
Assessing the Impact of Soil on Species Diversity Estimation Based on UAV Imaging Spectroscopy in a Natural Alpine Steppe

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
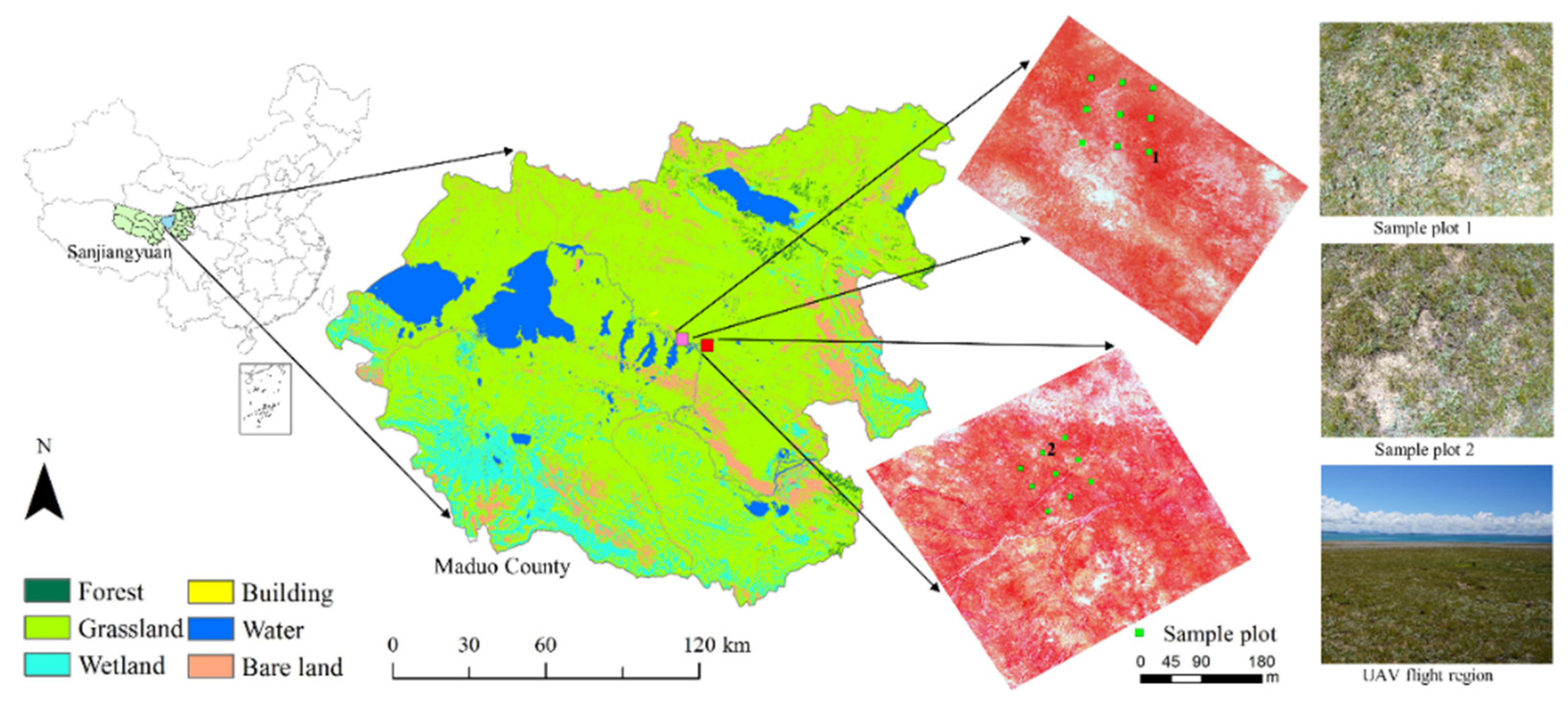
2.1. Study Area
2.2. Imaging Spectroscopy Data and Preprocessing
2.3. Field Measurements
2.4. Species Diversity Indices
2.5. Spectral Diversity Metrics
2.6. Soil Filtering
3. Results
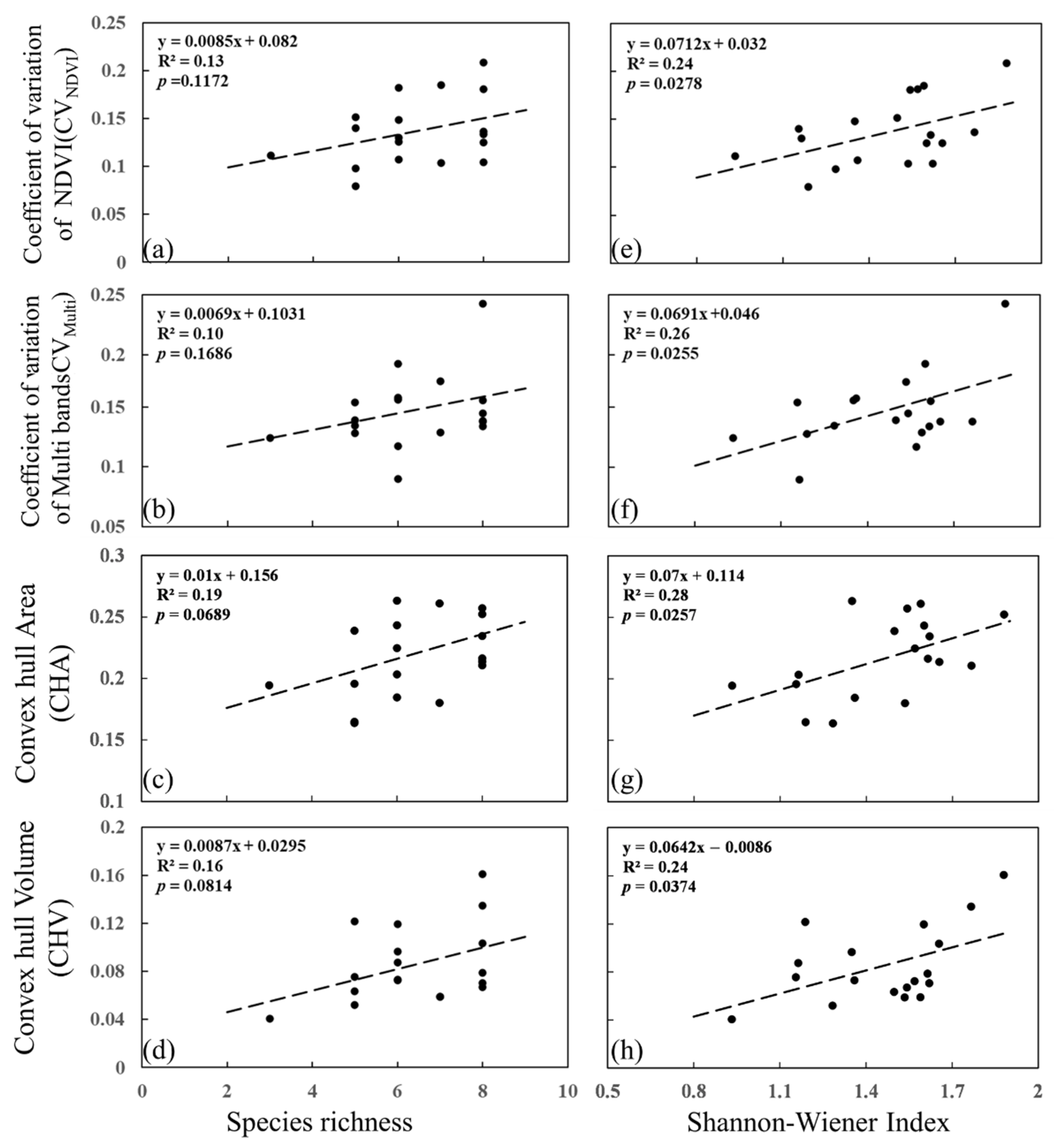
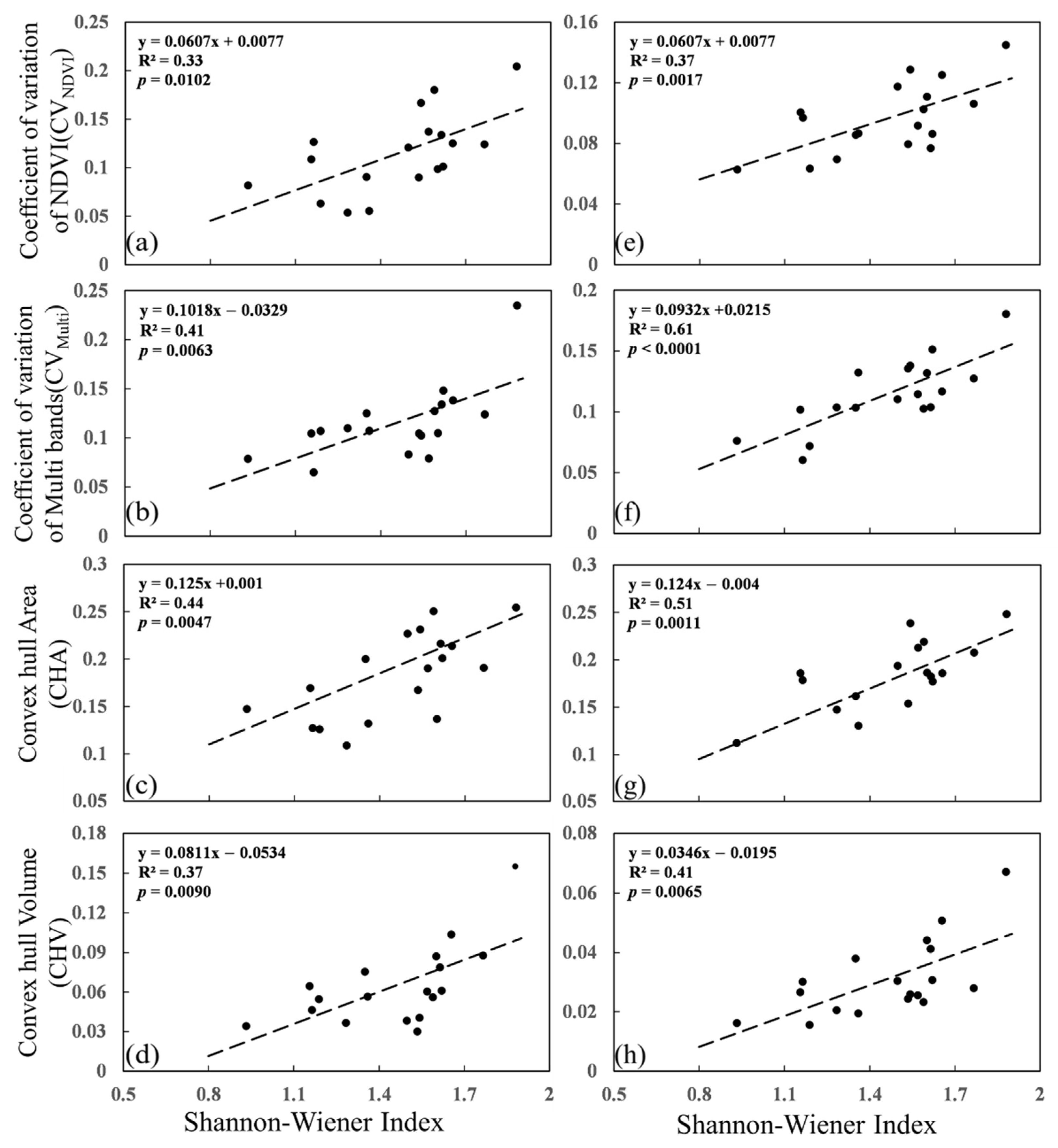
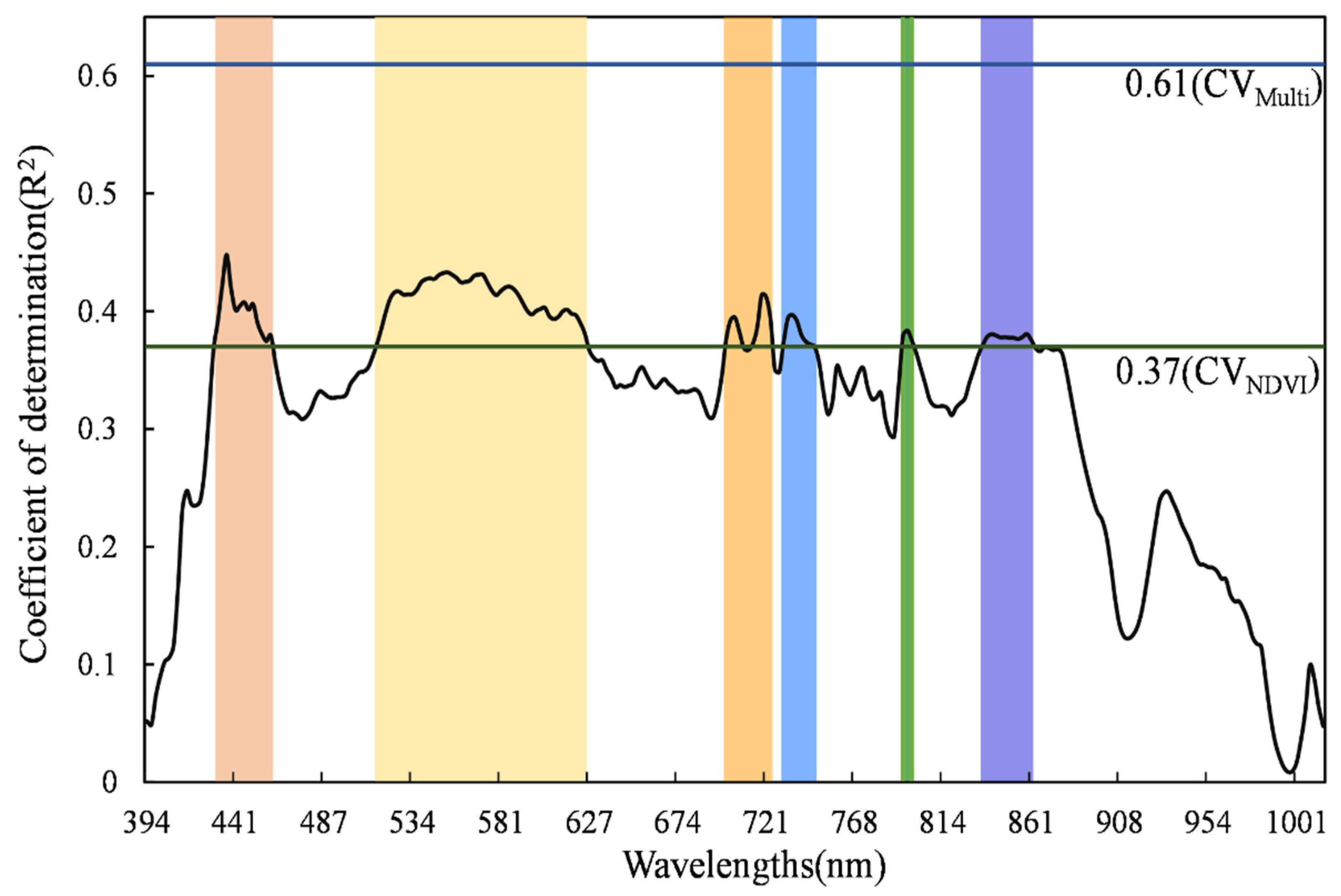
3.1. Responses of Spectral Diversity to Species Diversity
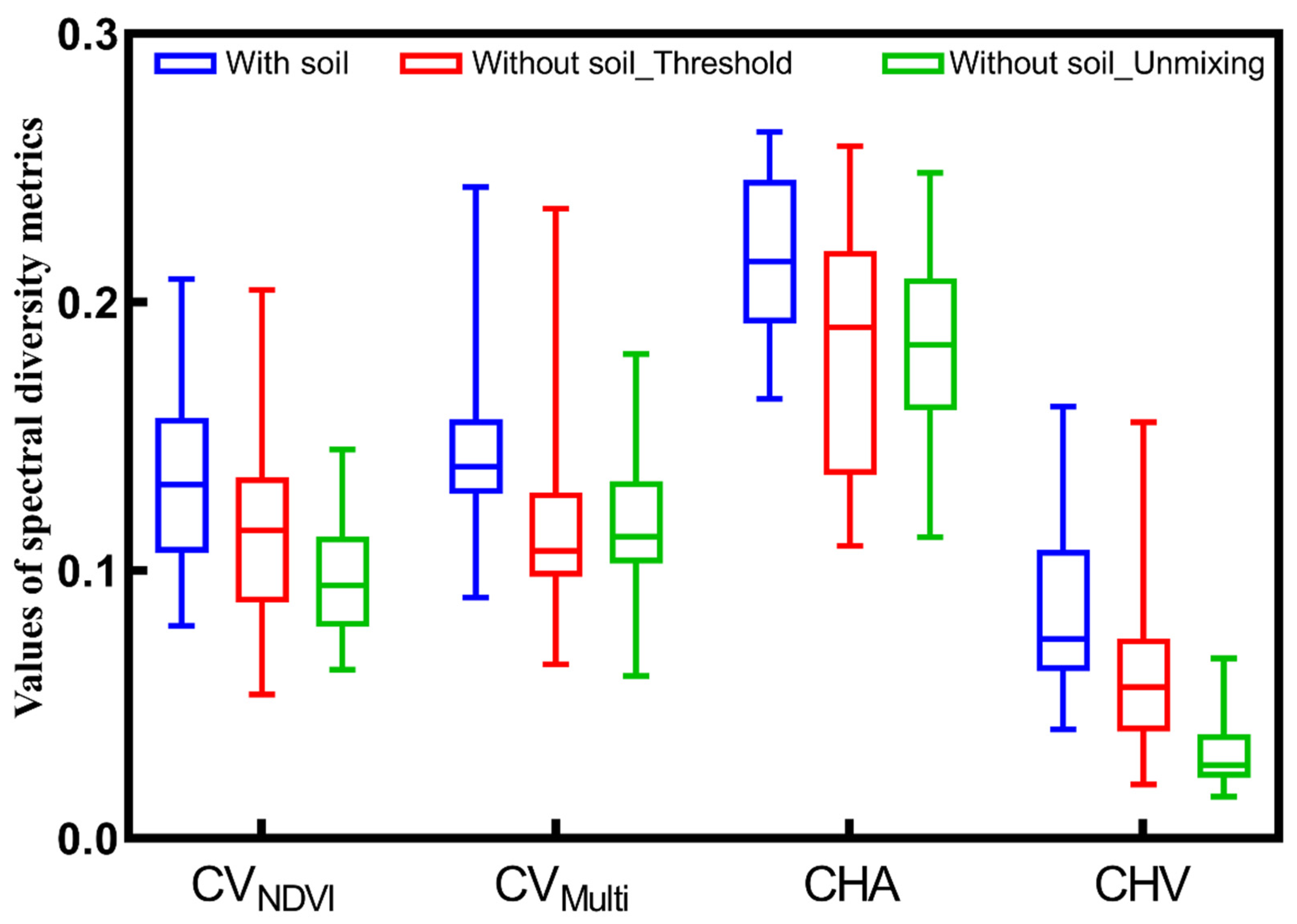
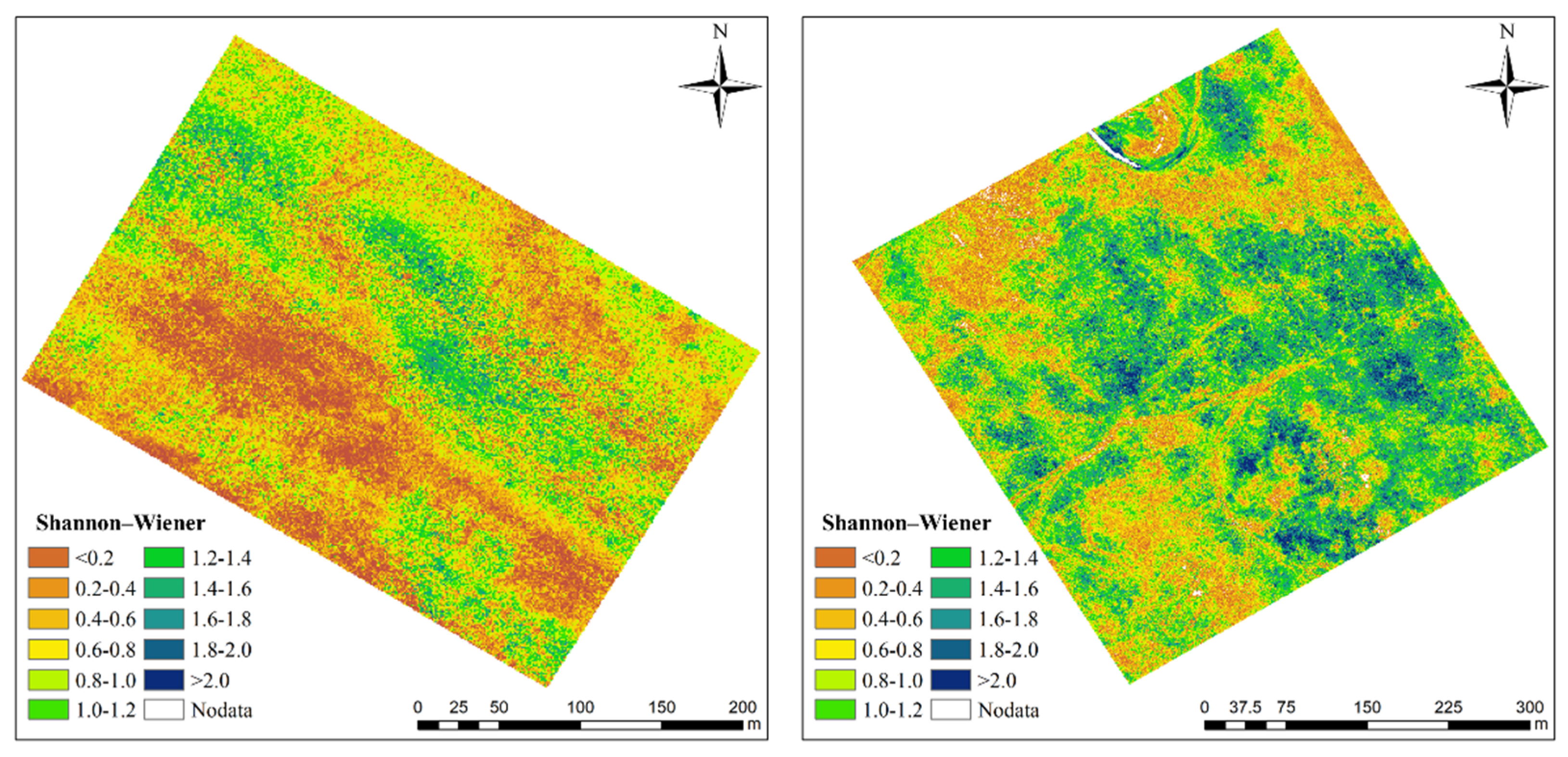
3.2. Impact of Soil on Spectral Diversity Metrics
4. Discussion
4.1. Methods for Grassland Species Diversity Estimation
4.2. Scales for Grassland Diversity Mapping
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Symstad, A.J.; Bai, Y.; Huang, J.; Wu, J.; Naeem, S.; Chen, D.; Tian, D.; Wang, Q.; Han, X. Biodiversity–productivity relationships in a natural grassland community vary under diversity loss scenarios. J. Ecol. 2021, 110, 210–220. [Google Scholar] [CrossRef]
- van Oijen, M.; Bellocchi, G.; Höglind, M. Effects of climate change on grassland biodiversity and productivity: The need for a diversity of models. Agronomy 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Dirzo, R.; Raven, P.H. Global state of biodiversity and loss. Annu. Rev. Environ. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef] [Green Version]
- Linquist, S.; Varner, G.; Newman, J.E. Precis of defending biodiversity. Biol. Philos. 2020, 35, 14. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lu Yonglong, Y.Y.; Sun, B.; Yuan, J.; Yu, M.; Stenseth, N.C.; James, M.; Bullock, M.O. Spatial variation in biodiversity loss across china under multiple environmental stressors. Sci. Adv. 2020, 6, 11. [Google Scholar] [CrossRef]
- Harpole, W.S.; Sullivan, L.L.; Lind, E.M.; Firn, J.; Adler, P.B.; Borer, E.T.; Chase, J.; Fay, P.A.; Hautier, Y.; Hillebrand, H.; et al. Addition of multiple limiting resources reduces grassland diversity. Nature 2016, 537, 93–96. [Google Scholar] [CrossRef]
- Harrison, S.P.; Gornish, E.S.; Copeland, S. Climate-driven diversity loss in a grassland community. Proc. Natl. Acad. Sci. USA 2015, 112, 8672–8677. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Cao, Y.; Yu, D.; Cao, M.; He, Y.; Gill, M.; Pereira, H.M. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 2021, 5, 411–418. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Isbell, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Diaz, S.; Hector, A.; Mace, G.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.E.; et al. Linking the influence and dependence of people on biodiversity across scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lausch, A.; Bannehr, L.; Beckmann, M.; Boehm, C.; Feilhauer, H.; Hacker, J.M.; Heurich, M.; Jung, A.; Klenke, R.; Neumann, C.; et al. Linking earth observation and taxonomic, structural and functional biodiversity: Local to ecosystem perspectives. Ecol. Indic. 2016, 70, 317–339. [Google Scholar] [CrossRef]
- Lyashevska, O.; Farnsworth, K.D. How many dimensions of biodiversity do we need? Ecol. Indic. 2012, 18, 485–492. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Harral, M.J.; Harral, J.E. Scale dependence in plant biodiversity. Science 2001, 291, 5. [Google Scholar] [CrossRef]
- Robert, J.; Whittaker, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 18, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 8. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in diflerent types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Jochum, M.; Fischer, M.; Isbell, F.; Roscher, C.; van der Plas, F.; Boch, S.; Boenisch, G.; Buchmann, N.; Catford, J.A.; Cavender-Bares, J.; et al. The results of biodiversity-ecosystem functioning experiments are realistic. Nat. Ecol. Evol. 2020, 4, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Mi, X.; Feng, G.; Hu, Y.; Zhang, J.; Chen, L.; Corlett, R.T.; Hughes, A.C.; Pimm, S.; Schmid, B.; Shi, S.; et al. The global significance of biodiversity science in china: An overview. Natl. Sci. Rev. 2021, 8, nwab032. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.; Cardoso, A.C.; et al. Ecology. Essential biodiversity variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [Green Version]
- Pettorelli, N.; Laurance, W.F.; O’Brien, T.G.; Wegmann, M.; Nagendra, H.; Turner, W.; Milner-Gulland, E.J. Satellite remote sensing for applied ecologists: Opportunities and challenges. J. Appl. Ecol. 2014, 51, 839–848. [Google Scholar] [CrossRef]
- Turner, W. Sensing biodiversity. Science 2015, 346, 301–304. [Google Scholar] [CrossRef]
- Turner, W.; Rondinini, C.; Pettorelli, N.; Mora, B.; Leidner, A.K.; Szantoi, Z.; Buchanan, G.; Dech, S.; Dwyer, J.; Herold, M.; et al. Free and open-access satellite data are key to biodiversity conservation. Biol. Conserv. 2015, 182, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, G.M.; Nelson, A.; Mayaux, P.; Hartley, A.; Donald, P.F. Delivering a global, terrestrial, biodiversity observation system through remote sensing. Conserv. Biol. 2009, 23, 499–502. [Google Scholar] [CrossRef]
- Gholizadeh, H.; Gamon, J.A.; Townsend, P.A.; Zygielbaum, A.I.; Helzer, C.J.; Hmimina, G.Y.; Yu, R.; Moore, R.M.; Schweiger, A.K.; Cavender-Bares, J. Detecting prairie biodiversity with airborne remote sensing. Remote Sens. Environ. 2019, 221, 38–49. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Foody, G.M.; Rocchini, D.; Giorgi, A.P.; Saatchi, S. Measuring and modelling biodiversity from space. Prog. Phys. Geogr. Earth Environ. 2008, 32, 203–221. [Google Scholar] [CrossRef]
- Pettorelli, N.; Owen, H.J.F.; Duncan, C.; Freckleton, R. How do we want satellite remote sensing to support biodiversity conservation globally? Methods Ecol. Evol. 2016, 7, 656–665. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, Y.; Chen, W.; Zhao, Y.; Liu, X.; Bai, Y. The potential of mapping grassland plant diversity with the links among spectral diversity, functional trait diversity, and species diversity. Remote Sens. 2021, 13, 3034. [Google Scholar] [CrossRef]
- Rossi, C.; Kneubühler, M.; Schütz, M.; Schaepman, M.E.; Haller, R.M.; Risch, A.C.; Disney, M.; He, K. Spatial resolution, spectral metrics and biomass are key aspects in estimating plant species richness from spectral diversity in species-rich grasslands. Remote Sens. Ecol. Conserv. 2021, 1–18. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A.; Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Zygielbaum, A.I.; Kothari, S. Influence of species richness, evenness, and composition on optical diversity: A simulation study. Remote Sens. Environ. 2018, 211, 218–228. [Google Scholar] [CrossRef]
- Palmer, M.W.; Earls, P.G.; Hoagland, B.W.; White, P.S.; Wohlgemuth, T. Quantitative tools for perfecting species lists. Environmetrics 2002, 13, 121–137. [Google Scholar] [CrossRef]
- Rocchini, D.; Bacaro, G.; Chirici, G.; Da Re, D.; Feilhauer, H.; Foody, G.M.; Galluzzi, M.; Garzon-Lopez, C.X.; Gillespie, T.W.; He, K.S.; et al. Remotely sensed spatial heterogeneity as an exploratory tool for taxonomic and functional diversity study. Ecol. Indic. 2018, 85, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Schmidtlein, S.; Fassnacht, F.E. The spectral variability hypothesis does not hold across landscapes. Remote Sens. Environ. 2017, 192, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Ustin, S.L.; Gamon, J.A. Remote sensing of plant functional types. New Phytol. 2010, 186, 795–816. [Google Scholar] [CrossRef]
- Cho, M.A.; Mathieu, R.; Asner, G.P.; Naidoo, L.; van Aardt, J.; Ramoelo, A.; Debba, P.; Wessels, K.; Main, R.; Smit, I.P.J.; et al. Mapping tree species composition in south african savannas using an integrated airborne spectral and lidar system. Remote Sens. Environ. 2012, 125, 214–226. [Google Scholar] [CrossRef]
- Schneider, F.D.; Morsdorf, F.; Schmid, B.; Petchey, O.L.; Hueni, A.; Schimel, D.S.; Schaepman, M.E. Mapping functional diversity from remotely sensed morphological and physiological forest traits. Nat. Commun. 2017, 8, 1441–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zeng, Y.; Zheng, Z.; Dong, W.; Zhao, D.; Wu, B.; Zhao, Q. Forest species diversity mapping using airborne lidar and hyperspectral data in a subtropical forest in china. Remote Sens. Environ. 2018, 213, 104–114. [Google Scholar] [CrossRef]
- Zheng, Z.; Zeng, Y.; Schneider, F.D.; Zhao, Y.; Zhao, D.; Schmid, B.; Schaepman, M.E.; Morsdorf, F. Mapping functional diversity using individual tree-based morphological and physiological traits in a subtropical forest. Remote Sens. Environ. 2021, 252, 1–16. [Google Scholar] [CrossRef]
- Klingler, A.; Schaumberger, A.; Vuolo, F.; Kalmár, L.B.; Pötsch, E.M. Comparison of direct and indirect determination of leaf area index in permanent grassland. PFG—J. Photogramm. Remote Sens. Geoinf. Sci. 2020, 88, 369–378. [Google Scholar] [CrossRef]
- Lu, B.; He, Y. Species classification using unmanned aerial vehicle (uav)-acquired high spatial resolution imagery in a heterogeneous grassland. ISPRS J. Photogramm. Remote Sens. 2017, 128, 73–85. [Google Scholar] [CrossRef]
- Wachendorf, M.; Fricke, T.; Möckel, T. Remote sensing as a tool to assess botanical composition, structure, quantity and quality of temperate grasslands. Grass Forage Sci. 2018, 73, 1–14. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Surfus, J.A.; Serrano, L.; Surfus, J.S. The photochemical refectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef]
- Dahlin, K.M. Spectral diversity area relationships for assessing biodiversity in a wildland–agriculture matrix. Ecol. Appl. 2016, 26, 11. [Google Scholar] [CrossRef]
- Gholizadeh, H.; Gamon, J.A.; Zygielbaum, A.I.; Wang, R.; Schweiger, A.K.; Cavender-Bares, J. Remote sensing of biodiversity: Soil correction and data dimension reduction methods improve assessment of α-diversity (species richness) in prairie ecosystems. Remote Sens. Environ. 2018, 206, 240–253. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.; Emmerton, C.; Li, H.; Nestola, E.; Pastorello, G.; Menzer, O. Integrated analysis of productivity and biodiversity in a southern alberta prairie. Remote Sens. 2016, 8, 214. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Rivard, B.; Sánchez-Azofeifa, A.; Castro-Esau, K. Intra- and inter-class spectral variability of tropical tree species at la selva, costa rica: Implications for species identification using hydice imagery. Remote Sens. Environ. 2006, 105, 129–141. [Google Scholar] [CrossRef]
- Rocchini, D. Effects of spatial and spectral resolution in estimating ecosystem α-diversity by satellite imagery. Remote Sens. Environ. 2007, 111, 423–434. [Google Scholar] [CrossRef]
- Peng, Y.; Fan, M.; Bai, L.; Sang, W.; Feng, J.; Zhao, Z.; Tao, Z. Identification of the best hyperspectral indices in estimating plant species richness in sandy grasslands. Remote Sens. 2019, 11, 588. [Google Scholar] [CrossRef] [Green Version]
- Imran, H.A.; Gianelle, D.; Rocchini, D.; Dalponte, M.; Martín, M.P.; Sakowska, K.; Wohlfahrt, G.; Vescovo, L. Vis-nir, red-edge and nir-shoulder based normalized vegetation indices response to co-varying leaf and canopy structural traits in heterogeneous grasslands. Remote Sens. 2020, 12, 2254. [Google Scholar] [CrossRef]
- Gould, W. Remote sensing of vegetation, plant species richness, and regional biodiversity hotspots. Ecol. Appl. 2000, 10, 10. [Google Scholar] [CrossRef]
- Fauvel, M.; Lopes, M.; Dubo, T.; Rivers-Moore, J.; Frison, P.-L.; Gross, N.; Ouin, A. Prediction of plant diversity in grasslands using sentinel-1 and -2 satellite image time series. Remote Sens. Environ. 2020, 237, 111536. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant spectral diversity integrates functional and phylogenetic components of biodiversity and predicts ecosystem function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef]
- Lucas, K.; Carter, G. The use of hyperspectral remote sensing to assess vascular plant species richness on horn island, mississippi. Remote Sens. Environ. 2008, 112, 3908–3915. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, Y.; Lu, X.; Zhao, X.; Yang, L.; Sun, Z.; Bai, Y. Hyperspectral retrieval of leaf physiological traits and their links to ecosystem productivity in grassland monocultures. Ecol. Indic. 2021, 122, 107267. [Google Scholar] [CrossRef]
- Yu, X.; Guo, Q.; Chen, Q.; Guo, X. Discrimination of senescent vegetation cover from landsat-8 oli imagery by spectral unmixing in the northern mixed grasslands. Can. J. Remote Sens. 2019, 45, 192–208. [Google Scholar] [CrossRef]
- Zhou, X. Diversity of vascular plants in qinghai. J. Qinghai Environ. 2011, 4, 165–169. [Google Scholar] [CrossRef]
- Feng, M.; Che, X. Monthly surface water extent dataset for tibetan plateau and central asia (2000–2015). Natl. Tibet. Plateau Data Cent. 2019. [Google Scholar] [CrossRef]
- Li, C.; Peng, F.; Xue, X.; Lai, C.; Zhang, W.; You, Q.; Chen, X.; Zhang, X.; Wang, T. Degradation stage effects on vegetation and soil properties interactions in alpine steppe. J. Mt. Sci. 2021, 18, 646–657. [Google Scholar] [CrossRef]
- Wu, B.; Qian, J.; Zeng, Y. Land Cover Atlas of the People’s Re-Public of China (1:1,000,000); Sinomaps Press: Beijing, China, 2017. [Google Scholar]
- Yi, L.; Chen, J.M.; Zhang, G.; Xu, X.; Ming, X.; Guo, W. Seamless mosaicking of uav-based push-broom hyperspectral images for environment monitoring. Remote Sens. 2021, 13, 4720. [Google Scholar] [CrossRef]
- Lopatin, J.; Fassnacht, F.E.; Kattenborn, T.; Schmidtlein, S. Mapping plant species in mixed grassland communities using close range imaging spectroscopy. Remote Sens. Environ. 2017, 201, 12–23. [Google Scholar] [CrossRef]
- Oldeland, J.; Wesuls, D.; Rocchini, D.; Schmidt, M.; Jürgens, N. Does using species abundance data improve estimates of species diversity from remotely sensed spectral heterogeneity? Ecol. Indic. 2010, 10, 390–396. [Google Scholar] [CrossRef]
- Adams, J.B.; Smith, M.O.; Johnson, P.E. Spectral mixture modeling: A new analysis of rock and soil types at the viking lander 1 site. J. Geophys. Res. 1986, 91, 8098–8112. [Google Scholar] [CrossRef]
- Ding, Y.; Zheng, X.; Zhao, K.; Xin, X.; Liu, H. Quantifying the impact of ndvisoil determination methods and ndvisoil variability on the estimation of fractional vegetation cover in northeast china. Remote Sens. 2016, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Schaepman, M.E.; Wu, B.; Clevers, J.G.P.W.; Bregt, A.K. Forest structural variables retrieval using eo-1 hyperion data in combination with linear spectral unmixing and an inverted geometric-optical model. J. Remote Sens. 2007, 11, 648–658. [Google Scholar]
- Li, F.; Zeng, Y.; Luo, J.; Ma, R.; Wu, B. Modeling grassland aboveground biomass using a pure vegetation index. Ecol. Indic. 2016, 62, 279–288. [Google Scholar] [CrossRef]
- Boegh, E.; Houborg, R.; Bienkowski, J.; Braban, C.F.; Dalgaard, T.; van Dijk, N.; Dragosits, U.; Holmes, E.; Magliulo, V.; Schelde, K.; et al. Remote sensing of lai, chlorophyll and leaf nitrogen pools of crop- and grasslands in five european landscapes. Biogeosciences 2013, 10, 6279–6307. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.; Yu, H.; Du, R.; Wang, Q. Quantitative estimation of biomass of alpine grasslands using hyperspectral remote sensing. Rangel. Ecol. Manag. 2019, 72, 336–346. [Google Scholar] [CrossRef]
- Schiefer, F.; Schmidtlein, S.; Kattenborn, T. The retrieval of plant functional traits from canopy spectra through rtm-inversions and statistical models are both critically affected by plant phenology. Ecol. Indic. 2021, 121, 1–13. [Google Scholar] [CrossRef]
- Van Cleemput, E.; Vanierschot, L.; Fernández-Castilla, B.; Honnay, O.; Somers, B. The functional characterization of grass- and shrubland ecosystems using hyperspectral remote sensing: Trends, accuracy and moderating variables. Remote Sens. Environ. 2018, 209, 747–763. [Google Scholar] [CrossRef]
- Rossi, C.; Kneubühler, M.; Schütz, M.; Schaepman, M.E.; Haller, R.M.; Risch, A.C. From local to regional: Functional diversity in differently managed alpine grasslands. Remote Sens. Environ. 2020, 236, 111415. [Google Scholar] [CrossRef]
- Conti, L.; Malavasi, M.; Galland, T.; Komárek, J.; Lagner, O.; Carmona, C.P.; Bello, F.; Rocchini, D.; Šímová, P.; Feilhauer, H. The relationship between species and spectral diversity in grassland communities is mediated by their vertical complexity. Appl. Veg. Sci. 2021, 24. [Google Scholar] [CrossRef]
- Guimarães-Steinicke, C.; Weigelt, A.; Proulx, R.; Lanners, T.; Eisenhauer, N.; Duque-Lazo, J.; Reu, B.; Roscher, C.; Wagg, C.; Buchmann, N.; et al. Biodiversity facets affect community surface temperature via 3d canopy structure in grassland communities. J. Ecol. 2021, 109, 1969–1985. [Google Scholar] [CrossRef]
- Zhang, X.; Bao, Y.; Wang, D.; Xin, X.; Ding, L.; Xu, D.; Hou, L.; Shen, J. Using uav lidar to extract vegetation parameters of inner mongolian grassland. Remote Sens. 2021, 13, 656. [Google Scholar] [CrossRef]
- Zhao, D.; Pang, Y.; Li, Z.; Liu, L. Isolating individual trees in a closed coniferous forest using small footprint lidar data. Int. J. Remote Sens. 2014, 35, 7199–7218. [Google Scholar] [CrossRef]
- Zhao, D.; Pang, Y.; Li, Z.; Sun, G. Filling invalid values in a lidar-derived canopy height model with morphological crown control. Int. J. Remote Sens. 2013, 34, 4636–4654. [Google Scholar] [CrossRef]
- Ali, I.; Cawkwell, F.; Dwyer, E.; Barrett, B.; Green, S. Satellite remote sensing of grasslands: From observation to management. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar] [CrossRef] [Green Version]
- Rapinel, S.; Mony, C.; Lecoq, L.; Clément, B.; Thomas, A.; Hubert-Moy, L. Evaluation of sentinel-2 time-series for mapping floodplain grassland plant communities. Remote Sens. Environ. 2019, 223, 115–129. [Google Scholar] [CrossRef]
- Dolnik, C.; Breuer, M. Scale dependency in the species-area relationship of plant communities. Folia Geobot. 2008, 43, 305–318. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral Diversity | Soil Filtering | Species Richness | Shannon–Wiener Index | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | p-Value | RMSE | Bias | R2 | p-Value | RMSE | Bias | ||
| CVNDVI | With soil | 0.13 | 0.12 | 3.40 | 0.24 | 0.24 | 0.03 * | 0.39 | 0.08 |
| NDVI threshold | 0.29 | 0.02 * | 2.27 | 0.11 | 0.33 | 0.01 ** | 0.17 | 0.07 | |
| Unmixing | 0.28 | 0.007 ** | 2.25 | 0.09 | 0.37 | 0.002 ** | 0.08 | 0.04 | |
| CVMulti | With soil | 0.10 | 0.17 | 3.76 | 0.31 | 0.26 | 0.03 * | 0.15 | 0.07 |
| NDVI threshold | 0.34 | 0.02 * | 1.89 | 0.08 | 0.41 | 0.006 ** | 0.07 | 0.04 | |
| Unmixing | 0.40 | 0.002 ** | 1.74 | 0.07 | 0.61 | <0.001 ** | 0.04 | 0.02 | |
| CHA | With soil | 0.19 | 0.07 | 2.94 | 0.17 | 0.28 | 0.003 ** | 0.14 | 0.06 |
| NDVI threshold | 0.36 | 0.01 * | 1.88 | 0.08 | 0.44 | 0.005 ** | 0.07 | 0.03 | |
| Unmixing | 0.40 | 0.005 ** | 1.77 | 0.07 | 0.51 | 0.001 ** | 0.06 | 0.03 | |
| CHV | With soil | 0.16 | 0.08 | 3.24 | 0.20 | 0.24 | 0.04 * | 0.17 | 0.07 |
| NDVI threshold | 0.24 | 0.04 * | 2.40 | 0.12 | 0.37 | 0.009 ** | 0.09 | 0.04 | |
| Unmixing | 0.29 | 0.03 * | 2.18 | 0.10 | 0.41 | 0.007 ** | 0.08 | 0.03 | |
| Spectral Diversity | NDVI Threshold | Species Richness | Shannon–Wiener Index | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | p-Value | RMSE | Bias | R2 | p-Value | RMSE | Bias | ||
| CVNDVI | 0.2 | 0.27 * | 0.03 * | 5.16 | 0.11 | 0.33 * | 0.01 * | 0.17 | 0.08 |
| 0.3 | 0.27 * | 0.03 * | 5.15 | 0.11 | 0.33 ** | 0.009 ** | 0.18 | 0.08 | |
| 0.4 | 0.29 * | 0.02 * | 5.15 | 0.11 | 0.33 ** | 0.01 ** | 0.17 | 0.07 | |
| CVMulti | 0.2 | 0.27 * | 0.03 * | 3.98 | 0.10 | 0.41 ** | 0.006 ** | 0.08 | 0.04 |
| 0.3 | 0.28 * | 0.03 * | 3.90 | 0.10 | 0.41 ** | 0.006 ** | 0.08 | 0.04 | |
| 0.4 | 0.34 * | 0.02 * | 3.57 | 0.08 | 0.41 ** | 0.006 ** | 0.07 | 0.04 | |
| CHA | 0.2 | 0.28 * | 0.03 * | 4.13 | 0.10 | 0.42 ** | 0.004 ** | 0.08 | 0.03 |
| 0.3 | 0.33 * | 0.02 * | 3.86 | 0.09 | 0.40 ** | 0.005 ** | 0.09 | 0.04 | |
| 0.4 | 0.36 * | 0.01 * | 3.53 | 0.08 | 0.44 ** | 0.005 ** | 0.07 | 0.03 | |
| CHV | 0.2 | 0.22 * | 0.03 * | 6.04 | 0.13 | 0.35 ** | 0.008 ** | 0.08 | 0.04 |
| 0.3 | 0.24 * | 0.02 * | 5.74 | 0.11 | 0.35 ** | 0.009 ** | 0.08 | 0.04 | |
| 0.4 | 0.24 * | 0.04 * | 5.76 | 0.12 | 0.37 ** | 0.009 ** | 0.09 | 0.04 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Zeng, Y.; Zheng, Z.; Zhao, D.; Liu, W.; Ma, Z.; Wu, B. Assessing the Impact of Soil on Species Diversity Estimation Based on UAV Imaging Spectroscopy in a Natural Alpine Steppe. Remote Sens. 2022, 14, 671. https://doi.org/10.3390/rs14030671
Xu C, Zeng Y, Zheng Z, Zhao D, Liu W, Ma Z, Wu B. Assessing the Impact of Soil on Species Diversity Estimation Based on UAV Imaging Spectroscopy in a Natural Alpine Steppe. Remote Sensing. 2022; 14(3):671. https://doi.org/10.3390/rs14030671
Chicago/Turabian StyleXu, Cong, Yuan Zeng, Zhaoju Zheng, Dan Zhao, Wenjun Liu, Zonghan Ma, and Bingfang Wu. 2022. "Assessing the Impact of Soil on Species Diversity Estimation Based on UAV Imaging Spectroscopy in a Natural Alpine Steppe" Remote Sensing 14, no. 3: 671. https://doi.org/10.3390/rs14030671
APA StyleXu, C., Zeng, Y., Zheng, Z., Zhao, D., Liu, W., Ma, Z., & Wu, B. (2022). Assessing the Impact of Soil on Species Diversity Estimation Based on UAV Imaging Spectroscopy in a Natural Alpine Steppe. Remote Sensing, 14(3), 671. https://doi.org/10.3390/rs14030671