
Physical and Biochemical Responses to Sequential Tropical Cyclones in the Arabian Sea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Remote Sensing Data
2.2. In Situ Observations
2.3. Model Data
2.4. Methods
2.4.1. Ekman Pumping Velocity
2.4.2. Mixed Layer Depth and Euphotic Layer Depth
2.4.3. Wind Work and Forcing Time
3. Results
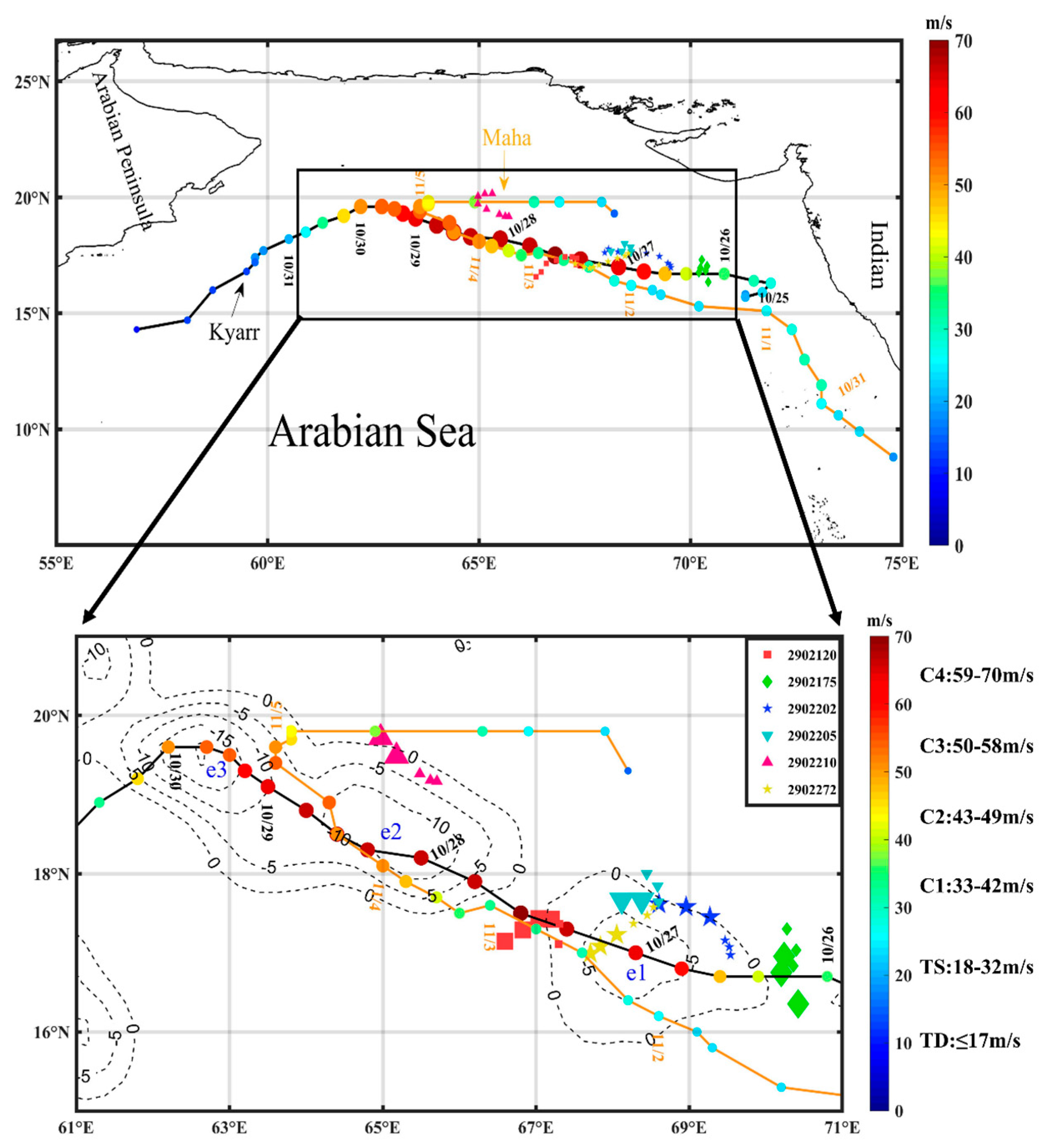
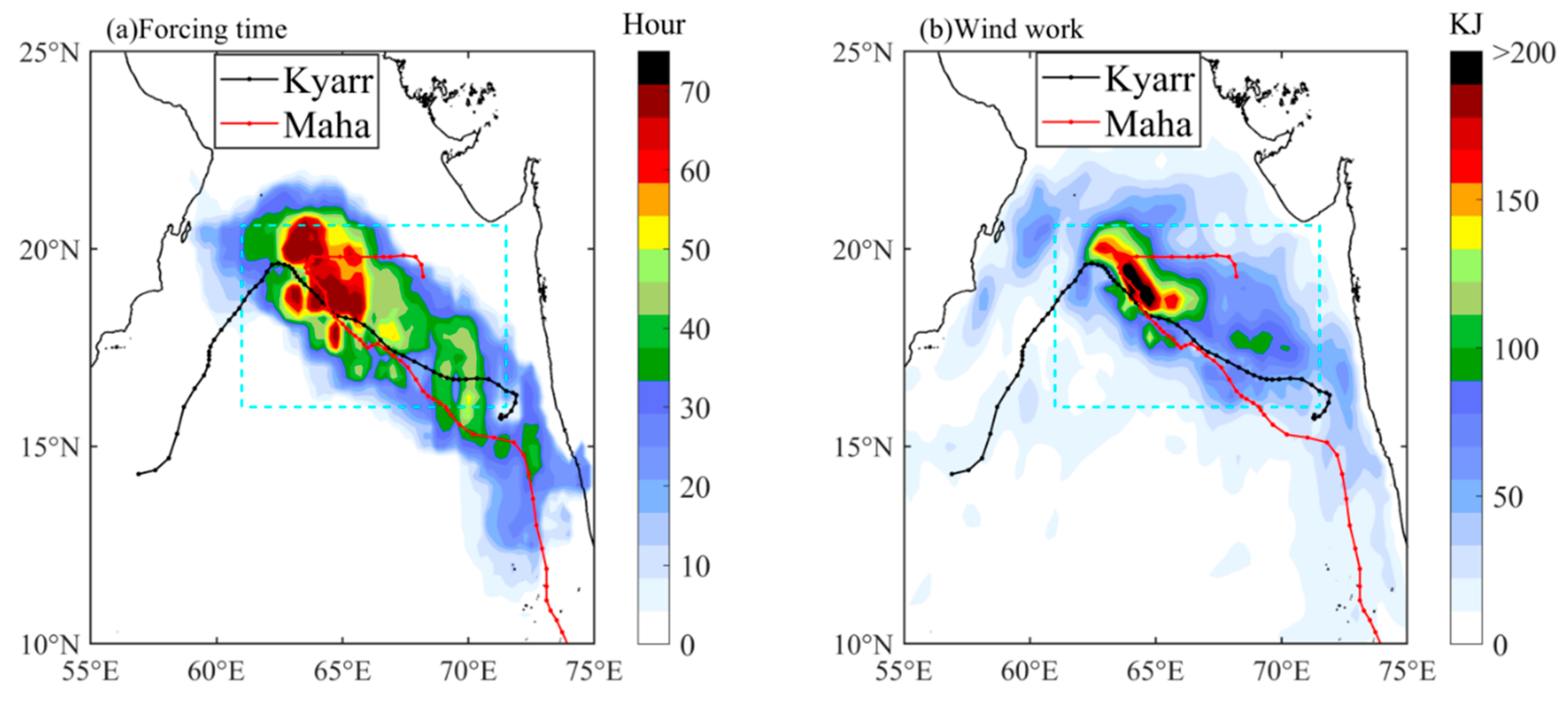
3.1. Sequential Tropical Cyclones and the Study Area
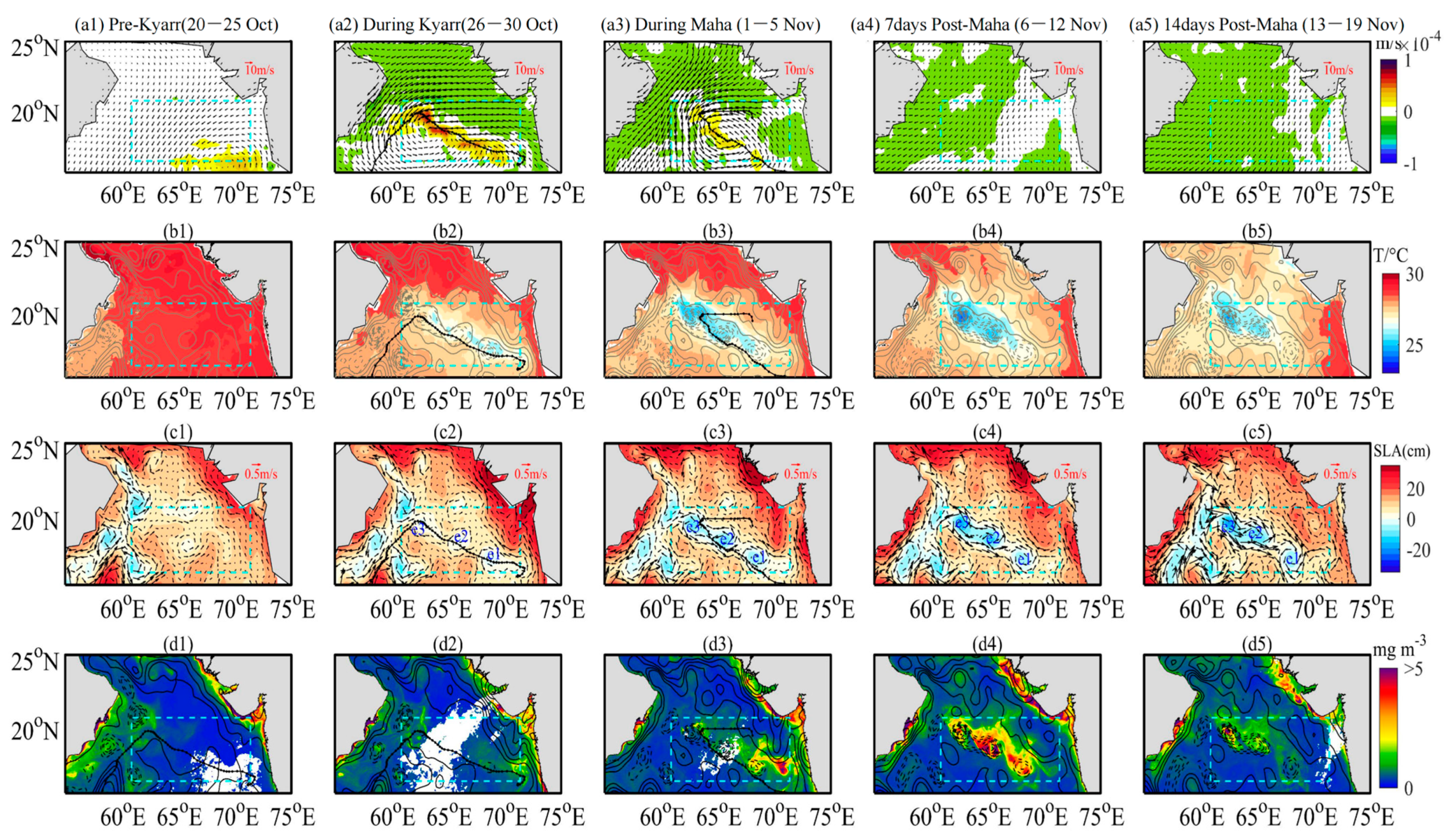
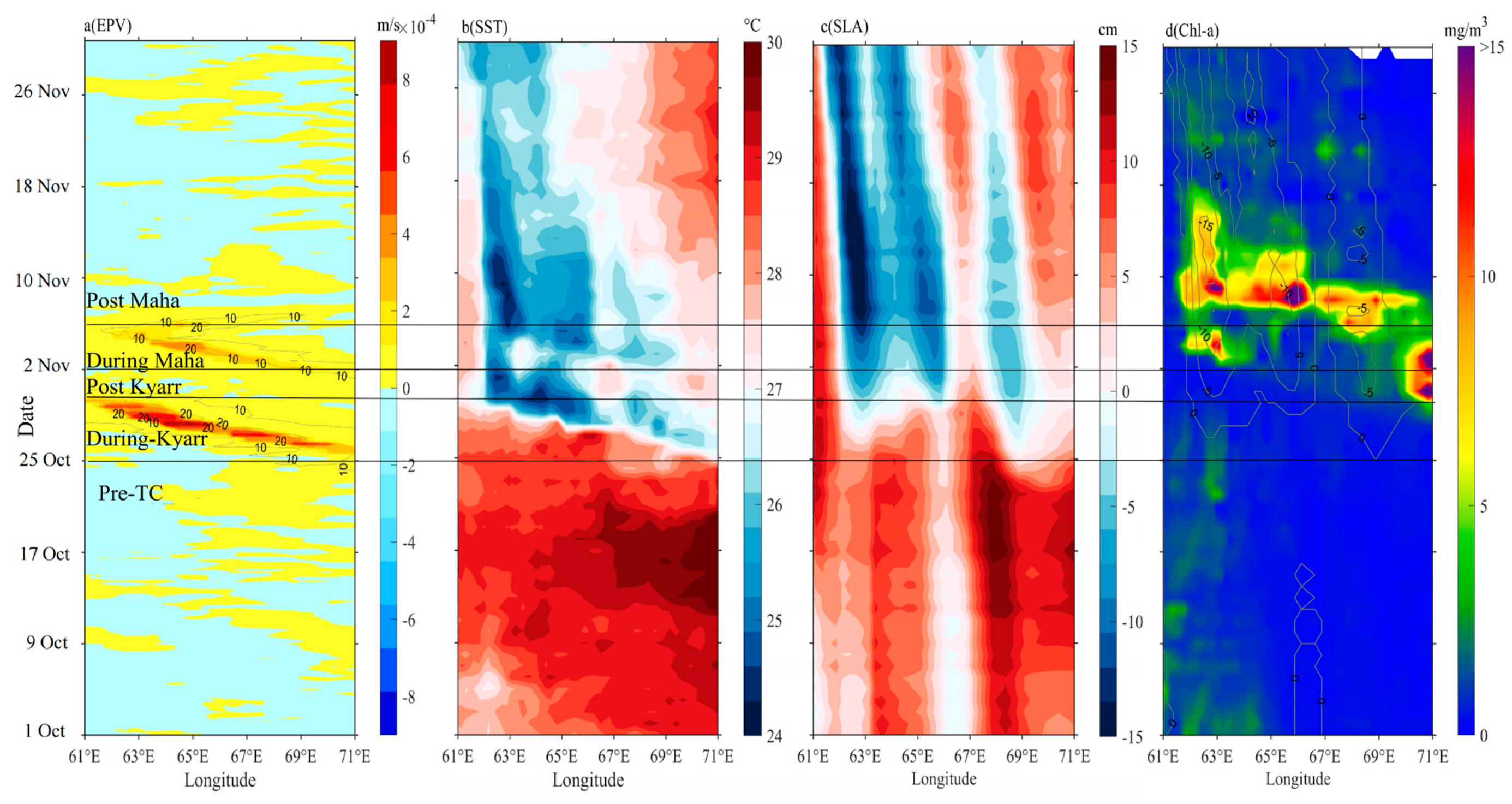
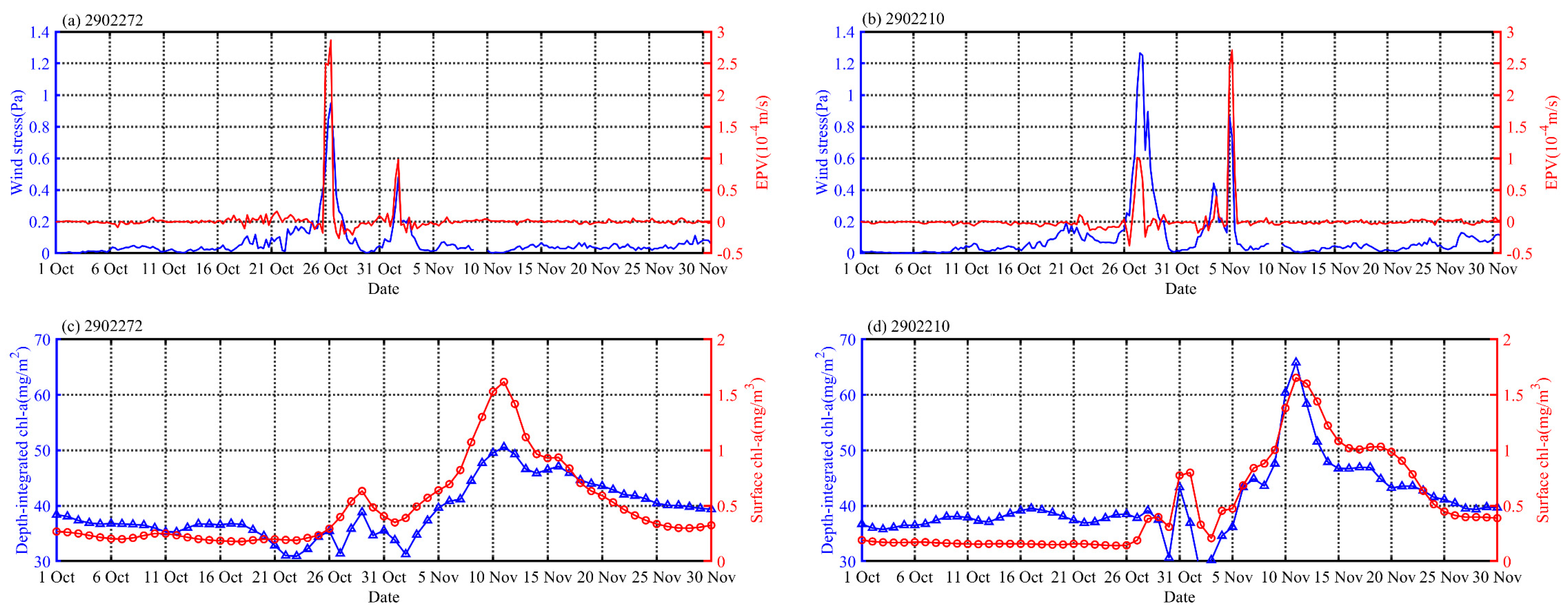
3.2. Horizontal Distributions of EPV, SST, SLA and Chl-a
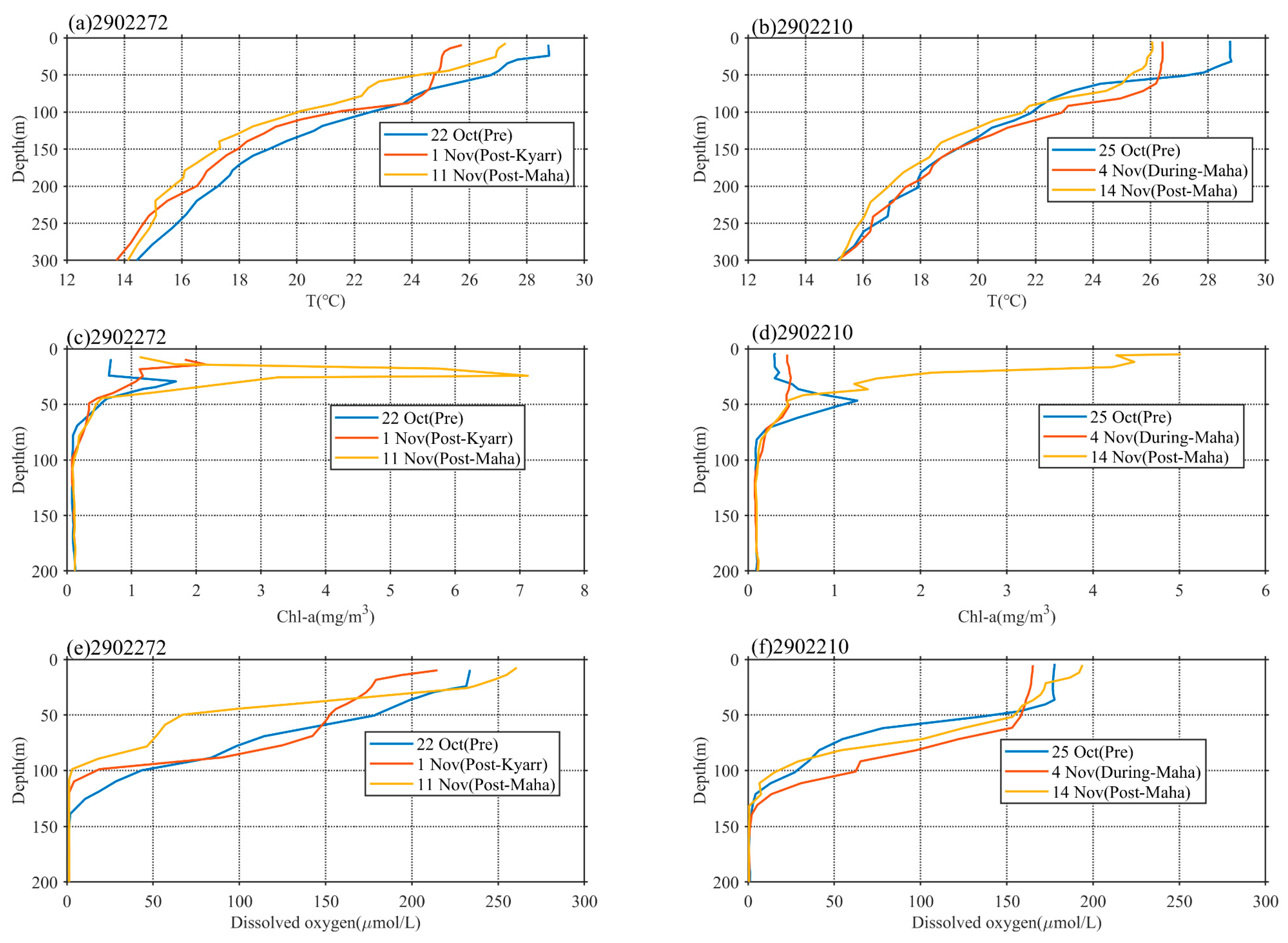
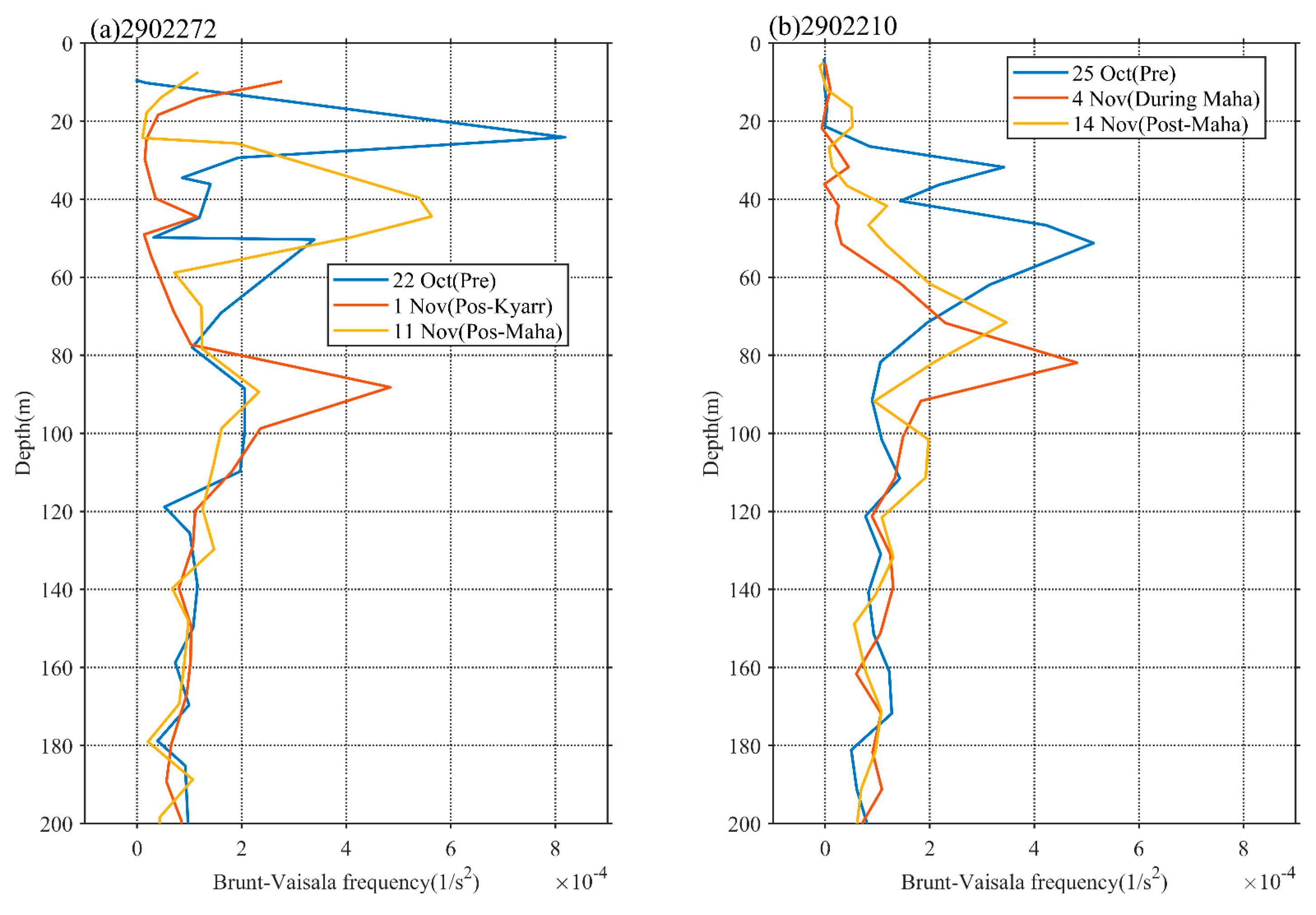
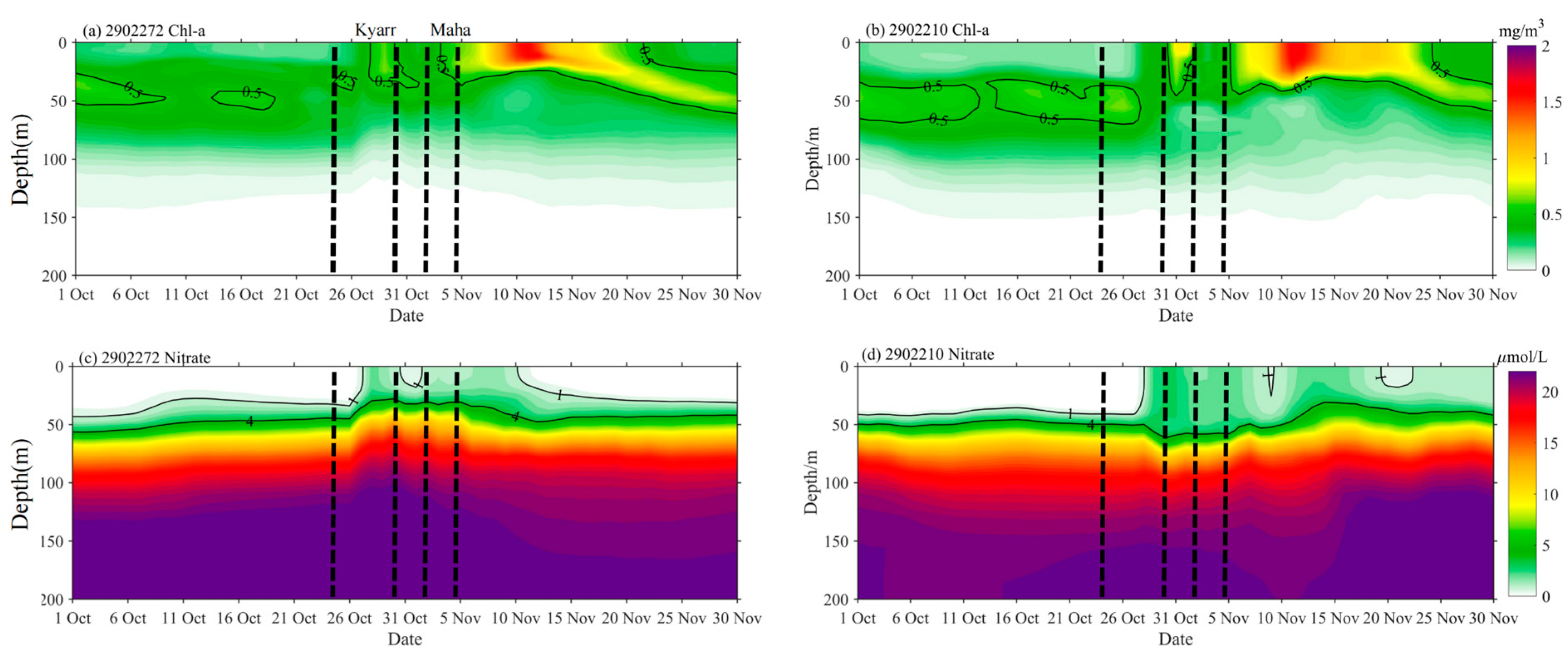
3.3. Vertical Distributions of Temperature, Chl-a, and DO
4. Discussion
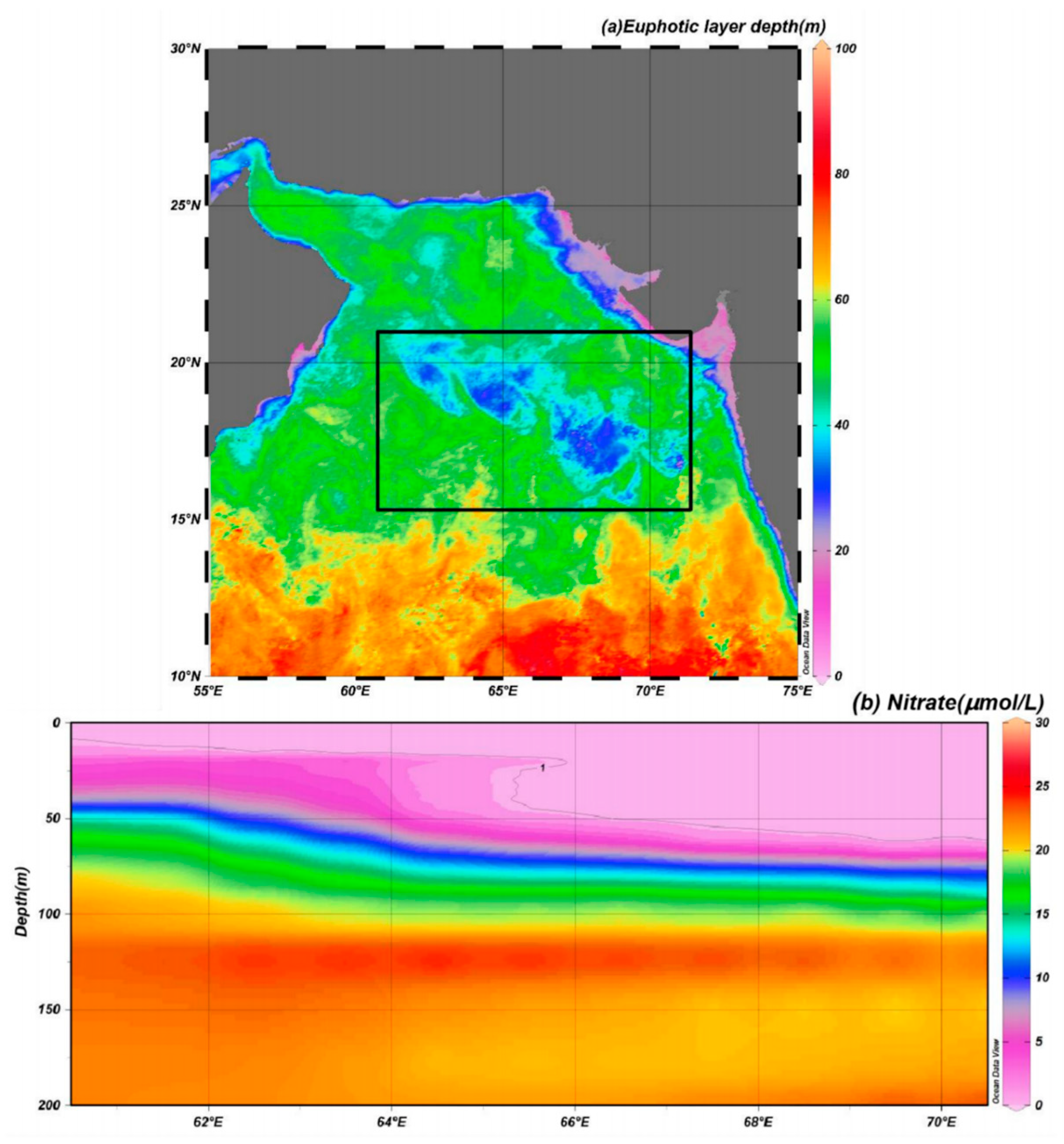
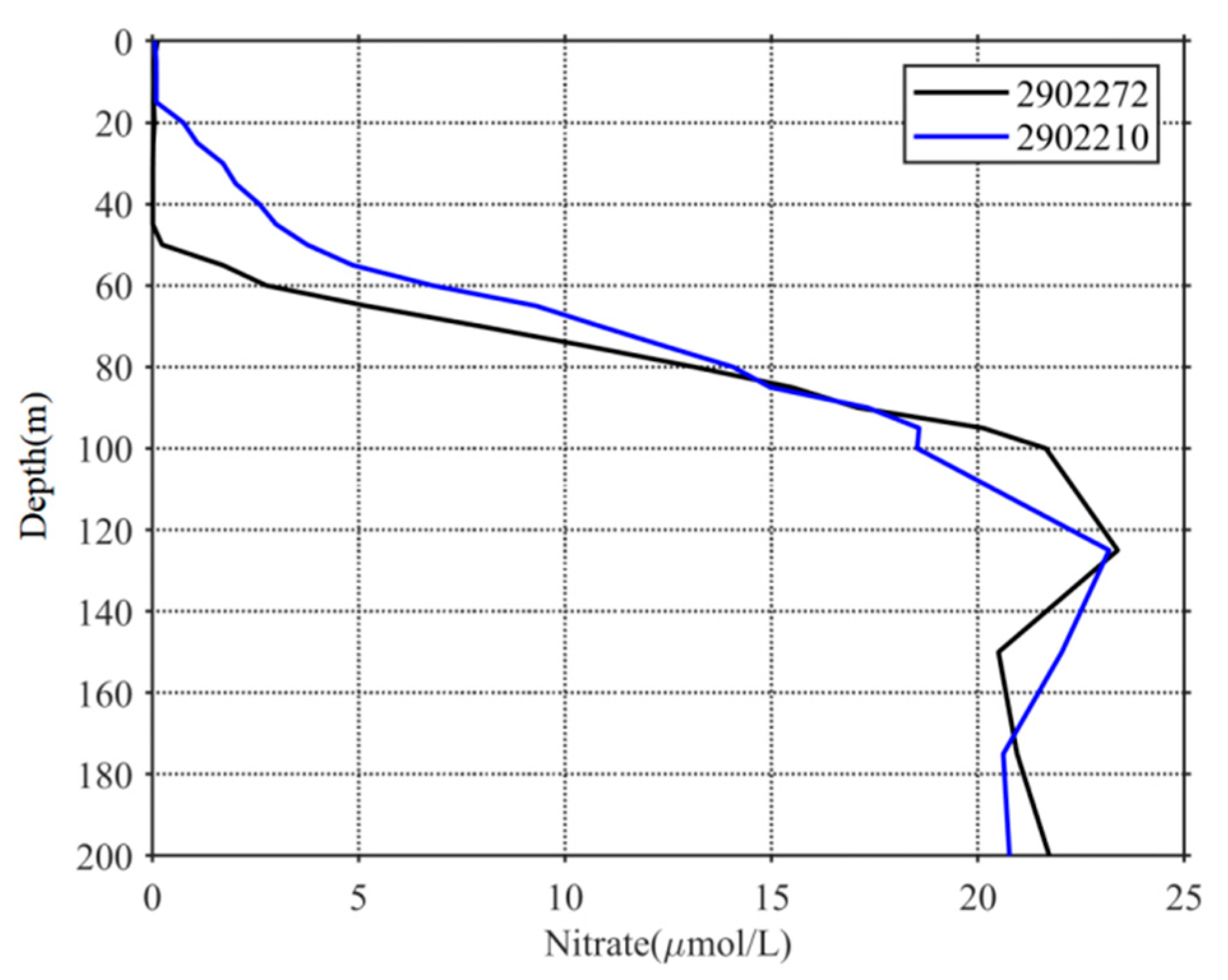
4.1. Light and Nutrient Conditions in the Euphotic Layer before the TCs
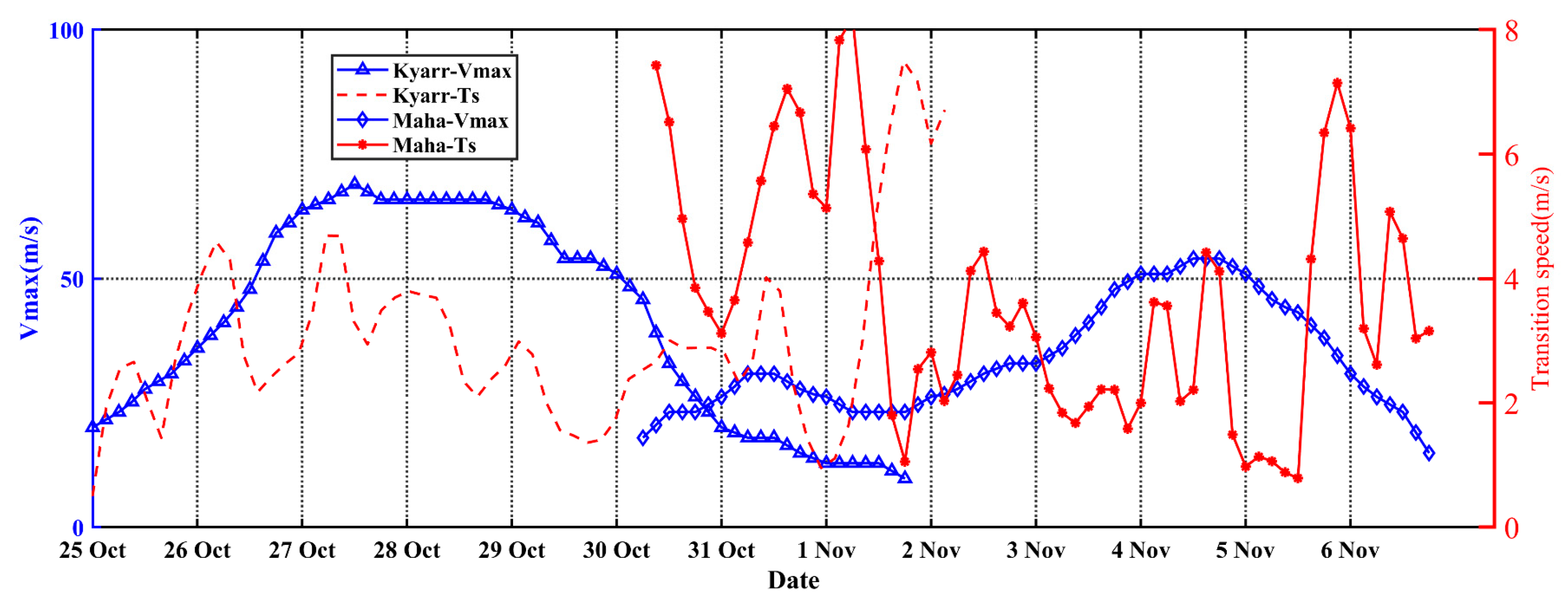
4.2. The Translation Speed and Wind Speed of TCs Kyarr and Maha
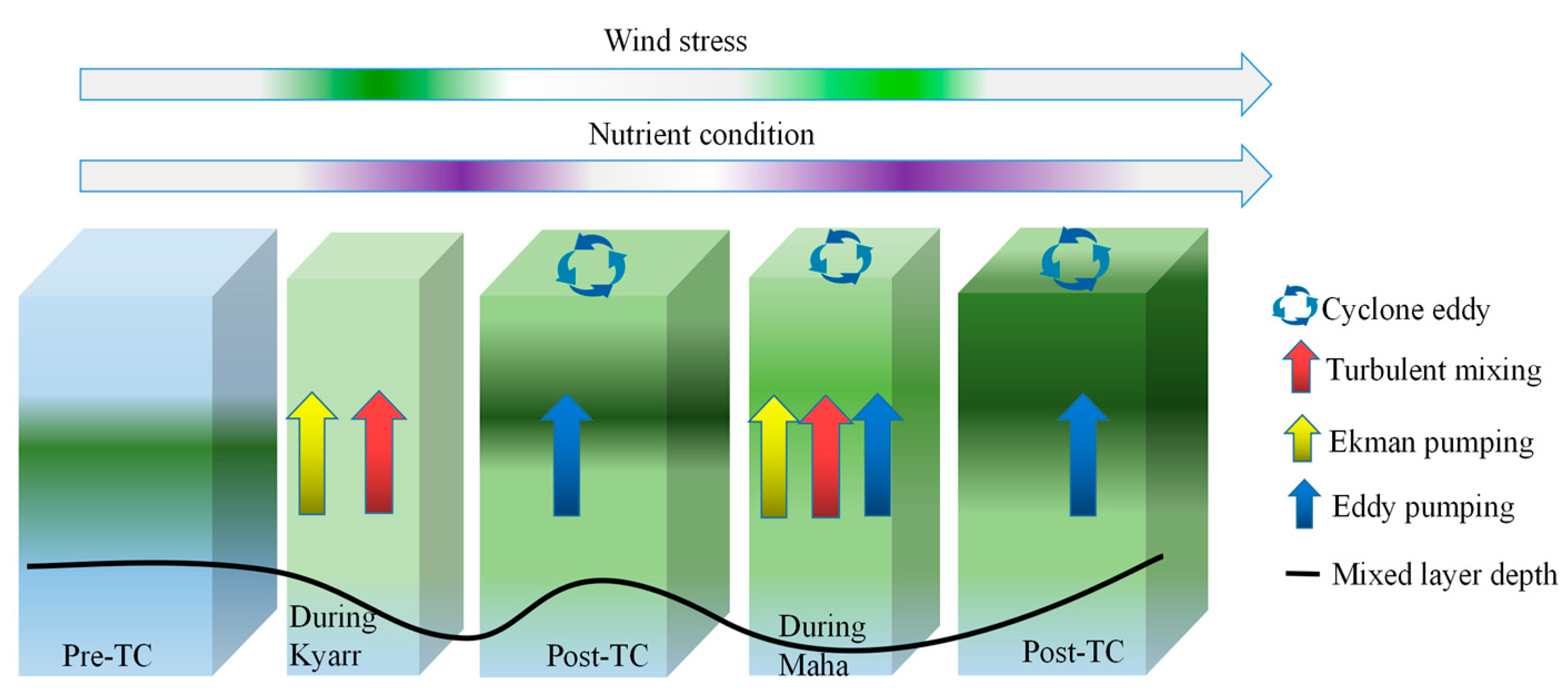
4.3. Impact of Physical Processes on Phytoplankton Enhancement in the Upper Ocean
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Price, J.F. Upper Ocean response to a hurricane. J. Phys. Oceanogr. 1981, 11, 153–175. [Google Scholar] [CrossRef] [Green Version]
- Chu, P.C.; Veneziano, J.M.; Fan, C.; Carron, M.J.; Liu, W.T. Response of the South China Sea to tropical cyclone Ernie 1996. J. Geophys. Res. Ocean. 2000, 105, 13991–14009. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Yan, X.H. Hurricane forcing on chlorophyll-a concentration off the northeast coast of the US. Geophys. Res. Lett. 2004, 31, L17304. [Google Scholar] [CrossRef]
- Sun, L.; Li, Y.X.; Yang, Y.J.; Wu, Q.; Chen, X.T.; Li, Q.Y.; Xian, T. Effects of super typhoons on cyclonic ocean eddies in the western North Pacific: A satellite data-based evaluation between 2000 and 2008. J. Geophys. Res. Ocean. 2014, 119, 5585–5598. [Google Scholar] [CrossRef]
- Price, J.F.; Sanford, T.B.; Forristall, G.Z. Forced stage response to a moving hurricane. J. Phys. Oceanogr. 1994, 24, 233–260. [Google Scholar] [CrossRef]
- Wentz, F.J.; Gentemann, C.; Smith, D.; Chelton, D. Satellite measurements of sea surface temperature through clouds. Science 2000, 288, 847–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, W.J.; Dickey, T.D. Observations and analyses of upper ocean responses to tropical storms and hurricanes in the vicinity of Bermuda. J. Geophys. Res. Ocean. 2008, 113, C08009. [Google Scholar] [CrossRef] [Green Version]
- D’Asaro, E.A.; Black, P.G.; Centurioni, L.R.; Chang, Y.T.; Chen, S.S.; Foster, R.C.; Lin, I.I. Impact of typhoons on the ocean in the Pacific. Bull. Am. Meteorol. Soc. 2014, 95, 1405–1418. [Google Scholar] [CrossRef]
- Chiang, T.L.; Wu, C.R.; Oey, L.Y. Typhoon Kai-Tak: An ocean’s perfect storm. J. Phys. Oceanogr. 2011, 41, 221–233. [Google Scholar] [CrossRef]
- Soloviev, A.V.; Lukas, R.; Donelan, M.A.; Haus, B.K.; Ginis, I. The air-sea interface and surface stress under tropical cyclones. Sci. Rep. 2014, 4, 5306. [Google Scholar] [CrossRef] [PubMed]
- Soloviev, A.V.; Lukas, R.; Donelan, M.A.; Haus, B.K.; Ginis, I. Is the state of the air-sea interface a factor in rapid intensification and rapid decline of tropical cyclones? J. Geophys. Res. Ocean. 2017, 122, 10174–10183. [Google Scholar] [CrossRef]
- Park, J.H.; Yeo, D.E.; Lee, K.; Lee, H.; Lee, S.W.; Noh, S.; Nam, S. Rapid decay of slowly moving Typhoon Soulik (2018) due to interactions with the strongly stratified northern East China Sea. Geophys. Res. Lett. 2019, 46, 14595–14603. [Google Scholar] [CrossRef] [Green Version]
- Shay, L.K.; Uhlhorn, E.W. Loop current response to Hurricanes Isidore and Lili. Mon. Weather. Rev. 2008, 136, 3248–3274. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zheng, H.; Xie, L.; Zheng, Q.; Ling, Z.; Li, M. Response of Total Suspended Sediment and Chlorophyll-a Concentration to Late Autumn Typhoon Events in the Northwestern South China Sea. Remote Sens. 2021, 13, 2863. [Google Scholar] [CrossRef]
- Naik, H.; Naqvi, S.W.A.; Suresh, T.; Narvekar, P.V. Impact of a tropical cyclone on biogeochemistry of the central Arabian Sea. Glob. Biogeochem. Cycles 2008, 22, GB3020. [Google Scholar] [CrossRef]
- Byju, P.; Kumar, S.P. Physical and biological response of the Arabian Sea to tropical cyclone Phyan and its implications. Mar. Environ. Res. 2011, 71, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaube, P.; Chelton, D.B.; Strutton, P.G.; Behrenfeld, M.J. Satellite observations of chlorophyll, phytoplankton biomass, and Ekman pumping in nonlinear mesoscale eddies. J. Geophys. Res. Ocean. 2013, 118, 6349–6370. [Google Scholar] [CrossRef] [Green Version]
- Girishkumar, M.S.; Thangaprakash, V.P.; Udaya Bhaskar, T.V.S.; Suprit, K.; Sureshkumar, N.; Baliarsingh, S.K.; Shivaprasad, S. Quantifying tropical cyclone’s effect on the biogeochemical processes using profiling float observations in the Bay of Bengal. J. Geophys. Res. Ocean. 2019, 124, 1945–1963. [Google Scholar] [CrossRef]
- Subrahmanyam, B.; Rao, K.H.; Srinivasa Rao, N.; Murty, V.S.N.; Sharp, R.J. Influence of a tropical cyclone on chlorophyll-a concentration in the Arabian Sea. Geophys. Res. Lett. 2002, 29, 2065. [Google Scholar] [CrossRef]
- Pan, J.; Huang, L.; Devlin, A.T.; Lin, H. Quantification of typhoon-induced phytoplankton blooms using satellite multi-sensor data. Remote Sens. 2018, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.J.; Sui, Y.; Tang, D.L.; Afanasyev, Y.D. A subsurface chlorophyll a bloom induced by typhoon in the South China Sea. J. Mar. Syst. 2013, 128, 138–145. [Google Scholar] [CrossRef]
- Pan, S.; Shi, J.; Gao, H.; Guo, X.; Yao, X.; Gong, X. Contributions of physical and biogeochemical processes to phytoplankton biomass enhancement in the surface and subsurface layers during the passage of typhoon Damrey. J. Geophys. Res. Biogeosci. 2017, 122, 212–229. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, S.; Chen, F.; Ma, Y.; Jiang, C.; Yu, J. Influence of sequential tropical cyclones on phytoplankton blooms in the northwestern South China Sea. J. Oceanol. Limnol. 2021, 39, 14–25. [Google Scholar] [CrossRef]
- Chai, F.; Wang, Y.; Xing, X.; Yan, Y.; Xue, H.; Wells, M.; Boss, E. A limited effect of sub-tropical typhoons on phytoplankton dynamics. Biogeosciences 2021, 18, 849–859. [Google Scholar] [CrossRef]
- Lin, I.; Liu, W.T.; Wu, C.C.; Wong, G.T.; Hu, C.; Chen, Z.; Liu, K.K. New evidence for enhanced ocean primary production triggered by tropical cyclone. Geophys. Res. Lett. 2003, 30, 1718. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yang, Y.J.; Xian, T.; Lu, Z.M.; Fu, Y.F. Strong enhancement of chlorophyll a concentration by a weak typhoon. Mar. Ecol. Prog. Ser. 2010, 404, 39–50. [Google Scholar] [CrossRef]
- Zhang, S.; Xie, L.; Hou, Y.; Zhao, H.; Qi, Y.; Yi, X. Tropical storm-induced turbulent mixing and chlorophyll-a enhancement in the continental shelf southeast of Hainan Island. J. Mar. Syst. 2014, 129, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Jones, B.H.; Brink, K.H.; Fischer, A.S. The upper-ocean response to monsoonal forcing in the Arabian Sea: Seasonal and spatial variability. Deep. Sea Res. Part II Trop. Stud. Oceanogr. 2000, 47, 1177–1226. [Google Scholar] [CrossRef]
- Fischer, A.S.; Weller, R.A.; Rudnick, D.L.; Eriksen, C.C.; Lee, C.M.; Brink, K.H.; Leben, R.R. Mesoscale eddies, coastal upwelling, and the upper-ocean heat budget in the Arabian Sea. Deep. Sea Res. Part II Trop. Stud. Oceanogr. 2002, 49, 2231–2264. [Google Scholar] [CrossRef]
- Evan, A.T.; Camargo, S.J. A climatology of Arabian Sea cyclonic storms. J. Clim. 2011, 24, 140–158. [Google Scholar] [CrossRef] [Green Version]
- Dube, S.K.; Rao, A.D.; Sinha, P.C.; Murty, T.S.; Bahulayan, N. Storm surge in the Bay of Bengal and Arabian Sea the problem and its prediction. Mausam 1997, 48, 283–304. [Google Scholar] [CrossRef]
- Kuttippurath, J.; Sunanda, N.; Martin, M.V.; Chakraborty, K. Tropical storms trigger phytoplankton blooms in the deserts of north Indian Ocean. Clim. Atmos. Sci. 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Murakami, H.; Vecchi, G.A.; Underwood, S. Increasing frequency of extremely severe cyclonic storms over the Arabian Sea. Nat. Clim. Chang. 2017, 7, 885–889. [Google Scholar] [CrossRef]
- Da, N.D.; Foltz, G.R.; Balaguru, K. Observed Global Increases in Tropical Cyclone-Induced Ocean Cooling and Primary Production. Geophys. Res. Lett. 2021, 48, e2021GL092574. [Google Scholar] [CrossRef]
- Maritorena, S.; d’Andon, O.H.F.; Mangin, A.; Siegel, D.A. Merged satellite ocean colordata products using a bio-optical model: Characteristics, benefits and issues. Remote Sens. Environ. 2010, 114, 1791–1804. [Google Scholar] [CrossRef]
- Garver, S.A.; Siegel, D.A. Inherent optical property inversion of ocean color spectra and its biogeochemical interpretation: 1. Time series from the Sargasso Sea. J. Geophys. Res. Ocean. 1997, 102, 18607–18625. [Google Scholar] [CrossRef]
- Aumont, O.; Éthé, C.; Tagliabue, A.; Bopp, L.; Gehlen, M. PISCES-v2: An ocean biogeochemical model for carbon and ecosystem studies. Geosci. Model Dev. 2015, 8, 2465–2513. [Google Scholar] [CrossRef] [Green Version]
- Lellouche, J.M.; Greiner, E.; Galloudec, O.L.; Garric, G.; Regnier, C.; Drevillon, M.; Traon, P.Y.L. Recent updates to the Copernicus Marine Service global ocean monitoring and forecasting real-time 1/12° high-resolution system. Ocean. Sci. 2018, 14, 1093–1126. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.D.; Vickery, P.J.; Reinhold, T.A. Reduced drag coefficient for high wind speeds in tropical cyclones. Nature 2003, 422, 279–283. [Google Scholar] [CrossRef] [PubMed]
- de Boyer Montégut, C.; Madec, G.; Fischer, A.S.; Lazar, A.; Iudicone, D. Mixed layer depth over the global ocean: An examination of profile data and a profile-based climatology. J. Geophys. Res. Ocean. 2004, 109, C12003. [Google Scholar] [CrossRef]
- Rochford, P.A.; Kara, A.B.; Wallcraft, A.J.; Arnone, R.A. Importance of solar subsurface heating in ocean general circulation models. J. Geophys. Res. Ocean. 2001, 106, 30923–30938. [Google Scholar] [CrossRef]
- Mei, W.; Pasquero, C.; Primeau, F. The effect of translation speed upon the intensity of tropical cyclones over the tropical ocean. Geophys. Res. Lett. 2012, 39, L07801. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Li, C. Upper ocean response to the passage of two sequential typhoons. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2018, 132, 68–79. [Google Scholar] [CrossRef]
- Nelson, D.M.; Smith, W., Jr. Sverdrup revisited: Critical depths, maximum chlorophyll levels, and the control of Southern Ocean productivity by the irradiance-mixing regime. Limnol. Oceanogr. 1991, 36, 1650–1661. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Y. Phytoplankton increases induced by tropical cyclones in the South China Sea during 1998–2015. J. Geophys. Res. Ocean. 2018, 123, 2903–2920. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, D.; Zhou, L.; Liu, X.; Ding, T.; Zhou, B. Upper Ocean response to typhoon Kalmaegi (2014). J. Geophys. Res. Ocean. 2016, 121, 6520–6535. [Google Scholar] [CrossRef]
- Busireddy, N.K.R.; Ankur, K.; Osuri, K.K.; Sivareddy, S.; Niyogi, D. The response of ocean parameters to tropical cyclones in the Bay of Bengal. Q. J. R. Meteorol. Soc. 2019, 145, 3320–3332. [Google Scholar] [CrossRef] [Green Version]
- Shay, L.K.; Goni, G.J.; Black, P.G. Effects of a warm oceanic feature on Hurricane Opal. Mon. Weather. Rev. 2000, 128, 1366–1383. [Google Scholar] [CrossRef]
- Liu, X.; Wang, M.; Shi, W. A study of a Hurricane Katrina–induced phytoplankton bloom using satellite observations and model simulations. J. Geophys. Res. Ocean. 2008, 114. [Google Scholar] [CrossRef] [Green Version]
- Mei, W.; Pasquero, C. Spatial and temporal characterization of sea surface temperature response to tropical cyclones. J. Clim. 2013, 26, 3745–3765. [Google Scholar] [CrossRef]
- Stramma, L.; Cornillon, P.; Price, J.F. Satellite observations of sea surface cooling by hurricanes. J. Geophys. Res. Ocean. 1986, 91, 5031–5035. [Google Scholar] [CrossRef]
- Geisler, J.E. Linear theory of the response of a two-layer ocean to a moving hurricane. Geophys. Astrophys. Fluid Dyn. 1970, 1, 249–272. [Google Scholar] [CrossRef]
- Chang, Y.C.; Tseng, R.S.; Chu, P.C.; Chen, J.M.; Centurioni, L.R. Observed strong currents under global tropical cyclones. J. Mar. Syst. 2016, 159, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Oey, L.Y. Right-side cooling and phytoplankton bloom in the wake of a tropical cyclone. J. Geophys. Res. Ocean. 2015, 120, 5735–5748. [Google Scholar] [CrossRef]
- Niu, L.; van Gelder, P.H.A.J.M.; Zhang, C.; Guan, Y.; Vrijling, J.K. Physical control of phytoplankton bloom development in the coastal waters of Jiangsu (China). Ecol. Model. 2016, 321, 75–83. [Google Scholar] [CrossRef]
- Large, W.G.; McWilliams, J.C.; Doney, S.C. Oceanic vertical mixing: A review and a model with a nonlocal boundary layer parameterization. Rev. Geophys. 1994, 32, 363–403. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, B.; Shay, L.K. Mixed layer cooling in mesoscale oceanic eddies during Hurricanes Katrina and Rita. Mon. Weather. Rev. 2009, 137, 4188–4207. [Google Scholar] [CrossRef] [Green Version]
- Chelton, D.B.; Gaube, P.; Schlax, M.G.; Early, J.J.; Samelson, R.M. The influence of nonlinear mesoscale eddies on near-surfaceoceanic chlorophyll. Science 2011, 334, 328–332. [Google Scholar] [CrossRef]
- Chang, Y.L.; Miyazawa, Y.; Oey, L.Y.; Kodaira, T.; Huang, S. The formation processes of phytoplankton growth and decline in mesoscale eddies in the western North Pacific Ocean. J. Geophys. Res. Ocean. 2017, 122, 4444–4455. [Google Scholar] [CrossRef]
- Wang, T.; Du, Y.; Liao, X.; Xiang, C. Evidence of Eddy-Enhanced Winter Chlorophyll-a Blooms in Northern Arabian Sea: 2017 Cruise Expedition. J. Geophys. Res. Ocean. 2020, 125, e2019JC015582. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, D.; Evgeny, M. Chlorophyll concentration response to the typhoon wind-pump induced upper ocean processes considering air–sea heat exchange. Remote Sens. 2019, 11, 1825. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Sharples, J.; Mahaffey, C.; Rippeth, T. Wind-driven nutrient pulses to the subsurface chlorophyll maximum in seasonally stratified shelf seas. Geophys. Res. Lett. 2013, 40, 5467–5472. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Matsuno, T.; Endoh, T.; Ishizaka, J.; Zhu, Y.; Takeda, S.; Sukigara, C. A role of vertical mixing on nutrient supply into the subsurface chlorophyll maximum in the shelf region of the East China Sea. Cont. Shelf Res. 2016, 143, 139–150. [Google Scholar] [CrossRef]
- Du, C.; Liu, Z.; Kao, S.J.; Dai, M. Diapycnal fluxes of nutrients in an oligotrophic oceanic regime: The South China Sea. Geophys. Res. Lett. 2017, 44, 11–510. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Boss, E.S. Resurrecting the Ecological Underpinnings of Ocean Plankton Blooms. Annu. Rev. Mar. Sci. 2014, 6, 167–208. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, B.; Shay, L.K.; Halliwell, G.R. The response of quasigeostrophic oceanic vortices to tropical cyclone forcing. J. Phys. Oceanogr. 2011, 41, 1965–1985. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | e1 | e2 | ||||
|---|---|---|---|---|---|---|
| Argo | 2902272 | 2902210 | ||||
| Period | Pre-TC (22 October) | Post Kyarr (1 November) | Post Maha (11 November) | Pre-TC (25 October) | During Maha (4 November) | Post Maha (14 November) |
| Surface chl-a (mg/m3) | 0.67 | 1.7 | 1.12 | 0.3 | 0.45 | 4.97 |
| Max chl-a (mg/m3)/depth (m) | 1.68/29 | 2.1/20 | 7.12/24 | 1.26/47 | 0.50/26 | 5/9 |
| Integrate chl-a (mg/m2) | 51.3 | 60 | 131.01 | 47.89 | 39.64 | 117 |
| MLD (m)/variance (m) | 24.12/5.2 | 44.51/7.6 | 25.69/4.5 | 31.78/6.2 | 58.53/6.9 | 30.67/5.4 |
| EPV (m/s) (period) | 2.3 × 10−5 (Pre-TC) | 2.8 × 10−4 (During Kyarr) | 1.0 × 10−4 (During Maha) | 1.8 × 10−5 (Pre-TC) | 9.1 × 10−5 (During Kyarr) | 2.7 × 10−4 (During Maha) |
| Total wind work (KJ) | 72 | 85 | ||||
| Total forcing time (hour) | 36 | 57 | ||||
| Eddy | Radius (km) | Maximum Geostrophic Current (m/s) | Maximum SST Cooling (°C) | Minimum SLA (cm) | Maximum Chl-a (mg/m3) | Maximum EPV (m/s) |
|---|---|---|---|---|---|---|
| e1 | 166.5 | 0.59 | −3.95 | −7.26 | 12.76 | 8.1 × 10−4 |
| e2 | 175.2 | 0.56 | −5.28 | −14.27 | 23.09 | 9.3 × 10−4 |
| e3 | 186.4 | 0.75 | −5.47 | −20.56 | 16.51 | 6.9 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Chen, F.; Zhang, S.; Pan, J.; Devlin, A.T.; Ning, H.; Zeng, W. Physical and Biochemical Responses to Sequential Tropical Cyclones in the Arabian Sea. Remote Sens. 2022, 14, 529. https://doi.org/10.3390/rs14030529
Wang T, Chen F, Zhang S, Pan J, Devlin AT, Ning H, Zeng W. Physical and Biochemical Responses to Sequential Tropical Cyclones in the Arabian Sea. Remote Sensing. 2022; 14(3):529. https://doi.org/10.3390/rs14030529
Chicago/Turabian StyleWang, Tongyu, Fajin Chen, Shuwen Zhang, Jiayi Pan, Adam T. Devlin, Hao Ning, and Weiqiang Zeng. 2022. "Physical and Biochemical Responses to Sequential Tropical Cyclones in the Arabian Sea" Remote Sensing 14, no. 3: 529. https://doi.org/10.3390/rs14030529
APA StyleWang, T., Chen, F., Zhang, S., Pan, J., Devlin, A. T., Ning, H., & Zeng, W. (2022). Physical and Biochemical Responses to Sequential Tropical Cyclones in the Arabian Sea. Remote Sensing, 14(3), 529. https://doi.org/10.3390/rs14030529