
Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia)

 , and
, and
Abstract
:1. Introduction
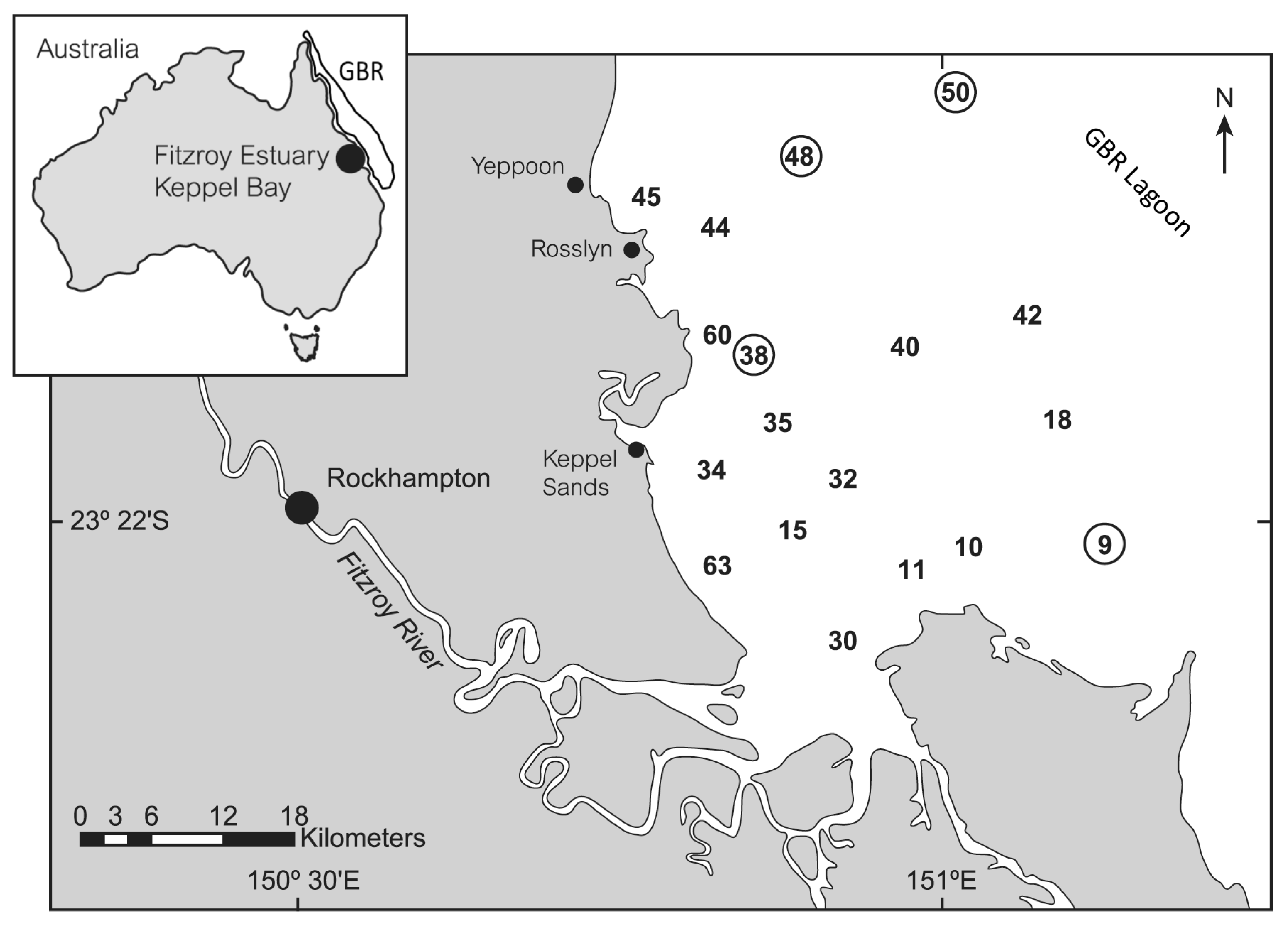
2. Materials and Methods
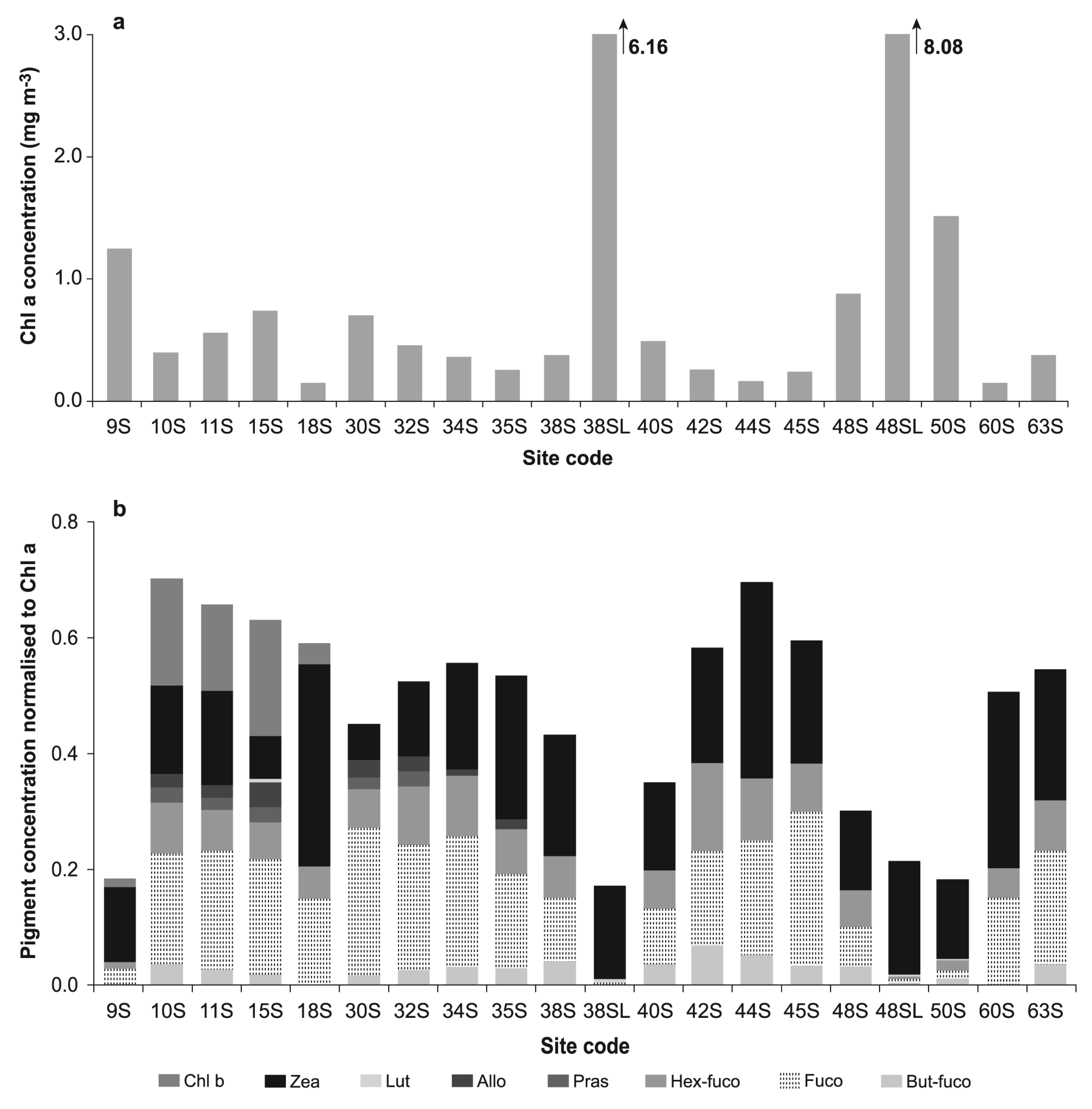
2.1. Pigment Analysis
2.2. Particulate and Detrital Absorption
2.3. CDOM Absorption
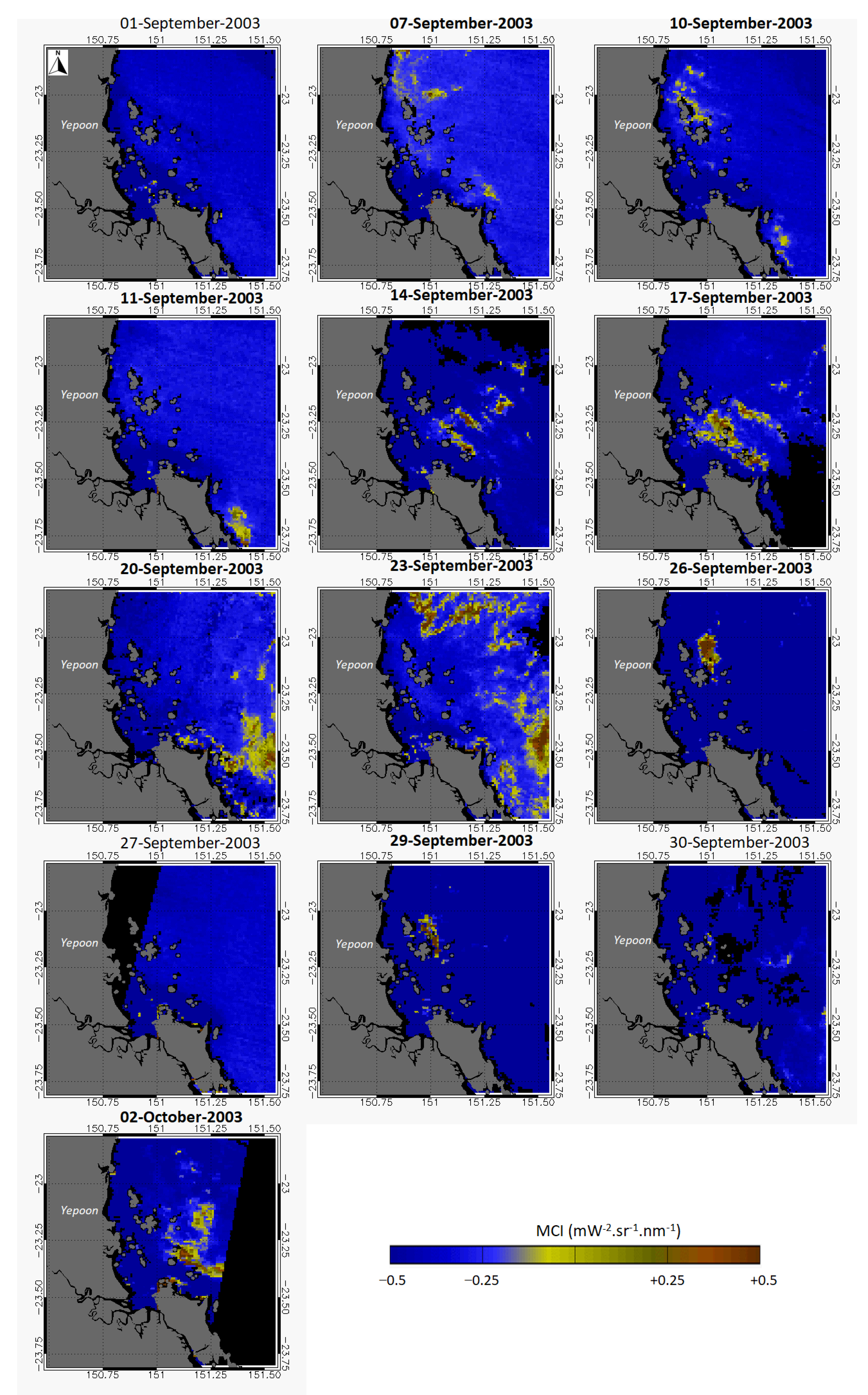
2.4. Remote Sensing
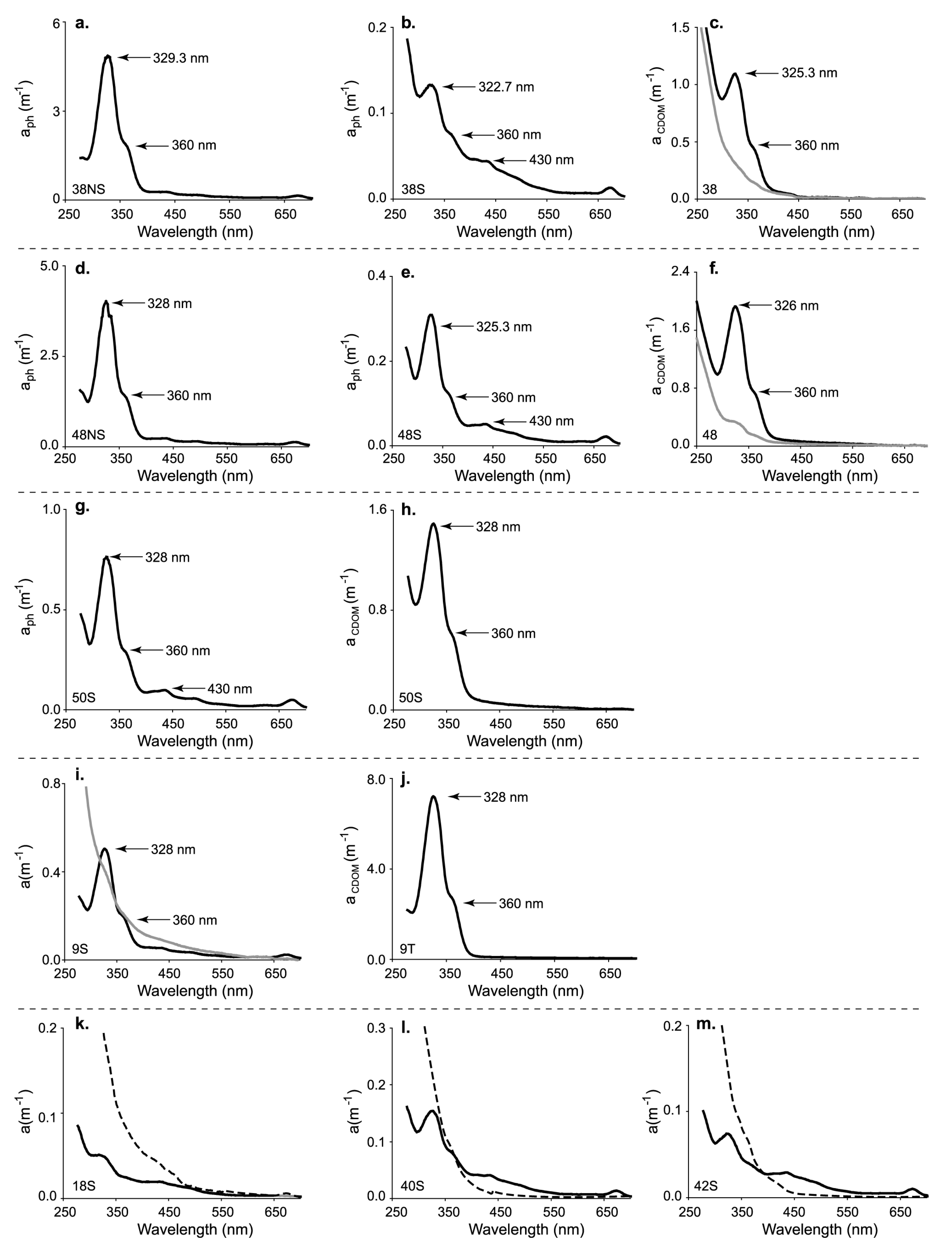
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnes, N.R.E.; Robson, P.W.; Robson, T.M.; Neale, P.J.; Williamson, C.E.; Zepp, R.G.; Wilson, S.R.; Madronich, S.; Andrady, A.L.; Heikkilä, A.M.; et al. Environmental effects of stratospheric ozone depletion, UV radiation, and interactions with climate change: UNEP Environmental Effects Assessment Panel, Update 2020. Photochem. Photobiol. Sci. 2021, 20, 1–67. [Google Scholar] [CrossRef]
- Häder, D.-P.; Williamson, C.E.; Wängberg, S.-Å.; Rautio, M.; Rose, K.C.; Gao, K.; Helbling, E.W.; Sinha, R.P.; Worrest, R. Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem. Photobiol. Sci. 2015, 14, 108–126. [Google Scholar] [CrossRef] [Green Version]
- Vernet, K.; Whitehead, K. Release of untraviolet-absorbing compounds by the red-tide dinoflagellate Lingulodinium polyedra. Mar. Biol. 1996, 127, 35–44. [Google Scholar] [CrossRef]
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.-P. Ultraviolet-absorbing/screening substances in cyanobacteria, phytoplankton and macroalgae. J. Photochem. Photobiol. B Biol. 1998, 47, 83–94. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; MacTavish, H.S.; Dunlap, W.C.; Vesk, M.; Groenewoud, K. Occurrence of UVA- and UVB-absorbing compounds in 152 species (206 strains) of marine microalgae. Mar. Ecol. Prog. Ser. 1999, 189, 35–51. [Google Scholar] [CrossRef]
- Subramaniam, A.; Carpenter, E.J.; Karentz, D.; Falkowski, P.G. Bio-optical properties of the marine diazotrophic cyanobacteria Trichodesmium spp. I. Absorption and photosynthetic action spectra. Limnol. Oceanogr. 1999, 44, 608–617. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Harbour, D.S. A temporal study of mycosporine-like amino acids in surface water phytoplankton from the English Channel and correlation with solar irradiation. J. Mar. Biol. Ass. UK 2003, 83, 1–9. [Google Scholar] [CrossRef]
- Karentz, D.; McEwan, F.S.; Land, M.C.; Dunlap, W.C. Survey of mycosporine-like amino acids compounds in Antarctic marine organisms: Potential protection from ultraviolet exposure. Mar. Biol. 1991, 108, 157–166. [Google Scholar] [CrossRef]
- Banaszak, A.T.; Lesser, M.P.; Kuffner, I.B.; Ondrusek, M. Relationship between ultraviolet (UV) radiation and mycosporine-like amino acids (MAAS) in marine organisms. Bull. Mar. Sci. 1998, 63, 617–628. [Google Scholar]
- Lamare, M.D.; Lesser, M.P.; Barker, M.F.; Barry, T.M.; Schimanski, K.B. Variation in sunscreen compounds (mycosporine-like amino acids) for marine species along a gradient of ultraviolet radiation transmission within doubtful sound, New Zealand. N. Z. J. Mar. Freshw. Res. 2004, 38, 775–793. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, D.K.; Nelson, N.B.; Carlson, C.A.; Prusak, A.C. Production of chromophoric dissolved organic matter (CDOM) in the open ocean by zooplankton and the colonial cyanobacterium Trichodesmium spp. Mar. Ecol. Prog. Ser. 2004, 267, 45–56. [Google Scholar] [CrossRef]
- Yakovleva, I.; Hidaka, M. Diel fluctuations of mycosporine-like amino acids in shallow-water scleractinian corals. Mar. Biol. 2004, 145, 863–873. [Google Scholar] [CrossRef]
- Marchant, H.J.; Davidson, A.T.; Kelly, G.J. UV-B protecting compounds in the marine alga Phaeocystis pouchetii from Antarctica. Mar. Biol. 1991, 109, 391–395. [Google Scholar] [CrossRef]
- Whitenead, K.; Vernet, M. Influence of mycosporine-like aminoacids (MAAs) on UV absorption by particulate and dissolved organic matter in La Jolla Bay. Limnol. Oceanogr. 2000, 45, 1788–1796. [Google Scholar] [CrossRef] [Green Version]
- Moisan, T.A.; Mitchell, B.G. UV absorption by mycosporine-like amino acids in Phaeocystis antarctica Karsten induced by photosynthetically available radiation. Mar. Biol. 2001, 138, 217–227. [Google Scholar] [CrossRef]
- Dupouy, C.; Neveux, J.; Dirberg, G.; Röttgers, R.; Tenório, M.M.B.; Ouillon, S. Bio-optical properties of the marine cyanobacteria Trichodesmium spp. J. App. Rem. Sens. 2008, 2, 023503. [Google Scholar] [CrossRef]
- Tilstone, G.H.; Airs, R.L.; Martinez-Vicente, V.; Widdicombe, C.; Llewellyn, C.A. High concentrations of mycosporine-like amino acids and colored dissolved organic matter in the sea surface microlayer off the Iberian Peninsula. Limnol. Oceanogr. 2010, 55, 1835–1850. [Google Scholar] [CrossRef]
- Stramski, D.; Reynolds, R.A.; Gernez, P.; Röttgers, R.; Wurl, O. Inherent optical properties and particle characteristics of the sea-surface microlayer. Prog. Oceanogr. 2019, 176, 102117. [Google Scholar] [CrossRef]
- Laurion, I.A.; Lami, A.; Sommaruga, R. Distribution of mycosporine-like amino acids and photoprotective carotenoids among freshwater phytoplankton assemblages. Aq. Microb. Ecol. 2002, 26, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef]
- Morrison, J.R.; Nelson, N.B. Seasonal cycle of phytoplankton UV absorption at the Bermuda Atlantic Time-series Study (BATS) site. Limnol. Oceanogr. 2004, 49, 215–224. [Google Scholar] [CrossRef]
- Nelson, N.B.; Carlson, C.A.; Steinberg, D.K. Production of chromophoric dissolved organic matter by Sargasso Sea microbes. Mar. Chem. 2004, 89, 273–287. [Google Scholar] [CrossRef]
- Oubelkheir, K.; Clementson, L.A.; Moore, G.F.; Tilstone, G.H. Production of mycosporine-like amino acids by phytoplankton under ultraviolet radiation exposure in the Sub-Antarctic Zone south of Tasmania. Mar. Ecol. Prog. Ser. 2013, 494, 41–63. [Google Scholar] [CrossRef] [Green Version]
- Dupouy, C.; Frouin, R.; Tedetti, M.; Maillard, M.; Rodier, M.; Lombard, F.; Guidi, L.; Picheral, M.; Neveux, J.; Duhamel, S.; et al. Diazotrophic Trichodesmium impact on UV-Vis radiance and pigment composition in the western tropical South Pacific. Biogeosciences 2018, 15, 5249–5269. [Google Scholar] [CrossRef] [Green Version]
- Dias, A.; Kurin, S.; Thayapurath, S. Influence of environmental parameters on bio-optical characteristics of colored dissolved organic matter in a complex tropical coastal and estuarine region. Est. Coast. Shelf Sci. 2020, 242, 106864. [Google Scholar] [CrossRef]
- Blondeau-Patissier, D.; Brando, V.E.; Lønborg, C.; Leahy, S.M.; Dekker, A.G. Phenology of Trichodesmium spp. blooms in the Great Barrier Reef lagoon, Australia, from the ESA-MERIS 10 year mission. PLoS ONE 2018, 13, e0208010. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, A.; Carpenter, E.J.; Falkowski, P.G. Bio-optical properties of the marine diazotrophic cyanobacteria Trichodesmium spp. II. A reflectance model for remote sensing. Limnol. Oceanogr. 1999, 44, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Westberry, T.K.; Siegel, D.A.; Subramaniam, A. An improved bio-optical model for the remote sensing of Trichodesmium spp. blooms. J. Geophys. Res. 2005, 110, C06012. [Google Scholar] [CrossRef] [Green Version]
- Dupouy, C.; Benielli-Gary, D.; Neveux, J.; Dandonneau, Y.; Westberry, T.K. An algorithm for detecting Trichodesmium surface blooms in the South Western tropical Pacific. Biogeosciences 2011, 8, 3631–3647. [Google Scholar] [CrossRef] [Green Version]
- McKinna, L.I.W.; Furnas, M.J.; Ridd, P.V. A simple binary classification algorithm for the detection of Trichodesmium spp. within the Great barrier Reef using MODIS imagery. Limnol. Oceanogr. Methods 2011, 9, 50–66. [Google Scholar] [CrossRef]
- McKinna, L.I.W. Three decades of ocean-color remote-sensing Trichodesmium spp. in the world’s oceans: A review. Prog. Oceanogr. 2015, 131, 177–199. [Google Scholar] [CrossRef]
- Oubelkheir, K.; Clementson, L.A.; Webster, I.T.; Ford, P.W.; Dekker, A.G.; Radke, L.C.; Daniel, P. Using inherent optical properties to investigate biogeochemical dynamics in a tropical macrotidal coastal system. J. Geophys. Res. 2006, 111, C07021. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C.; Llewellyn, C.A.; Bjornland, D.; Repeta, D.; Welschmeyer, N. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar] [CrossRef]
- Clementson, L.A.; Parslow, J.S.; Turnbull, A.R.; McKenzie, D.C.; Rathbone, C.E. Optical properties of waters in the Australasian sector of the Southern Ocean. J. Geophys. Res. 2001, 106, 31611–31625. [Google Scholar] [CrossRef]
- Mitchell, B.G. Algorithms for determining the absorption coefficient for aquatic particulates using the quantitative filter technique. Proc. Ocean Optics X 1990, 1302, 137–148. [Google Scholar]
- Gower, J.; King, S.; Goncalves, P. Global monitoring of plankton blooms using MERIS MCI. Int. J. Remote Sens. 2008, 29, 6209–6216. [Google Scholar] [CrossRef]
- Gower, J.; King, S. On the importance of a band at 709 nm. In Proceedings of the 2nd MERIS-AATSR Workshop, ESRIN, Frascati, Italy, 23 September 2008. [Google Scholar]
- Jeffrey, S.W.; Vesk, M. Introduction to marine phytoplankton and their pigment signatures. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 37–84. [Google Scholar]
- Jeffrey, S.W.; Wright, S.W. Photosynthetic pigments in marine microalgae: Insights from cultures and the sea. In Algal Cultures, Analogues of Blooms and Applications; Subba Rao, D.V., Ed.; Science Publishers: Enfield, NH, USA, 2006; pp. 33–90. [Google Scholar]
- Kuznetsova, M.; Lee, C.; Aller, J.; Frew, N. Enrichment of amino acids in the sea surface microlayer at coastal and open ocean sites in the North Atlantic Ocean. Limnol. Oceanogr. 2004, 49, 1605–1619. [Google Scholar] [CrossRef]
- Whitehead, K.; Hedges, J.I. Photodegradation and photosensitization of mycosporine-like amino acids. J. Photochem. Photobiol. B Biol. 2005, 80, 115–121. [Google Scholar] [CrossRef]
- Böhm, G.A.; Pfleiderer, W.; Böger, P.; Scherer, S. Structure of a novel oligosaccharide-mycosporine-amino acid ultraviolet A/B sunscreen pigment from the terrestrial cyanobacterium Nostoc commune. J. Biol. Chem. 1995, 270, 8536–8539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furnas, M.J. Pelagic Trichodesmium (=Oscillatoria) in the Great Barrier Reef region. In Marine Pelagic Cyanobacteria: Trichodesmium and Other Diazotrophs; Carpenter, E.J., Capone, D.G., Rueter, J.G., Eds.; Kluwer Academic Publishers: Bamberg, Germany, 1992; pp. 265–272. [Google Scholar]
- Jones, G.B. Effect of Trichodesmium blooms on water quality in the Great Barrier Reef Lagoon. In Marine Pelagic Cyanobacteria: Trichodesmium and Other Diazotrophs; Carpenter, E.J., Capone, D.G., Rueter, J.G., Eds.; Kluwer Academic Publishers: Bamberg, Germany, 1992; pp. 273–287. [Google Scholar]
- Bell, P.R.F.; Elemetri, I.; Uwins, P. Nitrogen fixation by Trichodesmium spp. in the Central and Northern Great Barrier Reef Lagoon: Relative importance of the fixed-nitrogen load. Mar. Ecol. Prog. Ser. 1999, 186, 119–126. [Google Scholar] [CrossRef]
- Walsby, A.E. The gas vesicles and buoyancy of Trichodesmium. In Marine Pelagic Cyanobacteria: Trichodesmium and Other Diazotrophs; Carpenter, E.J., Capone, D.G., Rueter, J.G., Eds.; Kluwer Academic Publishers: Bamberg, Germany, 1992; pp. 141–162. [Google Scholar]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Rtichodesmium, a globally significant marine cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Obernosterer, I.; Catala, P.; Lami, R.; Caparros, J.; Ras, J.; Bricaud, A.; Dupuy, C.; van Wambeke, F.; Lebaron, P. Biochemical characteristics and bacterial community structure of the sea surface microlayer in the South Pacific Ocean. Biogeosciences 2008, 5, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Webster, I.T.; Hutchinson, P.A. Effect of wind on the distribution of phytoplankton cells in lakes revisited. Limnol. Oceanogr. 1994, 39, 365–373. [Google Scholar] [CrossRef]
- Riegger, L.; Robinson, D. Photoinduction of UV-absorbing compounds in Antarctic diatoms and Phaeocystis antarctica. Mar. Ecol. Prog. Ser. 1997, 160, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Roesler, C.S.; Perry, M.J.; Carder, K.L. Modeling in situ phytoplankton absorption from total absorption spectra in productive inland marine waters. Limnol. Oceanogr. 1989, 34, 1510–1523. [Google Scholar] [CrossRef] [Green Version]
- Bricaud, A.; Morel, A.; Prieur, L. Absorption by dissolved organic matter of the sea (yellow substance) in the UV and visible domains. Limnol. Oceanogr. 1981, 26, 43–53. [Google Scholar] [CrossRef]
- Babin, M.; Stramski, D.; Ferrari, G.M.; Claustre, H.; Bricaud, A.; Obolensky, G.; Hoepffner, N. Variations in the light absorption coefficients of phytoplankton nonalgal particles and dissolved organic matter in coastal waters around Europe. J. Geophys. Res. 2003, 108, 3211. [Google Scholar] [CrossRef]
- Twardowski, M.S.; Donaghay, P.L. Separating in situ and terrigenous sources of absorption by dissolved materials in coastal waters. J. Geophys. Res. 2001, 106, 2545–2560. [Google Scholar] [CrossRef]
- Del Vecchio, R.; Blough, N.V. Photobleaching of chromophoric dissolved organic matter in natural waters: Kinetics and modelling. Mar. Chem. 2002, 78, 231–253. [Google Scholar] [CrossRef]
- Nelson, N.B.; Siegel, D.A.; Carlson, C.A.; Swan, C.M. Tracing global biogeochemical cycles and meridional overturning circulation using chromophoric dissolved organic matter. Geophys. Res. Lett. 2010, 37, L03610. [Google Scholar] [CrossRef]
- Lønborg, C.; McKinna, L.I.W.; Slivkoff, M.M.; Carreira, C. Coloured dissolved organic matter dynamics in the Great Barrier Reef. Cont. Shelf Res. 2021, 219, 104395. [Google Scholar] [CrossRef]
- Borstad, G.A.; Gower, J.F.R.; Carpenter, E.J. Development of algorithms for remote sensing of Trichodesmium blooms. In Marine Pelagic Cyanobacteria: Trichodesmium and Other Diazotrophs; Carpenter, E.J., Capone, D.G., Rueter, J.G., Eds.; Kluwer Academic Publishers: Bamberg, Germany, 1992; pp. 193–210. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude (°S) | Longitude (°E) | Date (Local) | Time (Local) |
|---|---|---|---|---|
| 9 | 23 41.84 | 151 15.63 | 6 September 2003 | 1035 |
| 10 | 23 40.27 | 151 04.60 | 6 September 2003 | 1252 |
| 11 | 23 42.70 | 150 99.06 | 6 September 2003 | 1424 |
| 15 | 23 38.55 | 150 89.78 | 7 September 2003 | 0959 |
| 18 | 23 31.98 | 151 11.69 | 7 September 2003 | 1231 |
| 30 | 23 50.16 | 150 95.95 | 9 September 2003 | 0847 |
| 32 | 23 36.52 | 150 95.20 | 9 September 2003 | 1040 |
| 34 | 23 34.50 | 150 83.44 | 9 September 2003 | 1226 |
| 35 | 23 32.16 | 150 88.90 | 9 September 2003 | 1328 |
| 38 | 23 26.24 | 150 86.80 | 10 September 2003 | 0840 |
| 40 | 23 26.27 | 150 99.51 | 10 September 2003 | 1018 |
| 42 | 23 24.29 | 151 09.43 | 10 September 2003 | 1143 |
| 44 | 23 17.09 | 150 83.85 | 10 September 2003 | 1500 |
| 45 | 23 15.21 | 150 79.52 | 11 September 2003 | 0747 |
| 48 | 23 10.49 | 150 91.08 | 11 September 2003 | 0941 |
| 50 | 23 07.11 | 151 04.13 | 11 September 2003 | 1130 |
| 60 | 23 25.64 | 150 84.09 | 12 September 2003 | 0859 |
| 63 | 23 42.17 | 150 83.24 | 12 September 2003 | 1050 |
| 328 nm | 440 nm | 676 nm | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Site | Slope | aCDOM | ap | aph | aCDOM | ap | aph | aCDOM | ap | aph |
| 9S | 0.012 | 0.4050 | 0.4979 | 0.4372 | 0.0840 | 0.0480 | 0.0311 | 0.0002 | 0.0185 | 0.0167 |
| 9T | 0.044 | 7.1430 | 0.0446 | 0.0002 | ||||||
| 10S | 0.015 | 0.2635 | 0.2971 | 0.0246 | 0.0455 | 0.0946 | 0.0283 | 0.0000 | 0.0155 | 0.0103 |
| 11S | 0.014 | 0.3335 | 0.4430 | 0.0592 | 0.0658 | 0.1366 | 0.0368 | 0.0000 | 0.0193 | 0.0114 |
| 15S | 0.015 | 0.5632 | 0.3941 | 0.0411 | 0.0965 | 0.1210 | 0.0374 | 0.0007 | 0.0206 | 0.0145 |
| 18S | 0.013 | 0.1982 | 0.0482 | 0.0292 | 0.0366 | 0.0179 | 0.0129 | 0.0017 | 0.0047 | 0.0042 |
| 30S | 0.015 | 0.4409 | 1.9937 | 0.0037 | 0.0795 | 0.4467 | 0.0434 | 0.0012 | 0.0401 | 0.0139 |
| 32S | 0.025 | 0.2964 | 0.3273 | 0.0396 | 0.0126 | 0.1089 | 0.0365 | 0.0000 | 0.0184 | 0.0128 |
| 34S | 0.022 | 0.5070 | 0.3349 | 0.0374 | 0.0474 | 0.0953 | 0.0234 | 0.0000 | 0.0132 | 0.0078 |
| 35S | 0.017 | 0.4862 | 0.1957 | 0.0206 | 0.0767 | 0.0612 | 0.0175 | 0.0011 | 0.0098 | 0.0061 |
| 38S | 0.024 | 0.2966 | 0.1312 | 0.0715 | 0.0218 | 0.0414 | 0.0254 | 0.0007 | 0.0117 | 0.0104 |
| 38NS | 0.035 | 1.0771 | 4.7801 | 4.4537 | 0.0316 | 0.2286 | 0.1784 | 0.0000 | 0.1062 | 0.1000 |
| 40S | 0.028 | 0.2217 | 0.1514 | 0.1028 | 0.0071 | 0.0379 | 0.0260 | 0.0000 | 0.0115 | 0.0105 |
| 42S | 0.031 | 0.1491 | 0.0723 | 0.0469 | 0.0062 | 0.0275 | 0.0204 | 0.0000 | 0.0086 | 0.0078 |
| 44S | 0.020 | 0.3395 | 0.1186 | 0.0136 | 0.0136 | 0.0377 | 0.0426 | 0.0151 | 0.0000 | 0.0065 |
| 45S | 0.014 | 0.3693 | 0.1601 | 0.0179 | 0.0637 | 0.0537 | 0.0185 | 0.0002 | 0.0088 | 0.0061 |
| 48S | 0.025 | 0.3166 | 0.3056 | 0.2654 | 0.0254 | 0.0477 | 0.0361 | 0.0000 | 0.0187 | 0.0169 |
| 48NS | 0.038 | 1.9050 | 3.9838 | 3.6738 | 0.0612 | 0.1966 | 0.0995 | 0.0005 | 0.0953 | 0.0674 |
| 50S | 0.037 | 1.4810 | 0.7567 | 0.7114 | 0.0476 | 0.0905 | 0.0773 | 0.0000 | 0.0431 | 0.0410 |
| 60S | 0.016 | 0.3246 | 0.1417 | 0.0077 | 0.0461 | 0.0427 | 0.0091 | 0.0000 | 0.0057 | 0.0030 |
| 63S | 0.017 | 0.5533 | 0.3258 | 0.0101 | 0.0766 | 0.0947 | 0.0207 | 0.0000 | 0.0122 | 0.0071 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clementson, L.A.; Oubelkheir, K.; Ford, P.W.; Blondeau-Patissier, D. Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia). Remote Sens. 2022, 14, 3686. https://doi.org/10.3390/rs14153686
Clementson LA, Oubelkheir K, Ford PW, Blondeau-Patissier D. Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia). Remote Sensing. 2022; 14(15):3686. https://doi.org/10.3390/rs14153686
Chicago/Turabian StyleClementson, Lesley A., Kadija Oubelkheir, Phillip W. Ford, and David Blondeau-Patissier. 2022. "Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia)" Remote Sensing 14, no. 15: 3686. https://doi.org/10.3390/rs14153686
APA StyleClementson, L. A., Oubelkheir, K., Ford, P. W., & Blondeau-Patissier, D. (2022). Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia). Remote Sensing, 14(15), 3686. https://doi.org/10.3390/rs14153686