Remote Sens. 2020, 57, 352–368. [Google Scholar] [CrossRef]
- Chirici, G.; Barbati, A.; Corona, P.; Marchetti, M.; Travaglini, D.; Maselli, F.; Bertini, R. Non-Parametric and Parametric Methods Using Satellite Images for Estimating Growing Stock Volume in Alpine and Mediterranean Forest Ecosystems. Remote Sens. Environ. 2008, 112, 2686–2700. [Google Scholar] [CrossRef] [Green Version]
- Goetz, S.J.; Baccini, A.; Laporte, N.T.; Johns, T.; Walker, W.; Kellndorfer, J.; Houghton, R.A.; Sun, M. Mapping and Monitoring Carbon Stocks with Satellite Observations: A Comparison of Methods. Carbon Balance Manag. 2009, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M. GlobBiomass—Global Datasets of Forest Biomass. PANGAEA 2018. [Google Scholar] [CrossRef]
- Bouvet, A.; Mermoz, S.; Le Toan, T.; Villard, L.; Mathieu, R.; Naidoo, L.; Asner, G. An Above-Ground Biomass Map of African Savannahs and Woodlands at 25 m Resolution Derived from ALOS PALSAR. Remote Sens. Environ. 2018, 206, 156–173. [Google Scholar] [CrossRef]
- Dennison, P.; Roberts, D.; Reggelbrugge, J. Characterizing Chaparral Fuels Using Combined Hyperspectral and Synthetic Aperture Radar Data; University of California, Santa Barbara: Santa Barbara, CA, USA, 2000. [Google Scholar]
- Wu, Z.; Dye, D.; Vogel, J.; Middleton, B. Estimating Forest and Woodland Aboveground Biomass Using Active and Passive Remote Sensing. Photogramm. Eng. Remote Sens. 2016, 82, 271–281. [Google Scholar] [CrossRef]
- Chang, J.; Shoshany, M. Mediterranean Shrublands biomass estimation using Sentinel-1 and Sentinel-2. In Proceedings of the 2016 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Beijing, China, 10–15 July 2016; IEEE: New York, NY, USA, 2016; pp. 5300–5303. [Google Scholar] [CrossRef]
- Narine, L.; Popescu, S.; Malambo, L. Using ICESat-2 to Estimate and Map Forest Aboveground Biomass: A First Example. Remote Sens. 2020, 12, 1824. [Google Scholar] [CrossRef]
- Narine, L.L.; Popescu, S.C.; Malambo, L. Synergy of ICESat-2 and Landsat for Mapping Forest Aboveground Biomass with Deep Learning. Remote Sens. 2019, 11, 1503. [Google Scholar] [CrossRef]
- Duncanson, L.; Neuenschwander, A.; Hancock, S.; Thomas, N.; Fatoyinbo, T.; Simard, M.; Silva, C.A.; Armston, J.; Luthcke, S.B.; Hofton, M.; et al. Biomass Estimation from Simulated GEDI, ICESat-2 and NISAR across Environmental Gradients in Sonoma County, California. Remote Sens. Environ. 2020, 242, 111779. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A New Global Map of Terrestrial Ecoregions Provides an Innovative Tool for Conserving Biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Gago, J.; Gallé, A.; Galmés, J.; Gulías, J.; Medrano, H. Photosynthetic Limitations in Mediterranean Plants: A Review. Response Abiotic Stress. Plants Mediterr. Type Ecosyst. 2014, 103, 12–23. [Google Scholar] [CrossRef]
- Underwood, E.; Hollander, A.D.; Huber, P.R.; Schrader-Patton, C. Mapping the Value of National Forest Landscapes for Ecosystem Service Provision. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 245–270. ISBN 978-3-319-68302-7. [Google Scholar]
- Solek, C.; Resh, V. Water provision in chaparral landscapes: Water quality and water quantity. In Valuing Chaparral; Underwood, E.C., Safford, H.D., Molinari, N.A., Keeley, J.E., Eds.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2018; pp. 207–235. ISBN 978-3-319-68302-7. [Google Scholar]
- Schrader-Patton, C.; Underwood, E.C. SoCal EcoServe. Available online: https://www.fs.fed.us/WETAC/brief/landscapes-socalecoserve.php (accessed on 12 January 2021).
- USDA Forest Service Sustainability and Climate. Available online: https://www.fs.usda.gov/managing-land/sc (accessed on 21 February 2021).




| Predictor Short Name | Predictor Name | Description | Source |
|---|---|---|---|
| aet | Actual evapotranspiration, average 1980–2010 | Units = mm/year | Flint et al. (2017). |
| aspN | Aspect “Northness” | Sin(aspect) | USGS National Elevation Dataset 1/3rd arc-second resolution. |
| aspE | Aspect “Eastness” | Cosin(aspect) | USGS National Elevation Dataset 1/3rd arc-second resolution. |
| cwd | Climatic water deficit, average 1980–2010 | Units = mm/year | http://climate.calcommons.org/variable/climatic-water-deficit, accessed on 17 March 2021. |
| dem | Digital elevation model | Elevation in m above sea level | USGS National Elevation Dataset, 1/3rd arc-second resolution. |
| facc | Flow accumulation | Sum of pixels “uphill” from a pixel | Derived from USGS National Elevation Dataset 1/3rd arc-second resolution. |
| geomorph | Geomorphons | Physiographic landscape facets | https://doi.org/10.1016/j.geomorph.2012.11.005, accessed on 17 March 2021. |
| PAM | Partitioning around medioids | 347 different landscape classes | Kaufman and Rousseuw (1987). |
| ppt_1yr | Annual precipitation | Downscaled 4 km PRISM data, units = mm | ClimateNA tool (Haman (2014)). PRISM—http://www.prism.oregonstate.edu/, accessed on 17 March 2021. |
| ppt_2yr | Biennial precipitation | Downscaled 4 km PRISM data, units = mm | ClimateNA tool (Haman (2014)). PRISM: http://www.prism.oregonstate.edu/, accessed on 17 March 2021. |
| ppt_avg | Precipitation, average, 1980–2000 | Downscaled from PRISM via BCM, units = mm | PRISM: http://climate.calcommons.org/bcm, accessed on 17 March 2021. |
| rch | Groundwater recharge, average 1980–2010 | Units mm/year | Basin Characterization Model (Flint et al. (2017)). |
| run | Water runoff, average 1980–2010 | Units mm/year | Basin Characterization Model (Flint et al. (2017)). |
| soils | Major soil component | String acronym, categorical | SSURGO http://websoilsurvey.nrcs.usda.gov/, accessed on 17 March 2021. |
| solrad | Solar radiation | Watt-hours/m2 year | Annual solar irradiation derived using GRASS 7 r.sun model https://grass.osgeo.org/grass78/manuals/r.sun.html, accessed on 17 March 2021. |
| slope | Topographic slope | Units = degrees | Derived from USGS digital elevation model raster layer. |
| NDVI | Bilinear interpolated NDVI value at plot location | See text for details on data acquisition and processing | USGS Landsat surface reflectance imagery: https://www.usgs.gov/land-resources/nli/landsat/landsat-surface-reflectance, accessed on 17 March 2021. |
| Plot Data Source | Training | Validation | Total |
|---|---|---|---|
| FIA | 401 | 96 | 497 |
| LFRDB | 204 | 50 | 254 |
| Vourlitis et al. | 5 | 5 | 10 |
| Powerhouse fire | 4 | 1 | 5 |
| Total | 614 | 152 | 766 |
| CWHR Type | % Area * | R2 | RMSE | Slope | Intercept | p-Value | N |
|---|---|---|---|---|---|---|---|
| Mixed Chaparral | 53 | 0.23 | 2.4 | 0.25 | 2 | <0.001 | 77 |
| Chamise-redshank Chaparral | 8.2 | 0.23 | 0.96 | 0.22 | 1.3 | 0.35 | 6 |
| Montane Hardwood | 7.8 | 0.24 | 4 | 0.33 | 3.7 | 0.045 | 17 |
| Pinyon-Juniper | 5.9 | 0.51 | 1.5 | 0.65 | 1.2 | <0.001 | 30 |
| All shrub 1 | 7 | 0.3 | 2.3 | 0.29 | 1.9 | <0.001 | 88 |
| All needle-leaf 2 | 12 | 0.53 | 3.9 | 0.47 | 1.9 | <0.001 | 43 |
| All hardwood 3 | 11 | 0.49 | 7 | 0.35 | 3.9 | 0.0002 | 25 |
| All CWHR types | 100 | 0.54 | 3.8 | 0.43 | 2.0 | <0.001 | 170 |
| Aboveground Live Biomass (kg/m2) SHRUB ONLY | Aboveground Live Biomass (kg/m2) NON-SHRUB ONLY | Aboveground Live Biomass (kg/m2) ALL VEGETATION TYPES | |||||||
|---|---|---|---|---|---|---|---|---|---|
| National Forest | WETAC -UCD | ARB | GHCD | WETAC -UCD | ARB | GHCD | WETAC -UCD | ARB | GHCD |
| Los Padres | 3.5 (3.2) | 3.1 (3.5) ** | 6.3 (4.9) ** | 5.8 (5.2) | 5.5 (7.0) ** | 7.5 (5.9) ** | 4.2 (4.1) | 3.9 (5.0) ** | 6.7 (5.2) ** |
| San Bernardino | 3.0 (2.3) | 3.1 (3.2) ** | 4.0 (3.0) ** | 4.7 (2.9) | 4.8 (5.1) ** | 3.9 (3.8) ** | 3.9 (2.8) | 4.0 (4.5) ** | 4.0 (3.4) ** |
| Cleveland | 2.3 (1.6) | 3.1 (2.8) ** | 4.6 (2.8) ** | 4.6 (3.6) | 6.0 (5.4) ** | 5.8 (3.3) ** | 2.5 (2.1) | 3.5 (3.4) ** | 4.8 (2.9) ** |
| Angeles | 2.3 (1.8) | 2.8 (2.9) ** | 5.2 (4.2) ** | 4.8 (3.2) | 5.4 (4.8) ** | 5.8 (4.8) ** | 3.0 (2.6) | 3.6 (3.8) ** | 5.4 (4.4) ** |
| Community Type (CDF-FRAP (FVEG)) | Bohlman AGLBM (kg/m2) | WETAC-UCD AGLBM (kg/m2) | Bohlman Annual Biomass Increment (kg/m2/yr) | WETAC-UCD Biomass Annual Increment (kg/m2/yr) |
|---|---|---|---|---|
| Mixed chaparral | ||||
| Age 1–10 y | 1.0 b | 2.1 | 0.20 b | 0.13 |
| Age 11–20 y | 2.9 b | 3.0 | 0.85 b | 0.29 |
| Chamise chaparral | ||||
| Age 1–10 y | 0.67 | 1.7 | 0.11 b | 0.086 |
| Age 11–30 y | 1.9 | 2.3 | - | 0.18 |
| Coastal Sage Scrub | ||||
| Age 1–10 y | 0.41 b | 1.5 | 0.31 b | 0.075 |
| Age >10 y | 1.0 b | 2.1 | 0.31 b | 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrader-Patton, C.C.; Underwood, E.C. New Biomass Estimates for Chaparral-Dominated Southern California Landscapes. Remote Sens. 2021, 13, 1581. https://doi.org/10.3390/rs13081581
Schrader-Patton CC, Underwood EC. New Biomass Estimates for Chaparral-Dominated Southern California Landscapes. Remote Sensing. 2021; 13(8):1581. https://doi.org/10.3390/rs13081581
Chicago/Turabian StyleSchrader-Patton, Charlie C., and Emma C. Underwood. 2021. "New Biomass Estimates for Chaparral-Dominated Southern California Landscapes" Remote Sensing 13, no. 8: 1581. https://doi.org/10.3390/rs13081581
APA StyleSchrader-Patton, C. C., & Underwood, E. C. (2021). New Biomass Estimates for Chaparral-Dominated Southern California Landscapes. Remote Sensing, 13(8), 1581. https://doi.org/10.3390/rs13081581