
Improving Remote Estimation of Vegetation Phenology Using GCOM-C/SGLI Land Surface Reflectance Data


Abstract
:1. Introduction
- (1)
- Are the observed seasonal trajectories of vegetation between the SGLI and near-surface consistent?
- (2)
- What is the agreement between the phenological transition dates derived from the SGLI and near-surface?
- (3)
- What is the difference in the spatial pattern of the phenological transition dates between the SGLI and the VIIRS?
2. Materials and Methods
2.1. SGLI Land Surface Reflectance and Vegetation Indices
2.2. Detection of Phenological Transition Dates from Time Series
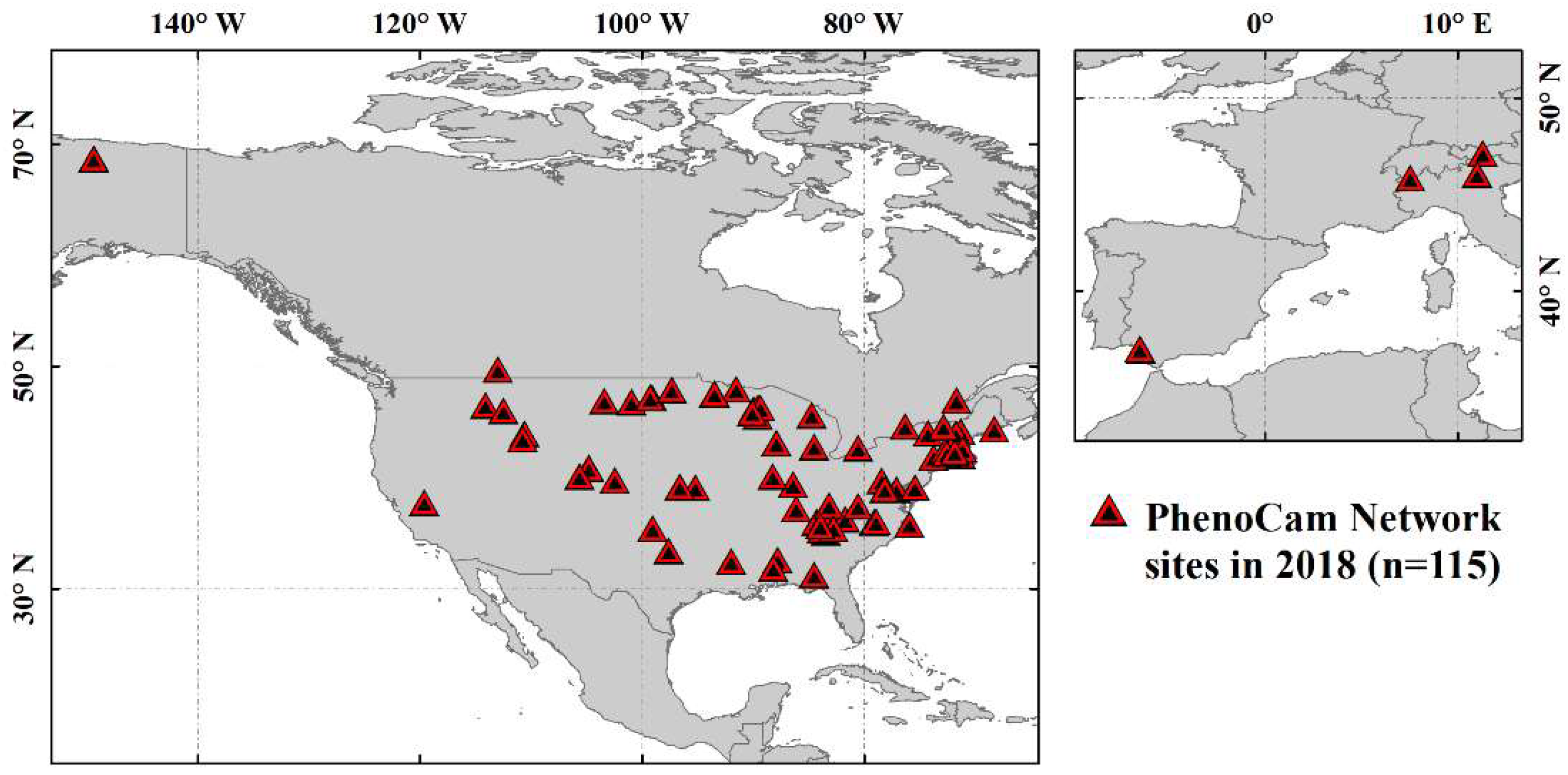
2.3. Near-Surface Phenology Observation Data
2.4. MODIS and VIIRS Land Surface Phenology Products
2.5. Evaluation of SGLI Phenology Using Near-Surface Phenology Observation
2.6. Comparison of SGLI Phenology with VIIRS Phenology
3. Results
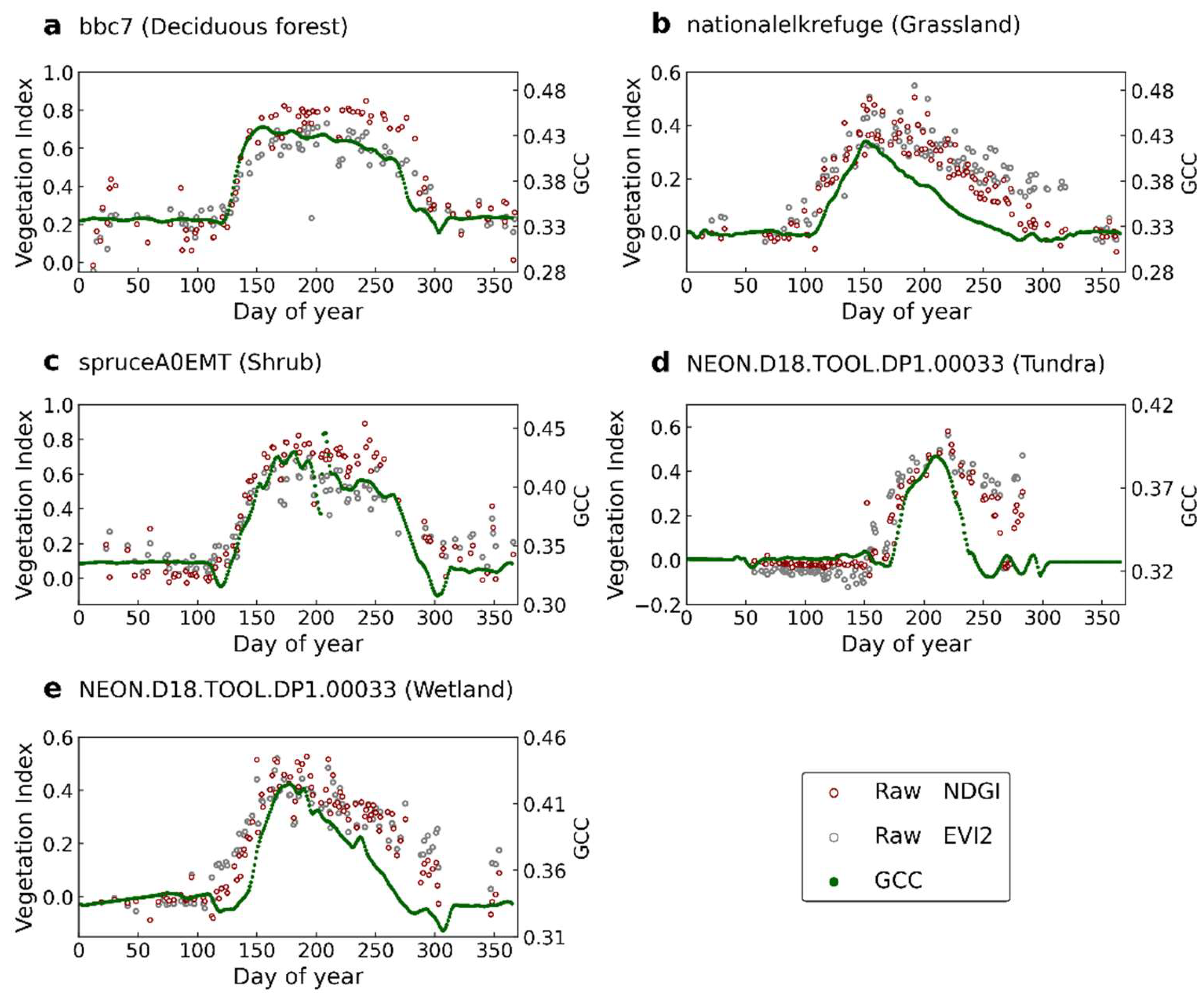
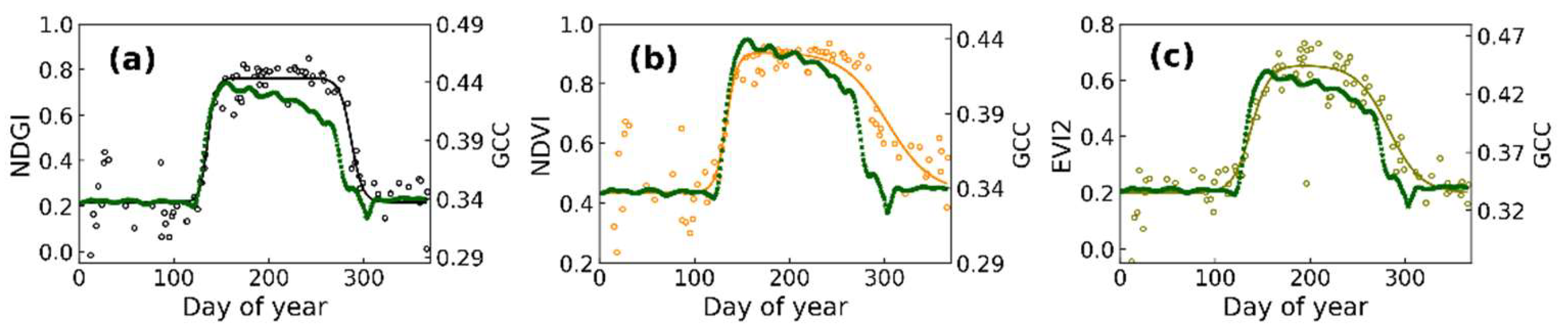
3.1. Time Series of SGLI and Near-Surface Observations and Determined Phenological Transition Dates
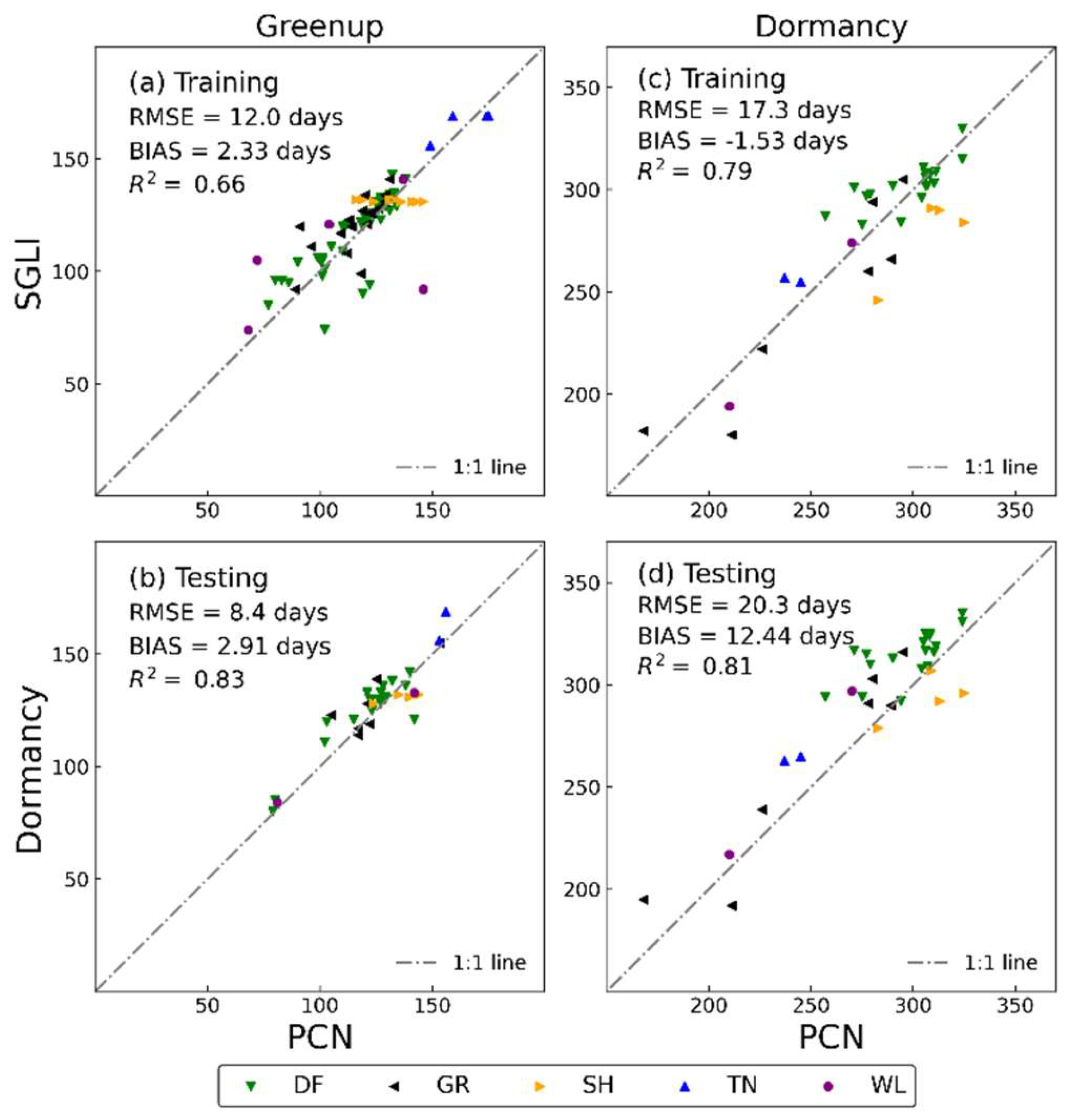
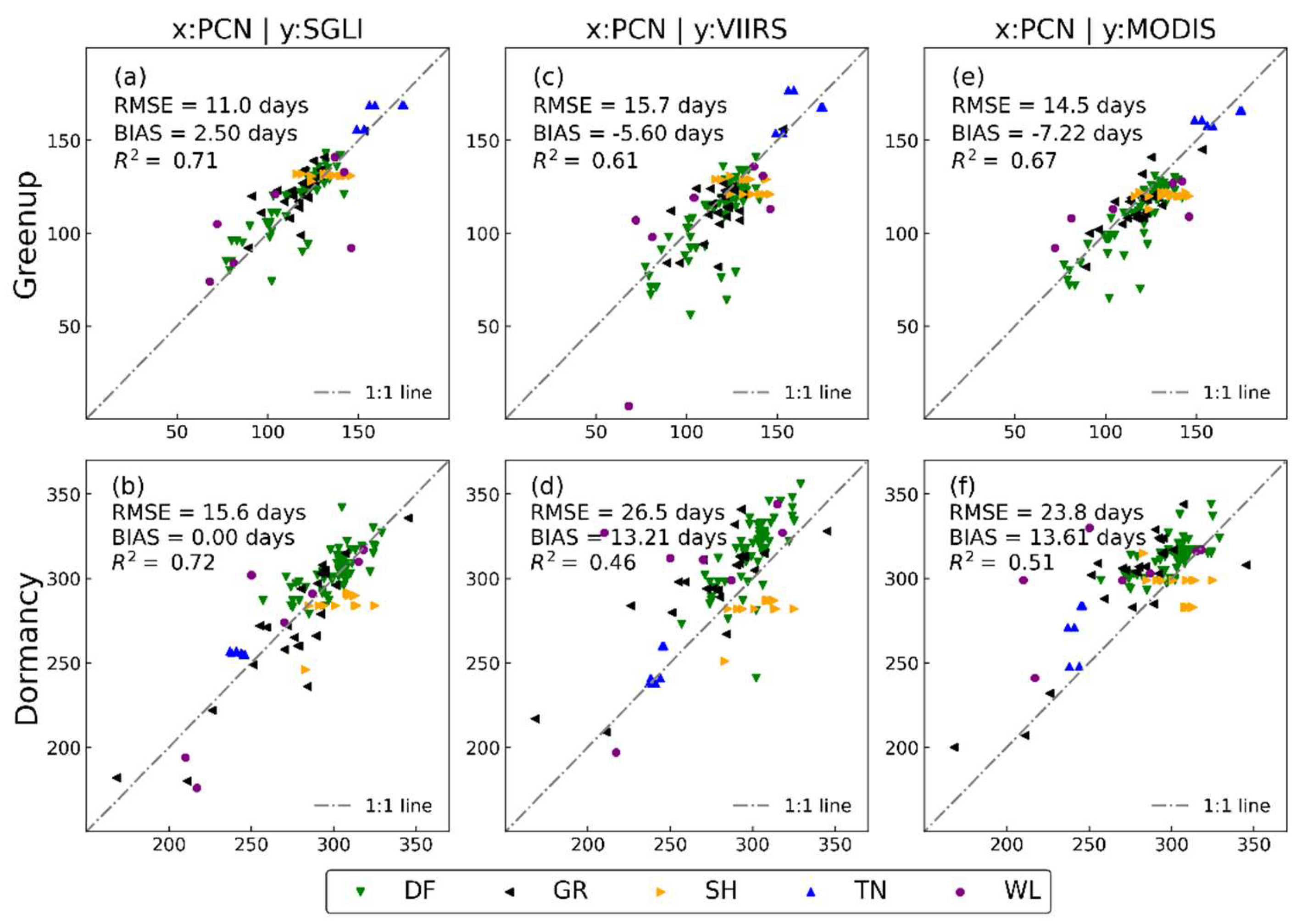
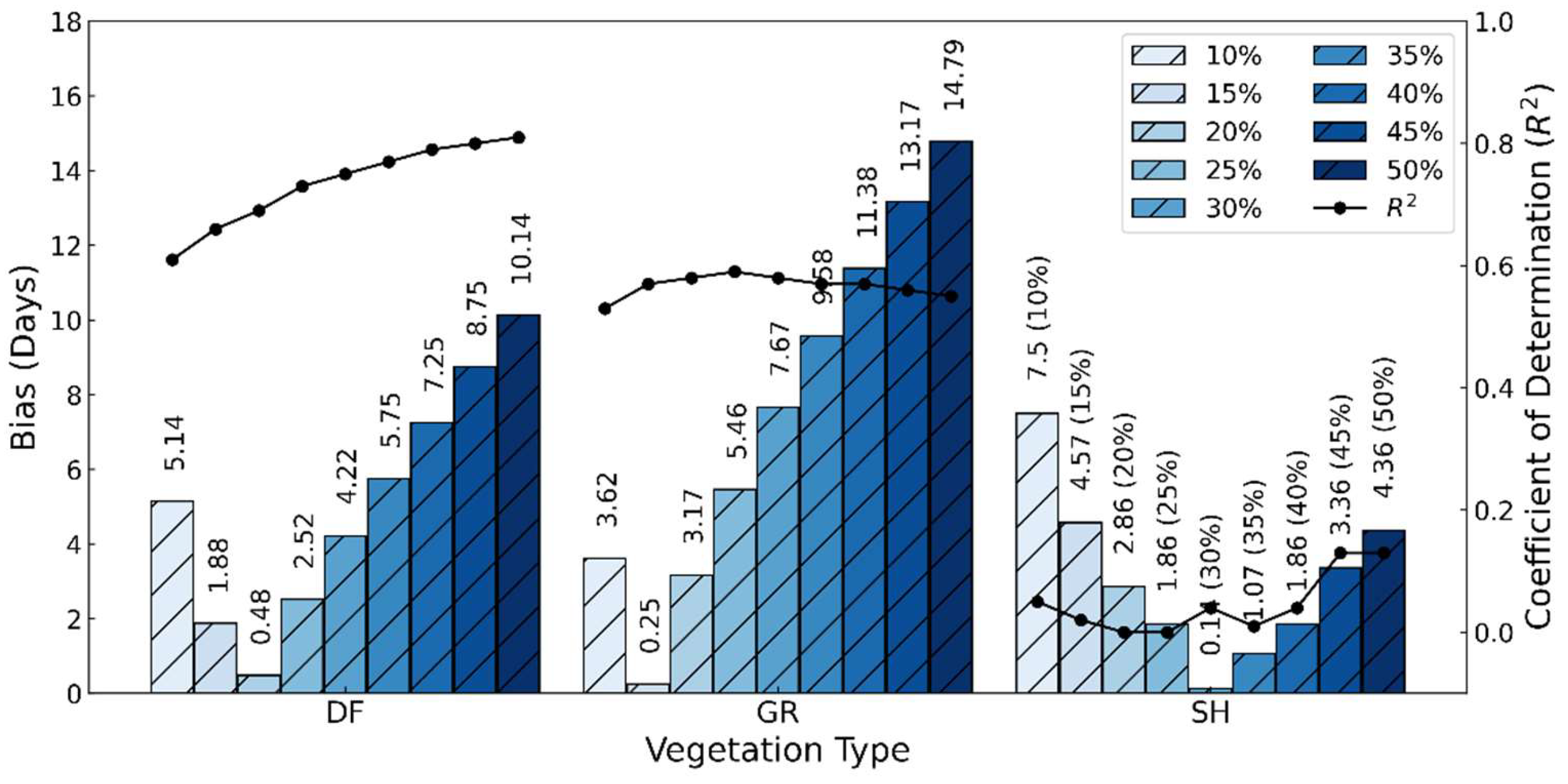
3.2. Comparison of Satellite Phenology with Near-Surface Phenology Observation
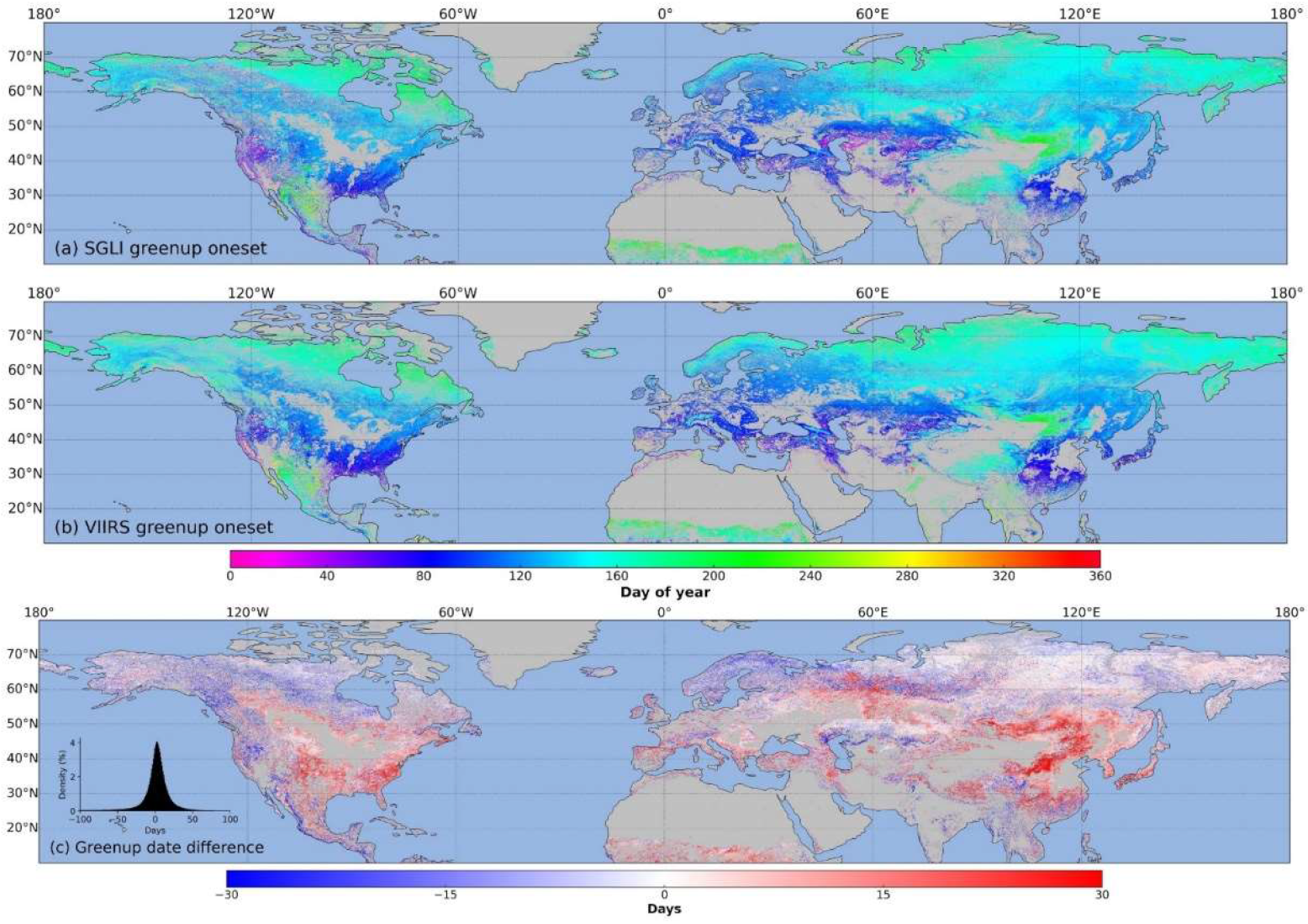
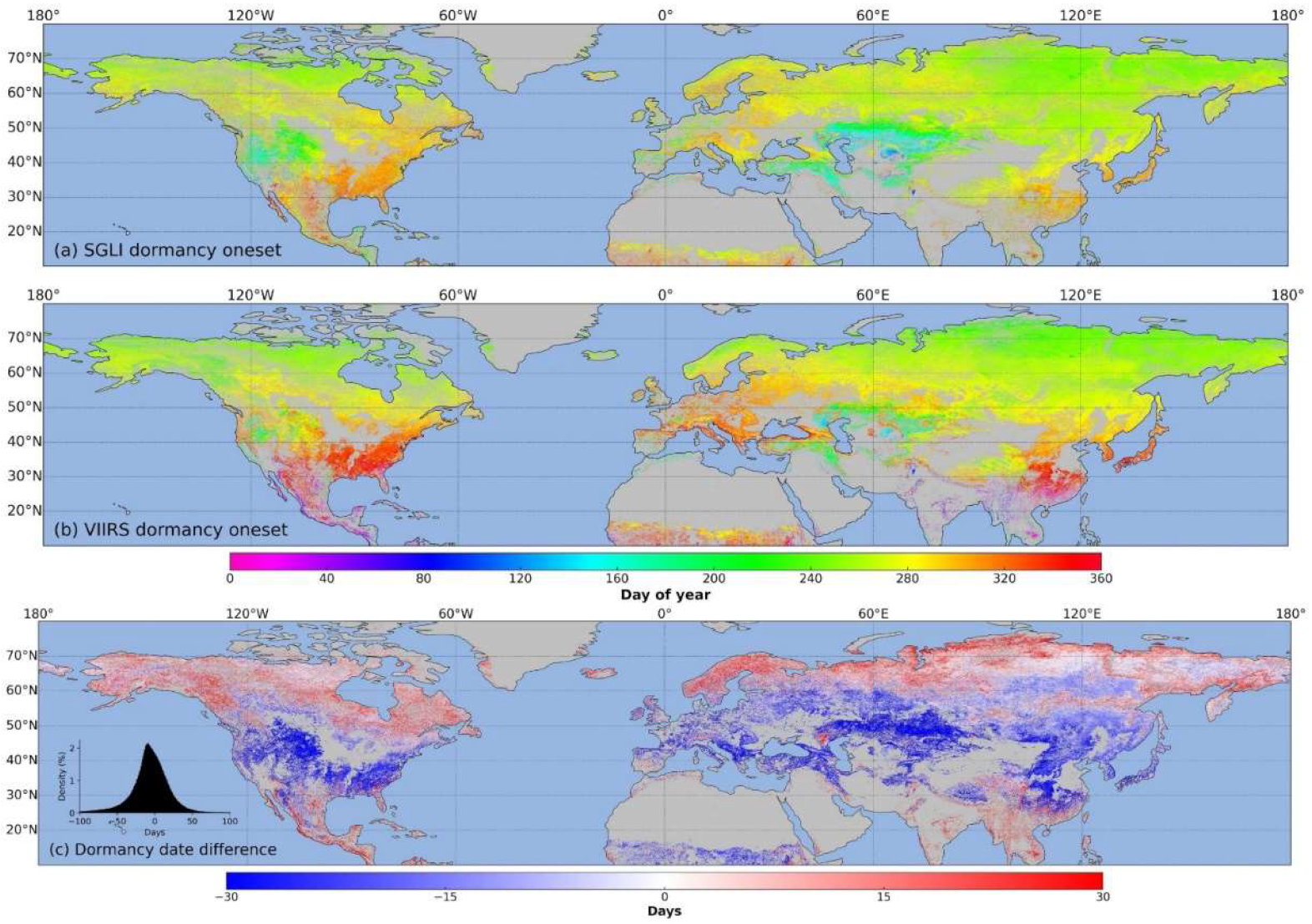
3.3. Spatial Distribution of SGLI Phenological Transition Dates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Badeck, F.W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of Spring Phenology to Climate Change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Peñuelas, J. Phenology Feedbacks on Climate Change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate Change, Phenology, and Phenological Control of Vegetation Feedbacks to the Climate System. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Sparks, T.H.; Carey, P.D. The Responses of Species to Climate Over Two Centuries: An Analysis of the Marsham Phenological Record, 1736–1947. J. Ecol. 1995, 83, 321. [Google Scholar] [CrossRef]
- Richardson, A.D.; Bailey, A.S.; Denny, E.G.; Martin, C.W.; O’Keefe, J. Phenology of a Northern Hardwood Forest Canopy. Glob. Change Biol. 2006, 12, 1174–1188. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring Phenological Variability from Satellite Imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring Vegetation Phenology Using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, Interpretation, and Assessment of Spring Phenology in North America Estimated from Remote Sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Busetto, L.; Colombo, R.; Migliavacca, M.; Cremonese, E.; Meroni, M.; Galvagno, M.; Rossini, M.; Siniscalco, C.; Morra Di Cella, U.; Pari, E. Remote Sensing of Larch Phenological Cycle and Analysis of Relationships with Climate in the Alpine Region. Glob. Chang. Biol. 2010, 16, 2504–2517. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, L.; Liu, Y.; Jayavelu, S.; Wang, J.; Moon, M.; Henebry, G.M.; Friedl, M.A.; Schaaf, C.B. Generation and Evaluation of the VIIRS Land Surface Phenology Product. Remote Sens. Environ. 2018, 216, 212–229. [Google Scholar] [CrossRef]
- Tateishi, R.; Ebata, M. Analysis of Phenological Change Patterns Using 1982–2000 Advanced Very High Resolution Radiometer (AVHRR) Data. Int. J. Remote Sens. 2004, 25, 2287–2300. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, X.; Nagai, S.; Yu, Y.; Akitsu, T.; Nasahara, K.N.; Ide, R.; Maeda, T. Evaluating Land Surface Phenology from the Advanced Himawari Imager Using Observations from MODIS and the Phenological Eyes Network. Int. J. Appl. Earth Obs. Geoinf. 2019, 79, 71–83. [Google Scholar] [CrossRef]
- Ganguly, S.; Friedl, M.A.; Tan, B.; Zhang, X.; Verma, M. Land Surface Phenology from MODIS: Characterization of the Collection 5 Global Land Cover Dynamics Product. Remote Sens. Environ. 2010, 114, 1805–1816. [Google Scholar] [CrossRef]
- Gray, J.; Sulla-Menashe, D.; Friedl, M.A. User Guide to Collection 6 MODIS Land Cover Dynamics (MCD12Q2) Product; NASA EOSDIS Land Processes DAAC: Missoula, MT, USA, 2019; Volume 6, pp. 1–8. Available online: https://modis-land.gsfc.nasa.gov/pdf/MCD12Q2_Collection6_UserGuide.pdf (accessed on 16 June 2022).
- Zhang, X.; Friedl, M.A.; Henebry, G.M. VIIRS Global Land Surface Phenology Product User Guide; United States Geological Survey: Reston, VA, USA, 2017; Volume 22. Available online: https://lpdaac.usgs.gov/documents/637/VNP22_User_Guide_V1.pdf (accessed on 16 June 2022).
- Zhang, X.; Liu, L.; Yan, D. Comparisons of Global Land Surface Seasonality and Phenology Derived from AVHRR, MODIS, and VIIRS Data. J. Geophys. Res. Biogeosci. 2017, 122, 1506–1525. [Google Scholar] [CrossRef]
- Li, L.; Li, X.; Asrar, G.; Zhou, Y.; Chen, M.; Zeng, Y.; Li, X.; Li, F.; Luo, M.; Sapkota, A.; et al. Detection and Attribution of Long-Term and Fine-Scale Changes in Spring Phenology over Urban Areas: A Case Study in New York State. Int. J. Appl. Earth Obs. Geoinf. 2022, 110, 102815. [Google Scholar] [CrossRef]
- Misra, G.; Cawkwell, F.; Wingler, A. Status of Phenological Research Using Sentinel-2 Data: A Review. Remote Sens. 2020, 12, 2760. [Google Scholar] [CrossRef]
- Wheeler, K.I.; Dietze, M.C. Improving the Monitoring of Deciduous Broadleaf Phenology Using the Geostationary Operational Environmental Satellite (GOES) 16 and 17. Biogeosciences 2021, 18, 1971–1985. [Google Scholar] [CrossRef]
- Xin, Q.; Li, J.; Li, Z.; Li, Y.; Zhou, X. Evaluations and Comparisons of Rule-Based and Machine-Learning-Based Methods to Retrieve Satellite-Based Vegetation Phenology Using MODIS and USA National Phenology Network Data. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102189. [Google Scholar] [CrossRef]
- Stanimirova, R.; Cai, Z.; Melaas, E.K.; Gray, J.M.; Eklundh, L.; Jönsson, P.; Friedl, M.A. An Empirical Assessment of the MODIS Land Cover Dynamics and TIMESAT Land Surface Phenology Algorithms. Remote Sens. 2019, 11, 2201. [Google Scholar] [CrossRef]
- Jin, H.; Eklundh, L. A Physically Based Vegetation Index for Improved Monitoring of Plant Phenology. Remote Sens. Environ. 2014, 152, 512–525. [Google Scholar] [CrossRef]
- Wang, C.; Chen, J.; Wu, J.; Tang, Y.; Shi, P.; Black, T.A.; Zhu, K. A Snow-Free Vegetation Index for Improved Monitoring of Vegetation Spring Green-up Date in Deciduous Ecosystems. Remote Sens. Environ. 2017, 196, 1–12. [Google Scholar] [CrossRef]
- Delbart, N.; Kergoat, L.; le Toan, T.; Lhermitte, J.; Picard, G. Determination of Phenological Dates in Boreal Regions Using Normalized Difference Water Index. Remote Sens. Environ. 2005, 97, 26–38. [Google Scholar] [CrossRef]
- Cao, R.; Feng, Y.; Liu, X.; Shen, M.; Zhou, J. Uncertainty of Vegetation Green-up Date Estimated from Vegetation Indices Due to Snowmelt at Northern Middle and High Latitudes. Remote Sens. 2020, 12, 190. [Google Scholar] [CrossRef]
- Yang, W.; Kobayashi, H.; Wang, C.; Shen, M.; Chen, J.; Matsushita, B.; Tang, Y.; Kim, Y.; Bret-Harte, M.S.; Zona, D.; et al. A Semi-Analytical Snow-Free Vegetation Index for Improving Estimation of Plant Phenology in Tundra and Grassland Ecosystems. Remote Sens. Environ. 2019, 228, 31–44. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yokozawa, M.; Toritani, H.; Shibayama, M.; Ishitsuka, N.; Ohno, H. A Crop Phenology Detection Method Using Time-Series MODIS Data. Remote Sens. Environ. 2005, 96, 366–374. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. Seasonality Extraction by Function Fitting to Time-Series of Satellite Sensor Data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Shang, R.; Liu, R.; Xu, M.; Liu, Y.; Zuo, L.; Ge, Q. The Relationship between Threshold-Based and Inflexion-Based Approaches for Extraction of Land Surface Phenology. Remote Sens. Environ. 2017, 199, 167–170. [Google Scholar] [CrossRef]
- Huang, X.; Liu, J.; Zhu, W.; Atzberger, C.; Liu, Q. The Optimal Threshold and Vegetation Index Time Series for Retrieving Crop Phenology Based on a Modified Dynamic Threshold Method. Remote Sens. 2019, 11, 2725. [Google Scholar] [CrossRef]
- Tian, F.; Cai, Z.; Jin, H.; Hufkens, K.; Scheifinger, H.; Tagesson, T.; Smets, B.; van Hoolst, R.; Bonte, K.; Ivits, E.; et al. Calibrating Vegetation Phenology from Sentinel-2 Using Eddy Covariance, PhenoCam, and PEP725 Networks across Europe. Remote Sens. Environ. 2021, 260, 112456. [Google Scholar] [CrossRef]
- White, M.A.; Nemani, R.R. Real-Time Monitoring and Short-Term Forecasting of Land Surface Phenology. Remote Sens. Environ. 2006, 104, 43–49. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Aubrecht, D.M.; Chen, M.; Gray, J.M.; Johnston, M.R.; Keenan, T.F.; Klosterman, S.T.; Kosmala, M.; et al. Tracking Vegetation Phenology across Diverse North American Biomes Using PhenoCam Imagery. Sci. Data 2018, 5, 180028. [Google Scholar] [CrossRef]
- Nasahara, K.N.; Nagai, S. Review: Development of an in Situ Observation Network for Terrestrial Ecological Remote Sensing: The Phenological Eyes Network (PEN). Ecol. Res. 2015, 30, 211–223. [Google Scholar] [CrossRef]
- Zhang, X.; Jayavelu, S.; Liu, L.; Friedl, M.A.; Henebry, G.M.; Liu, Y.; Schaaf, C.B.; Richardson, A.D.; Gray, J. Evaluation of Land Surface Phenology from VIIRS Data Using Time Series of PhenoCam Imagery. Agric. For. Meteorol. 2018, 256–257, 137–149. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating Remote Sensing of Deciduous Forest Phenology at Multiple Spatial Scales Using PhenoCam Imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef]
- Imaoka, K.; Kachi, M.; Fujii, H.; Murakami, H.; Hori, M.; Ono, A.; Igarashi, T.; Nakagawa, K.; Oki, T.; Honda, Y.; et al. Global Change Observation Mission (GCOM) for Monitoring Carbon, Water Cycles, and Climate Change. Proc. IEEE 2010, 98, 717–734. [Google Scholar] [CrossRef]
- Murakami, H. GCOM-C/SGLI Land Atmospheric Correction Algorithm; Japan Aerospace Exploration Agency: Tokyo, Japan, 2018; Volume 2018, pp. 1–12. Available online: https://suzaku.eorc.jaxa.jp/GCOM_C/data/ATBD/ver1/SGLI_Atmcorr_ATBD_v10.pdf (accessed on 16 June 2022).
- Ma, M.; Veroustraete, F. Reconstructing Pathfinder AVHRR Land NDVI Time-Series Data for the Northwest of China. Adv. Space Res. 2006, 37, 835–840. [Google Scholar] [CrossRef]
- Viovy, N.; Arino, O.; Belward, A.S. The Best Index Slope Extraction (BISE): A Method for Reducing Noise in NDVI Time-Series. Int. J. Remote Sens. 1992, 13, 1585–1590. [Google Scholar] [CrossRef]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 Global Land Cover: Algorithm Refinements and Characterization of New Datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Richards, F.J. A Flexible Growth Function for Empirical Use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H.J. A Flexible Sigmoid Function of Determinate Growth. Ann. Bot. 2003, 91, 361–371. [Google Scholar] [CrossRef]
- Seyednasrollah, B.; Young, A.M.; Hufkens, K.; Milliman, T.; Friedl, M.A.; Frolking, S.; Richardson, A.D. Tracking Vegetation Phenology across Diverse Biomes Using Version 2.0 of the PhenoCam Dataset. Sci. Data 2019, 6, 222. [Google Scholar] [CrossRef]
- Zhang, X.; Mark, A.; Friedl, G.M.H. VIIRS/NPP Land Cover Dynamics Yearly L3 Global 500m SIN Grid V001. Available online: https://lpdaac.usgs.gov/products/vnp22q2v001/ (accessed on 16 June 2022).
- Verger, A.; Filella, I.; Baret, F.; Peñuelas, J. Vegetation Baseline Phenology from Kilometric Global LAI Satellite Products. Remote Sens. Environ. 2016, 178, 1–14. [Google Scholar] [CrossRef]
- Fridley, J.D. Extended Leaf Phenology and the Autumn Niche in Deciduous Forest Invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green Leaf Phenology at Landsat Resolution: Scaling from the Field to the Satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Frolking, S. Intercomparison of Phenological Transition Dates Derived from the PhenoCam Dataset V1.0 and MODIS Satellite Remote Sensing. Sci. Rep. 2018, 8, 5679. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Threshold (%) | RMSE (Days) | Bias (Days) | R2 | Phase | Threshold (%) | RMSE (Days) | Bias (Days) | R2 |
|---|---|---|---|---|---|---|---|---|---|
| Green-up date | 10 | 13.9 | −5.51 | 0.63 | Dormancy date | 10 | 38.8 | 32.77 | 0.55 |
| 15 | 11.9 | −2.05 | 0.67 | 15 | 31.3 | 25.13 | 0.61 | ||
| 20 | 11.2 | 0.47 | 0.69 | 20 | 26.1 | 19.43 | 0.66 | ||
| 25 | 11.0 | 2.50 | 0.71 | 25 | 22.2 | 14.71 | 0.68 | ||
| 30 | 11.4 | 4.40 | 0.72 | 30 | 19.2 | 10.61 | 0.70 | ||
| 35 | 11.9 | 6.03 | 0.73 | 35 | 17.1 | 6.78 | 0.71 | ||
| 40 | 12.7 | 7.57 | 0.73 | 40 | 16.0 | 3.37 | 0.72 | ||
| 45 | 13.6 | 9.16 | 0.74 | 45 | 15.6 | 0.00 | 0.72 | ||
| 50 | 14.6 | 10.61 | 0.74 | 50 | 16.1 | −3.26 | 0.72 |
| Green-Up | Statistic | DF (n = 67) | GR (n = 24) | SH (n = 14) | TN (n = 6) | WL (n = 7) | |
|---|---|---|---|---|---|---|---|
| PhenoCan network GCC | SGLI NDGI | RMSE ± SD | 9.1 ± 8.8 | 10.5 ± 9.0 | 9.3 ± 9.1 | 8.0 ± 7.2 | 25.2 ± 25.2 |
| R2 | 0.73 ** | 0.59 ** | 0.00 | 0.50 | 0.39 | ||
| Bias | 2.52 | 3.17 | −2.86 | 0.67 | −2.43 | ||
| VIIRS EVI2 | RMSE ± SD | 14.8 ± 13.2 | 12.9 ± 12.2 | 14.1 ± 11.7 | 12.1 ± 10.8 | 30.9 ± 30.4 | |
| R2 | 0.57 ** | 0.43 ** | 0.23 * | 0.15 | 0.45 * | ||
| Bias | −6.58 | −4.38 | −7.93 | 5.33 | −5.57 | ||
| MODIS EVI2 | RMSE ± SD | 12.3 ± 9.4 | 8.8 ± 8.5 | 15.7 ± 9.0 | 7.7 ± 7.7 | 35.5 ± 33.5 | |
| R2 | 0.70 ** | 0.62 ** | 0.04 | 0.62 * | 0.43 | ||
| Bias | −8.05 | −2.38 | −12.86 | 0.67 | −11.71 | ||
| Dormancy | Statistic | DF (n = 67) | GR (n = 24) | SH (n = 14) | TN (n = 6) | WL (n = 7) | |
| PhenoCan network GCC | SGLI NDGI | RMSE ± SD | 11.1 ± 10.2 | 16.6 ± 16.3 | 23.3 ± 10.6 | 14.7 ± 4.1 | 25.9 ± 25.9 |
| R2 | 0.52 ** | 0.81 ** | 0.32 ** | 0.6 * | 0.78 ** | ||
| Bias | 4.31 | −2.83 | −20.71 | 14.17 | −0.43 | ||
| VIIRS EVI2 | RMSE ± SD | 22.0 ± 14.8 | 28.8 ± 19.1 | 24.9 ± 9.9 | 8.7 ± 7.4 | 54.4 ± 41.0 | |
| R2 | 0.46 ** | 0.69 ** | 0.32 ** | 0.61 * | 0.30 | ||
| Bias | 16.36 | 21.46 | −22.86 | 4.5 | 35.71 | ||
| MODIS EVI2 | RMSE ± SD | 16.4 ± 11.0 | 29.4 ± 18.9 | 21.2 ± 18.5 | 29.3 ± 13.7 | 47.8 ± 33.6 | |
| R2 | 0.42 ** | 0.73 ** | 0.26 * | 0.19 | 0.31 | ||
| Bias | 12.16 | 22.46 | −10.36 | 25.83 | 34 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Yang, W.; Kondoh, A. Improving Remote Estimation of Vegetation Phenology Using GCOM-C/SGLI Land Surface Reflectance Data. Remote Sens. 2022, 14, 4027. https://doi.org/10.3390/rs14164027
Li M, Yang W, Kondoh A. Improving Remote Estimation of Vegetation Phenology Using GCOM-C/SGLI Land Surface Reflectance Data. Remote Sensing. 2022; 14(16):4027. https://doi.org/10.3390/rs14164027
Chicago/Turabian StyleLi, Mengyu, Wei Yang, and Akihiko Kondoh. 2022. "Improving Remote Estimation of Vegetation Phenology Using GCOM-C/SGLI Land Surface Reflectance Data" Remote Sensing 14, no. 16: 4027. https://doi.org/10.3390/rs14164027
APA StyleLi, M., Yang, W., & Kondoh, A. (2022). Improving Remote Estimation of Vegetation Phenology Using GCOM-C/SGLI Land Surface Reflectance Data. Remote Sensing, 14(16), 4027. https://doi.org/10.3390/rs14164027