
Mapping the Abundance of Multipurpose Agroforestry Faidherbia albida Trees in Senegal

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
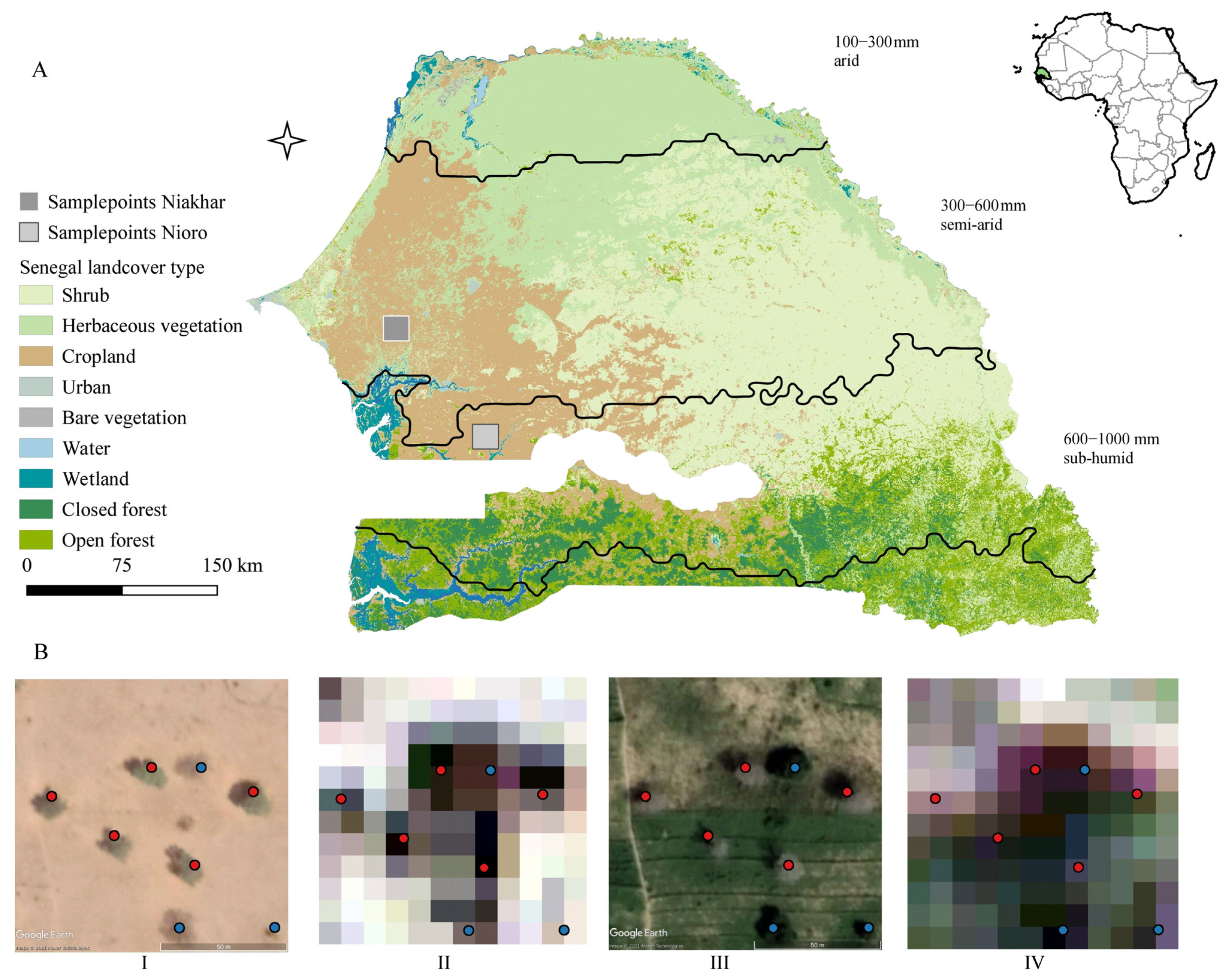
2.1. Study Area and Sample Data
2.2. Sentinel-2 Data Pre-Processing
2.2.1. Vegetation Indices
2.2.2. Feature Selection
2.3. Mapping Faidherbia albida Trees Using Artificial Neural Networks
2.4. Post Processing
3. Results
3.1. Intra-Annual Difference in Vegetation Indices
3.2. Mapping Faidherbia albida Using the MLP Model
3.3. Comparing Faidherbia albida Canopy Cover Maps with Potential Occurrence Maps
4. Discussion
4.1. Mapping Faidherbia albida
4.2. Comparison with Ecological Niche Modelling Products
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez, P.; Tucker, C.J.; Sy, H. Tree density and species decline in the African Sahel attributable to climate. J. Arid. Environ. 2012, 78, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Karlson, M.; Reese, H.; Ostwald, M. Tree crown mapping in managed woodlands (parklands) of semi-arid West Africa using WorldView-2 imagery and geographic object based image analysis. Sensors 2014, 14, 22643–22669. [Google Scholar] [CrossRef] [PubMed]
- Sinare, H.; Gordon, L.J. Ecosystem services from woody vegetation on agricultural lands in Sudano-Sahelian West Africa. Agr. Ecosyst. Environ. 2015, 200, 186–199. [Google Scholar] [CrossRef]
- Kuyah, S.; Öborn, I.; Jonsson, M.; Dahlin, A.S.; Barrios, E.; Muthuri, C.; Malmer, A.; Nyaga, J.; Magaju, C.; Namirembe, S. Trees in agricultural landscapes enhance provision of ecosystem services in Sub-Saharan Africa. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2016, 12, 255–273. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.; Rasmussen, K.; Hiernaux, P.; Herrmann, S.; Tucker, C.J.; Tong, X.; Tian, F.; Mertz, O.; Kergoat, L.; Mbow, C. Reduction of tree cover in West African woodlands and promotion in semi-arid farmlands. Nat. Geosci. 2018, 11, 328–333. [Google Scholar] [CrossRef]
- Koffi, C.; Lourme-Ruiz, A.; Djoudi, H.; Bouquet, E.; Dury, S.; Gautier, D. The contributions of wild tree resources to food and nutrition security in sub-Saharan African drylands: A review of the pathways and beneficiaries. Int. Forest. Rev. 2020, 22, 64–82. [Google Scholar] [CrossRef]
- Tschakert, P.; Khouma, M.; Sene, M. Biophysical potential for soil carbon sequestration in agricultural systems of the Old Peanut Basin of Senegal. J. Arid. Environ. 2004, 59, 511–533. [Google Scholar] [CrossRef]
- Hiernaux, P.H.; Cissé, M.; Diarra, L.; De Leeuw, P. Fluctuations saisonnières de la feuillaison des arbres et des buissons sahéliens. Conséquences pour la quantification des ressources fourragères. Rev. d’Elevage. Méd. Vét. Pays. Trop. 1994, 47, 117–125. [Google Scholar] [CrossRef]
- Garrity, D.P.; Akinnifesi, F.K.; Ajayi, O.C.; Weldesemayat, S.G.; Mowo, J.G.; Kalinganire, A.; Larwanou, M.; Bayala, J. Evergreen Agriculture: A robust approach to sustainable food security in Africa. Food Secur. 2010, 2, 197–214. [Google Scholar] [CrossRef]
- Wood, P.J. Faidherbia albida (Del.) A Chev. (Synonym: Acacia albida Del.): A Monograph; CTFT: Nogent-sur-Marne, France, 1989. [Google Scholar]
- Boffa, J.M. Agroforestry Parklands in Sub-Saharan Africa; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999. [Google Scholar]
- Barnes, R.; Fagg, C.W. Faidherbia albida Monograph and Annotated Bibliography. 2003. Available online: https://ora.ox.ac.uk/objects/uuid:fe18e8c9-1a92-435f-94c2-7c5827cbea57 (accessed on 23 November 2021).
- Glover, J.D.; Reganold, J.P.; Cox, C.M. Plant perennials to save Africa’s soils. Nature 2012, 489, 359–361. [Google Scholar] [CrossRef]
- Noulekoun, F.; Birhane, E.; Chude, S.; Zenebe, A. Characterization of Faidherbia albida (Del.) A. Chev. population in agroforestry parklands in the highlands of Northern Ethiopia: Impact of conservation, environmental factors and human disturbances. Agroforest. Syst. 2017, 91, 123–135. [Google Scholar] [CrossRef]
- Vandenbeldt, R.; Williams, J. The effect of soil surface temperature on the growth of millet in relation to the effect of Faidherbia albida trees. Agr. Forest Meteorol. 1992, 60, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Saka, A.; Bunderson, W.; Itimu, O.; Phombeya, H.; Mbekeani, Y. The effects of Acacia albida on soils and maize grain yields under smallholder farm conditions in Malawi. Forest. Ecol. Manag. 1994, 64, 217–230. [Google Scholar] [CrossRef]
- Hadgu, K.M.; Kooistra, L.; Rossing, W.A.; van Bruggen, A.H. Assessing the effect of Faidherbia albida based land use systems on barley yield at field and regional scale in the highlands of Tigray, Northern Ethiopia. Food Secur. 2009, 1, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Sida, T.S.; Baudron, F.; Kim, H.; Giller, K.E. Climate-smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. Forest Meteorol. 2018, 248, 339–347. [Google Scholar] [CrossRef]
- Leroux, L.; Falconnier, G.N.; Diouf, A.A.; Ndao, B.; Gbodjo, J.; Tall, L.; Balde, A.; Clermont-Dauphin, C.; Bégué, A.; Affholder, F. Using remote sensing to assess the effect of trees on millet yield in complex parklands of Central Senegal. Agric. Syst. 2020, 184, 102918. [Google Scholar] [CrossRef]
- Roupsard, O.; Audebert, A.; Ndour, A.P.; Clermont-Dauphin, C.; Agbohessou, Y.; Sanou, J.; Koala, J.; Faye, E.; Sambakhe, D.; Jourdan, C. How far does the tree affect the crop in agroforestry? New spatial analysis methods in a Faidherbia parkland. Agric. Ecosyst. Environ. 2020, 296, 106928. [Google Scholar] [CrossRef]
- Rinaudo, T. The development of farmer managed natural regeneration. LEISA Mag. 2007, 23, 32–34. [Google Scholar]
- Haglund, E.; Ndjeunga, J.; Snook, L.; Pasternak, D. Dry land tree management for improved household livelihoods: Farmer managed natural regeneration in Niger. J. Environ. Manag. 2011, 92, 1696–1705. [Google Scholar] [CrossRef] [Green Version]
- Weston, P.; Hong, R.; Kaboré, C.; Kull, C.A. Farmer-managed natural regeneration enhances rural livelihoods in dryland West Africa. Environ. Manag. 2015, 55, 1402–1417. [Google Scholar] [CrossRef] [Green Version]
- Chomba, S.; Sinclair, F.; Savadogo, P.; Bourne, M.; Lohbeck, M. Opportunities and constraints for using farmer managed natural regeneration for land restoration in sub-Saharan Africa. Front. For. Glob. Chang. 2020, 3, 122. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Chakraborty, A.; Joshi, P.; Sachdeva, K. Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region. Ecol. Eng. 2016, 97, 593–609. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Karlson, M.; Ostwald, M.; Reese, H.; Bazié, H.R.; Tankoano, B. Assessing the potential of multi-seasonal WorldView-2 imagery for mapping West African agroforestry tree species. Int. J. Appl. Earth Observ. Geoinform. 2016, 50, 80–88. [Google Scholar] [CrossRef]
- Wang, L.; Sousa, W.; Gong, P. Integration of object-based and pixel-based classification for mapping mangroves with IKONOS imagery. Int. J. Remote Sens. 2004, 25, 5655–5668. [Google Scholar] [CrossRef]
- Tigges, J.; Lakes, T.; Hostert, P. Urban vegetation classification: Benefits of multitemporal RapidEye satellite data. Remote Sens. Environ. 2013, 136, 66–75. [Google Scholar] [CrossRef]
- Adelabu, S.; Dube, T. Employing ground and satellite-based QuickBird data and random forest to discriminate five tree species in a Southern African Woodland. Geocarto. Int. 2015, 30, 457–471. [Google Scholar] [CrossRef]
- Cho, M.A.; Malahlela, O.; Ramoelo, A. Assessing the utility WorldView-2 imagery for tree species mapping in South African subtropical humid forest and the conservation implications: Dukuduku forest patch as case study. Int. J. Appl. Earth Observ. Geoinform. 2015, 38, 349–357. [Google Scholar] [CrossRef]
- Korpela, I.; Ørka, H.O.; Maltamo, M.; Tokola, T.; Hyyppä, J. Tree species classification using airborne LiDAR–effects of stand and tree parameters, downsizing of training set, intensity normalization, and sensor type. Silva. Fenn. 2010, 44, 319–339. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Krzystek, P.; Heurich, M. Tree species classification and estimation of stem volume and DBH based on single tree extraction by exploiting airborne full-waveform LiDAR data. Remote Sens. Environ. 2012, 123, 368–380. [Google Scholar] [CrossRef]
- Heinzel, J.; Koch, B. Exploring full-waveform LiDAR parameters for tree species classification. Int. J. Appl. Earth Observ. Geoinform. 2011, 13, 152–160. [Google Scholar] [CrossRef]
- Cho, M.A.; Mathieu, R.; Asner, G.P.; Naidoo, L.; Van Aardt, J.; Ramoelo, A.; Debba, P.; Wessels, K.; Main, R.; Smit, I.P. Mapping tree species composition in South African savannas using an integrated airborne spectral and LiDAR system. Remote Sens. Environ. 2012, 125, 214–226. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Tree species classification in the Southern Alps based on the fusion of very high geometrical resolution multispectral/hyperspectral images and LiDAR data. Remote Sens. Environ. 2012, 123, 258–270. [Google Scholar] [CrossRef]
- Kukkonen, M.; Maltamo, M.; Korhonen, L.; Packalen, P. Multispectral Airborne Lidar Data in the Prediction of Boreal Tree Species Composition. IEEE Trans. Geosci. Remote Sens. 2019, 57, 3462–3471. [Google Scholar] [CrossRef]
- Gärtner, P.; Förster, M.; Kleinschmit, B. The Benefit of Synthetically Generated Rapideye and Landsat 8 Data Fusion Time Series for Riparian Forest Disturbance Monitoring. Remote Sens. Environ. 2016, 177, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Schriever, J.R.; Congalton, R.G. Evaluating Seasonal Variability as an Aid to Cover-Type Mapping from Landsat Thematic Mapper Data in the Northeast. Photogramm. Eng. Remote Sens. 1995, 61, 321–327. [Google Scholar]
- Hesketh, M.; Sánchez-Azofeifa, G.A. The Effect of Seasonal Spectral Variation on Species Classification in the Panamanian Tropical Forest. Remote Sens. Environ. 2012, 118, 73–82. [Google Scholar] [CrossRef]
- Key, T.; Warner, T.A.; McGraw, J.B.; Fajvan, M.A. A Comparison of Multispectral and Multitemporal Information in High Spatial Resolution Imagery for Classification of Individual Tree Species in a Temperate Hardwood Forest. Remote Sens. Environ. 2001, 75, 100–112. [Google Scholar] [CrossRef]
- Sheeren, D.; Fauvel, M.; Josipović, V.; Lopes, M.; Planque, C.; Willm, J.; Dejoux, J.-F. Tree Species Classification in Temperate Forests Using Formosat-2 Satellite Image Time Series. Remote Sens. 2016, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Mickelson, J.G.; Civco, D.L.; Silander, J.A. Delineating Forest Canopy Species in the Northeastern United States Using Multi-Temporal Tm Imagery. Photogramm. Eng. Remote Sens. 1998, 64, 891–904. [Google Scholar]
- Triboulet, C. Identification des parcs à Faidherbia albida par télédétection. Cirad-Forêt Cahiers Sci. 1996, 12, 203–216. [Google Scholar]
- Lelong, C.C.; Tshingomba, U.K.; Soti, V. Assessing Worldview-3 multispectral imaging abilities to map the tree diversity in semi-arid parklands. Int. J. Appl. Earth Observ. Geoinform. 2020, 93, 102211. [Google Scholar] [CrossRef]
- Zhang, W.; Brandt, M.; Wang, Q.; Prishchepov, A.V.; Tucker, C.J.; Li, Y.; Lyu, H.; Fensholt, R. From woody cover to woody canopies: How Sentinel-1 and Sentinel-2 data advance the mapping of woody plants in savannas. Remote Sens. Environ. 2019, 234, 111465. [Google Scholar] [CrossRef]
- Chan, J.C.-W.; Paelinckx, D. Evaluation of Random Forest and Adaboost tree-based ensemble classification and spectral band selection for ecotope mapping using airborne hyperspectral imagery. Remote Sens. Environ. 2008, 112, 2999–3011. [Google Scholar] [CrossRef]
- Dalponte, M.; Ørka, H.O.; Gobakken, T.; Gianelle, D.; Næsset, E. Tree species classification in boreal forests with hyperspectral data. IEEE Transac. Geosci. Remote Sens. 2012, 51, 2632–2645. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree species classification with random forest using very high spatial resolution 8-band WorldView-2 satellite data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef] [Green Version]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P. Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Tottrup, C.; Rasmussen, M.S. Mapping long-term changes in savannah crop productivity in Senegal through trend analysis of time series of remote sensing data. Agr. Ecosyst. Environ. 2004, 103, 545–560. [Google Scholar] [CrossRef]
- Lufafa, A.; Diédhiou, I.; Samba, S.; Séné, M.; Khouma, M.; Kizito, F.; Dick, R.; Dossa, E.; Noller, J. Carbon stocks and patterns in native shrub communities of Senegal’s Peanut Basin. Geoderma 2008, 146, 75–82. [Google Scholar] [CrossRef]
- Lamprecht, H. Silviculture in the Tropics; GTZ: Eschborn, Germany, 1989; p. 296. [Google Scholar]
- Sileshi, G.W. The magnitude and spatial extent of influence of Faidherbia albida trees on soil properties and primary productivity in drylands. J. Arid. Environ. 2016, 132, 1–14. [Google Scholar] [CrossRef]
- Tschakert, P. Carbon for farmers: Assessing the potential for soil carbon sequestration in the old peanut basin of Senegal. Clim. Chang. 2004, 67, 273–290. [Google Scholar] [CrossRef]
- Ndao, B.; Leroux, L.; Gaetano, R.; Diouf, A.A.; Soti, V.; Bégué, A.; Mbow, C.; Sambou, B. Landscape heterogeneity analysis using geospatial techniques and a priori knowledge in Sahelian agroforestry systems of Senegal. Ecol. Indic. 2021, 125, 107481. [Google Scholar] [CrossRef]
- Ndao, B.; Leroux, L.; Hema, A.; Diouf, A.A.; Bégué, A.; Sambou, B. Tree Species Diversity Mapping Using Species Distribution Models: A Faidherbia albida Parkland Case Study in Senegal. Ecol. Model 2021. submitted. [Google Scholar]
- Brandt, M.; Tucker, C.J.; Kariryaa, A.; Rasmussen, K.; Abel, C.; Small, J.; Chave, J.; Rasmussen, L.V.; Hiernaux, P.; Diouf, A.A. An unexpectedly large count of trees in the West African Sahara and Sahel. Nature 2020, 587, 78–82. [Google Scholar] [CrossRef]
- Frampton, W.J.; Dash, J.; Watmough, G.; Milton, E.J. Evaluating the capabilities of Sentinel-2 for quantitative estimation of biophysical variables in vegetation. ISPRS J. Photogramm. Remote Sens. 2013, 82, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Hripcsak, G.; Rothschild, A.S. Agreement, the f-measure, and reliability in information retrieval. J. Am. Med. Inform. Assoc. 2005, 12, 296–298. [Google Scholar] [CrossRef]
- Kindt, R. Ensemble species distribution modelling with transformed suitability values. Environ. Model Softw. 2018, 100, 136–145. [Google Scholar] [CrossRef]
- Yan, E.; Wang, G.; Lin, H.; Xia, C.; Sun, H. Phenology-Based Classification of Vegetation Cover Types in Northeast China Using Modis Ndvi and Evi Time Series. Int. J. Remote Sens. 2015, 36, 489–512. [Google Scholar] [CrossRef]
- Bradley, B.A. Remote detection of invasive plants: A review of spectral, textural and phenological approaches. Biol. Invas. 2014, 16, 1411–1425. [Google Scholar] [CrossRef]
- Franklin, S.E.; Hall, R.J.; Moskal, L.M.; Maudie, A.J.; Lavigne, M.B. Incorporating Texture into Classification of Forest Species Composition from Airborne Multispectral Images. Int. J. Remote Sens. 2000, 21, 61–79. [Google Scholar] [CrossRef]
- Lykke, A.M. Local perceptions of vegetation change and priorities for conservation of woody-savanna vegetation in Senegal. J. Environ. Manag. 2000, 59, 107–120. [Google Scholar] [CrossRef]
- Lykke, A.; Kristensen, M.; Ganaba, S. Valuation of local use and dynamics of 56 woody species in the Sahel. Biodiv. Conserv. 2004, 13, 1961–1990. [Google Scholar] [CrossRef]
- Sambou, A.; Sambou, B.; Ræbild, A. Farmers’ contributions to the conservation of tree diversity in the Groundnut Basin, Senegal. J. Forest. Res. 2017, 28, 1083–1096. [Google Scholar] [CrossRef]
- Kindt, R. A climate change atlas for Africa. In Presentation Made during a Workshop on the Estimation of the Potential of Agroforestry to Mitigate Climate Change in Sub-Saharan Africa; CIRAD: Montpellier, France, 2021. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monthly Feature | Seasonal Dynamic Feature | ||||
|---|---|---|---|---|---|
| Band/Index | Month | Feature Importance | Band/Index | Month | Feature Importance |
| NDI54 | 2 | 0.040 | NDVI | 2–6 | 0.063 |
| Band12 | 6 | 0.035 | NDI54 | 2–6 | 0.043 |
| Band12 | 3 | 0.032 | NDVI | 2–5 | 0.041 |
| Band11 | 6 | 0.031 | NDI54 | 8–12 | 0.038 |
| Band12 | 5 | 0.030 | NDVI | 4–6 | 0.031 |
| NDI54 | 12 | 0.029 | NDVI | 7–12 | 0.031 |
| NDI54 | 3 | 0.029 | NDI54 | 2–5 | 0.029 |
| Band2 | 10 | 0.026 | NDI54 | 2–8 | 0.024 |
| Band12 | 4 | 0.025 | Band12 | 3–11 | 0.023 |
| NDI54 | 4 | 0.025 | NDVI | 2–7 | 0.023 |
| NDI54 | 6 | 0.025 | NDVI | 6–12 | 0.022 |
| NDVI | 6 | 0.023 | NDI54 | 10–11 | 0.020 |
| Band3 | 10 | 0.022 | NDI54 | 3–8 | 0.020 |
| Band8 | 10 | 0.019 | NDI54 | 2–10 | 0.018 |
| NDVI | 10 | 0.017 | Band12 | 3–10 | 0.018 |
| NDVI | 7 | 0.017 | Band12 | 4–11 | 0.017 |
| Band3 | 10–12 | 0.017 | |||
| NDVI | 4–7 | 0.015 | |||
| Band4 | 10–11 | 0.014 | |||
| NDI54 | 10–12 | 0.014 | |||
| NDI54 | 1–8 | 0.014 | |||
| Band4 | 6–12 | 0.014 | |||
| NDVI | 3–7 | 0.010 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.; Brandt, M.; Tong, X.; Hiernaux, P.; Leroux, L.; Ndao, B.; Fensholt, R. Mapping the Abundance of Multipurpose Agroforestry Faidherbia albida Trees in Senegal. Remote Sens. 2022, 14, 662. https://doi.org/10.3390/rs14030662
Lu T, Brandt M, Tong X, Hiernaux P, Leroux L, Ndao B, Fensholt R. Mapping the Abundance of Multipurpose Agroforestry Faidherbia albida Trees in Senegal. Remote Sensing. 2022; 14(3):662. https://doi.org/10.3390/rs14030662
Chicago/Turabian StyleLu, Tingting, Martin Brandt, Xiaoye Tong, Pierre Hiernaux, Louise Leroux, Babacar Ndao, and Rasmus Fensholt. 2022. "Mapping the Abundance of Multipurpose Agroforestry Faidherbia albida Trees in Senegal" Remote Sensing 14, no. 3: 662. https://doi.org/10.3390/rs14030662
APA StyleLu, T., Brandt, M., Tong, X., Hiernaux, P., Leroux, L., Ndao, B., & Fensholt, R. (2022). Mapping the Abundance of Multipurpose Agroforestry Faidherbia albida Trees in Senegal. Remote Sensing, 14(3), 662. https://doi.org/10.3390/rs14030662