
Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Methods
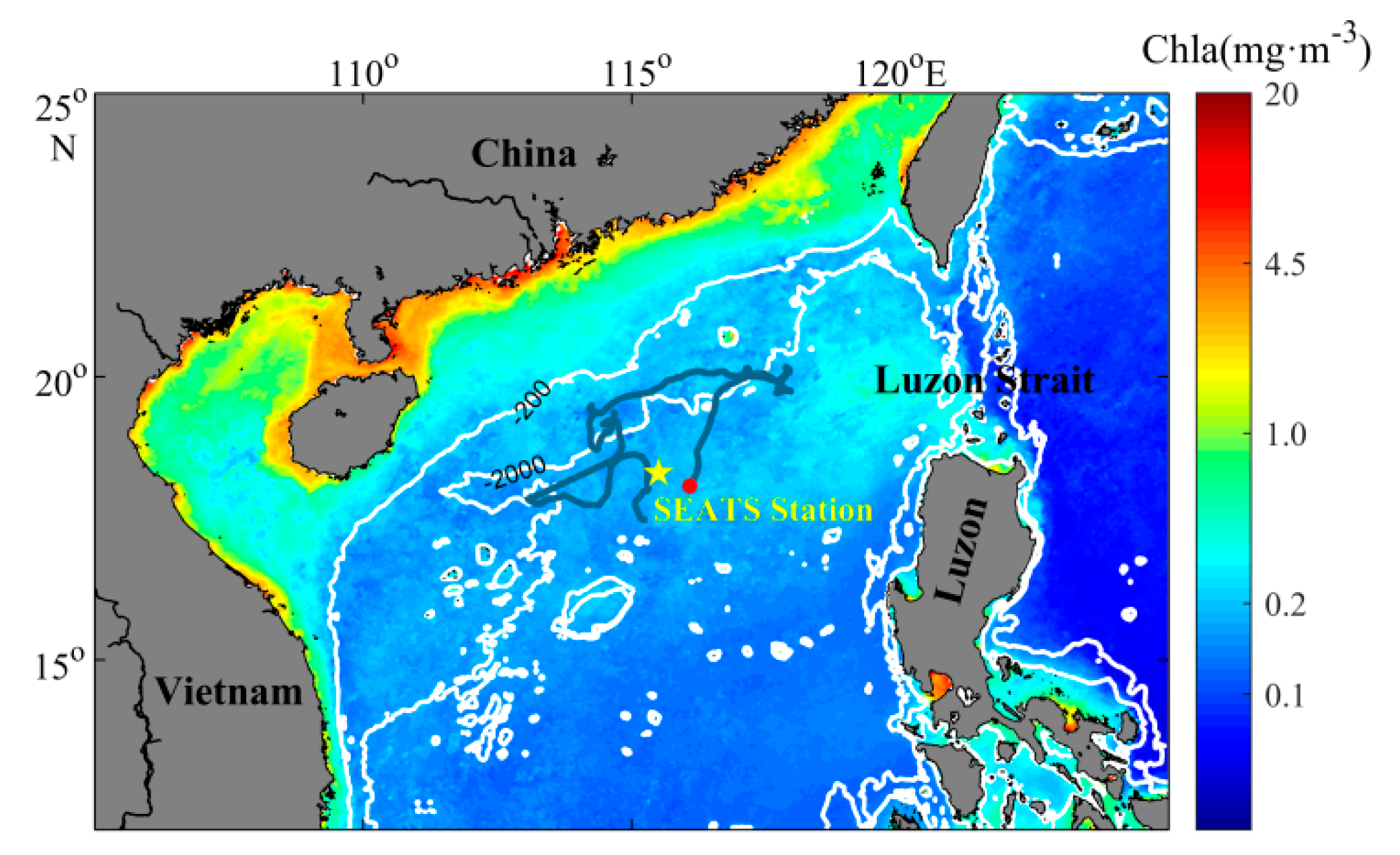
2.1. Model Description
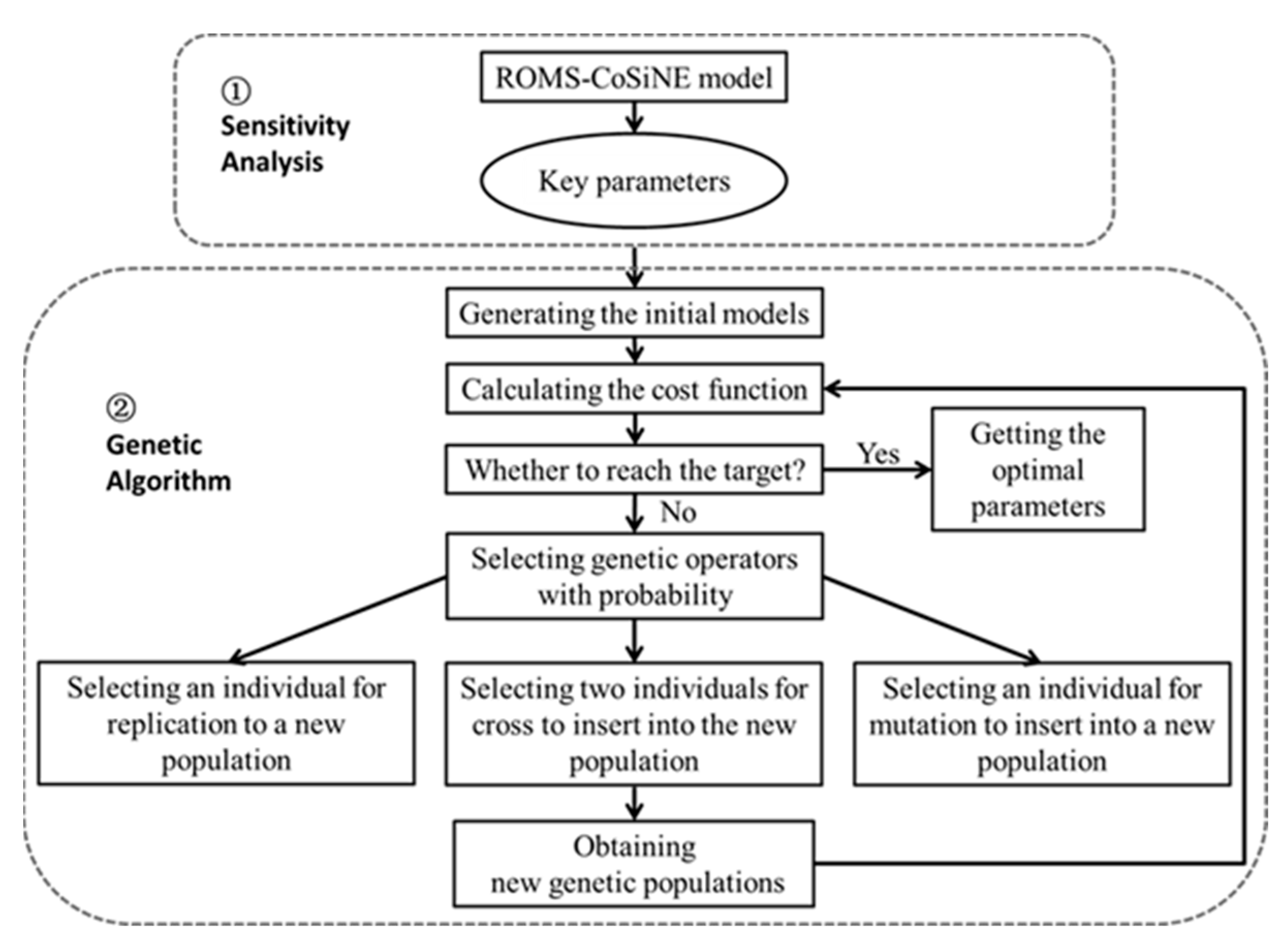
2.2. Sensitivity Analysis
2.3. Genetic Algorithm
2.4. Data and Optimization Experiments
3. Results
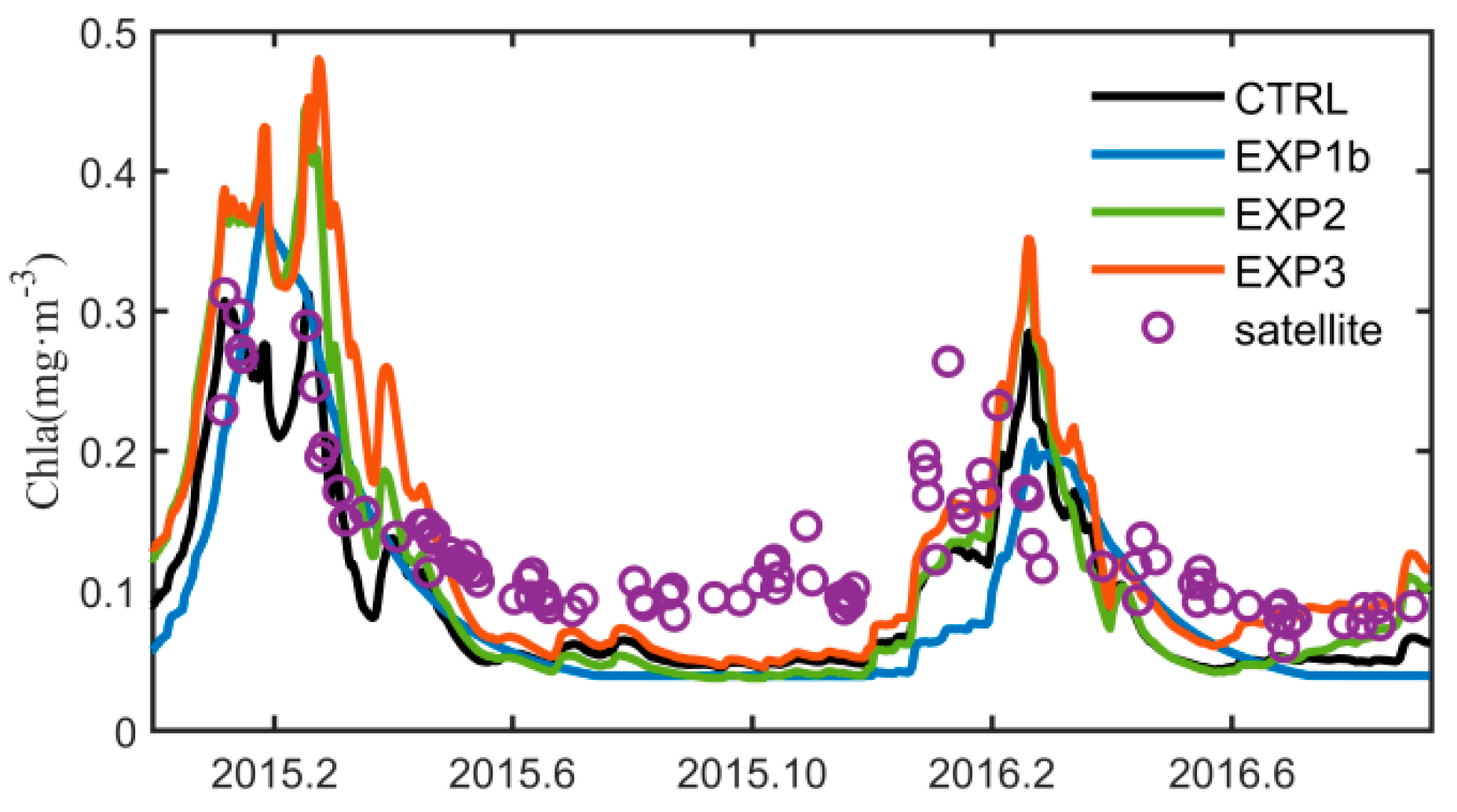
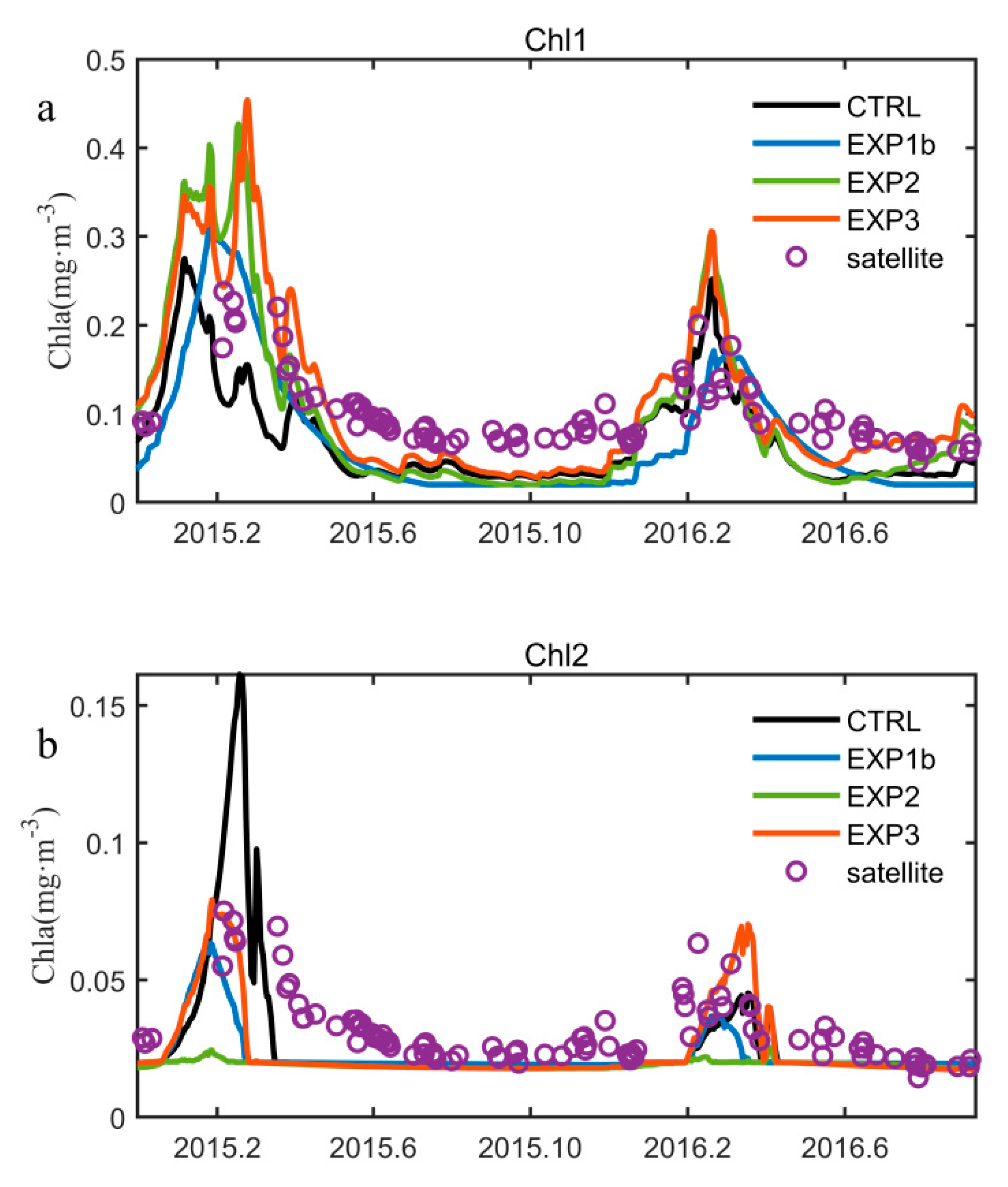
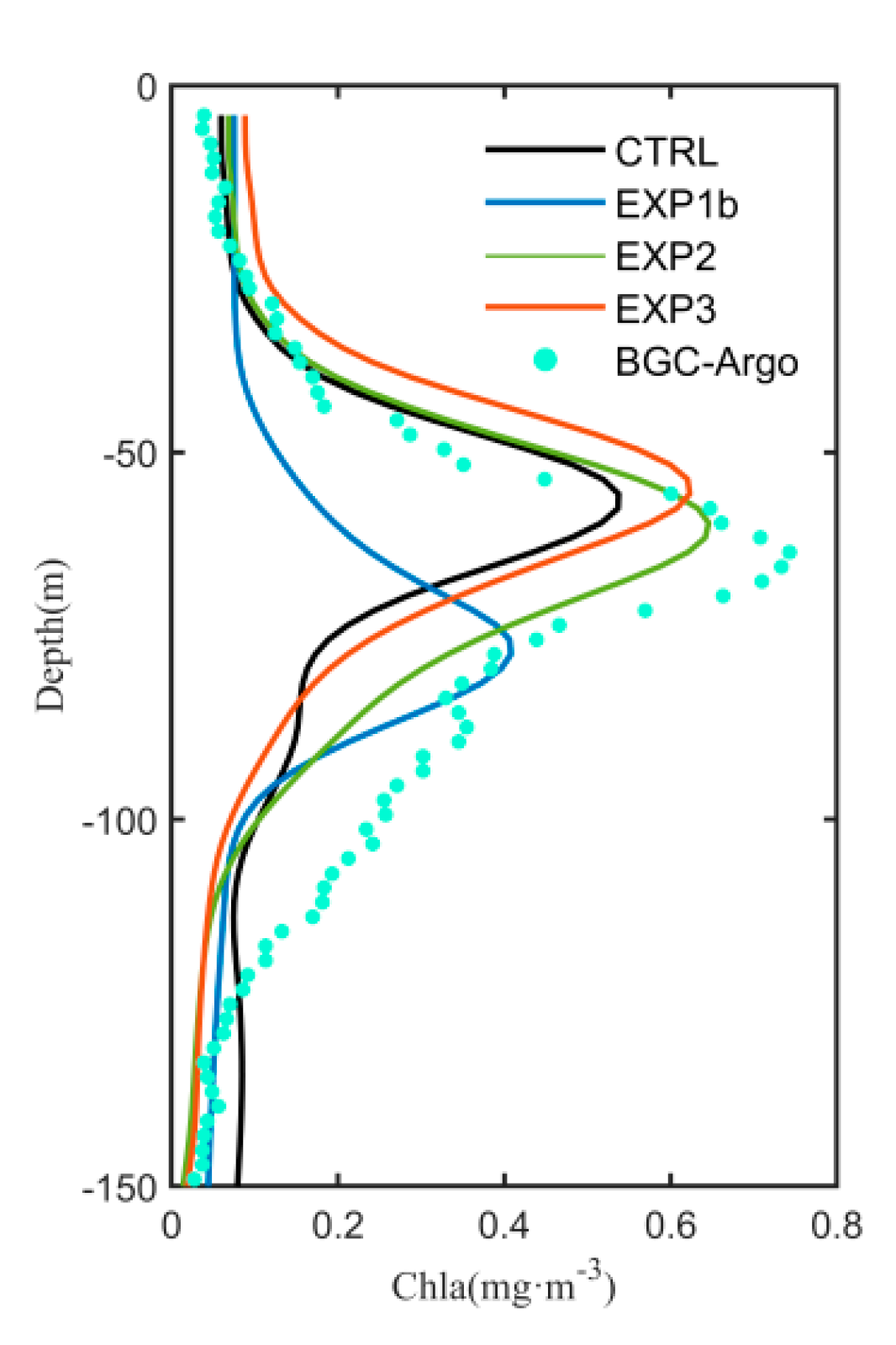
3.1. Seasonal Variation of Chlorophyll-a
3.2. Optimizable Parameter Selection
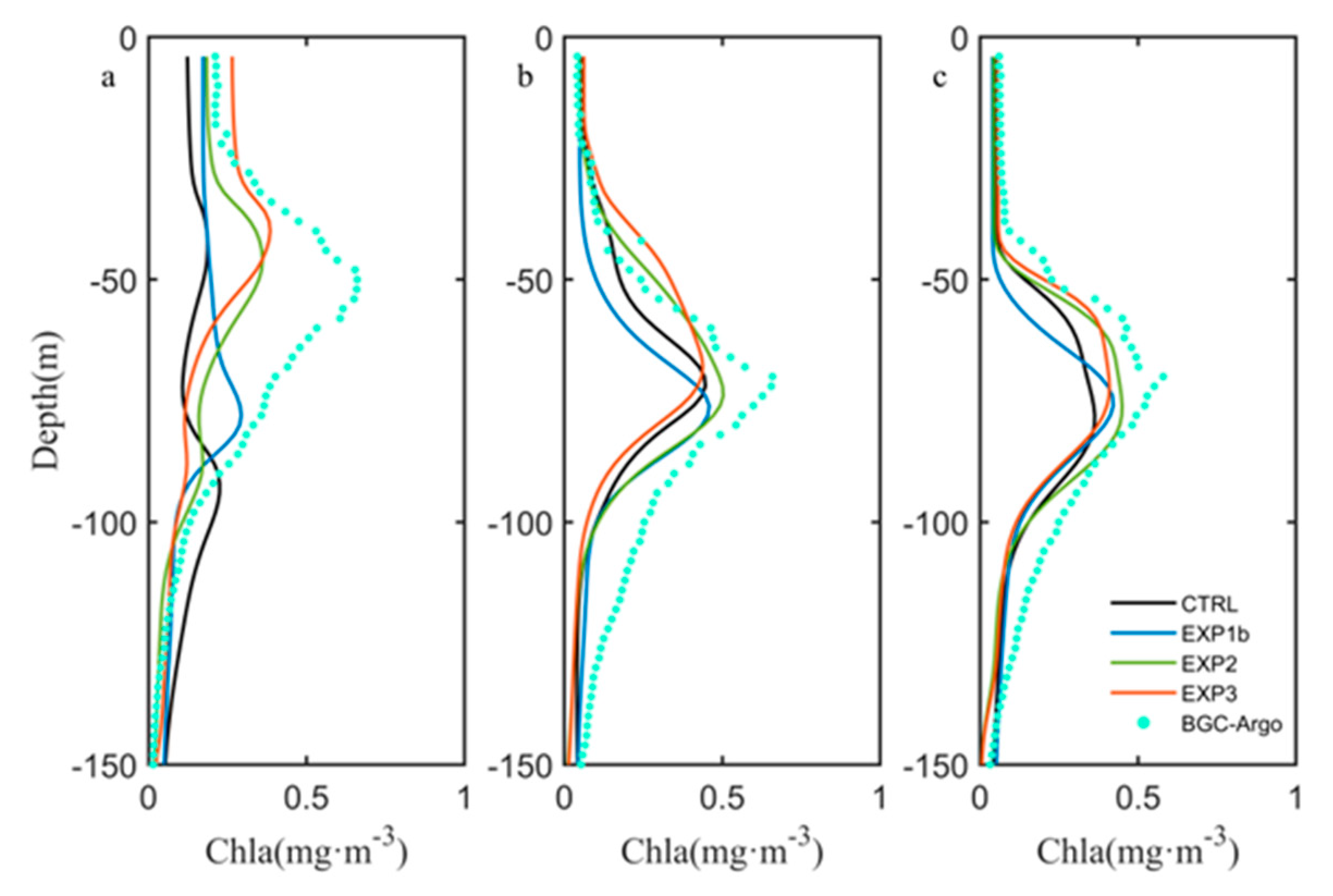
3.3. Optimization Results
4. Discussion
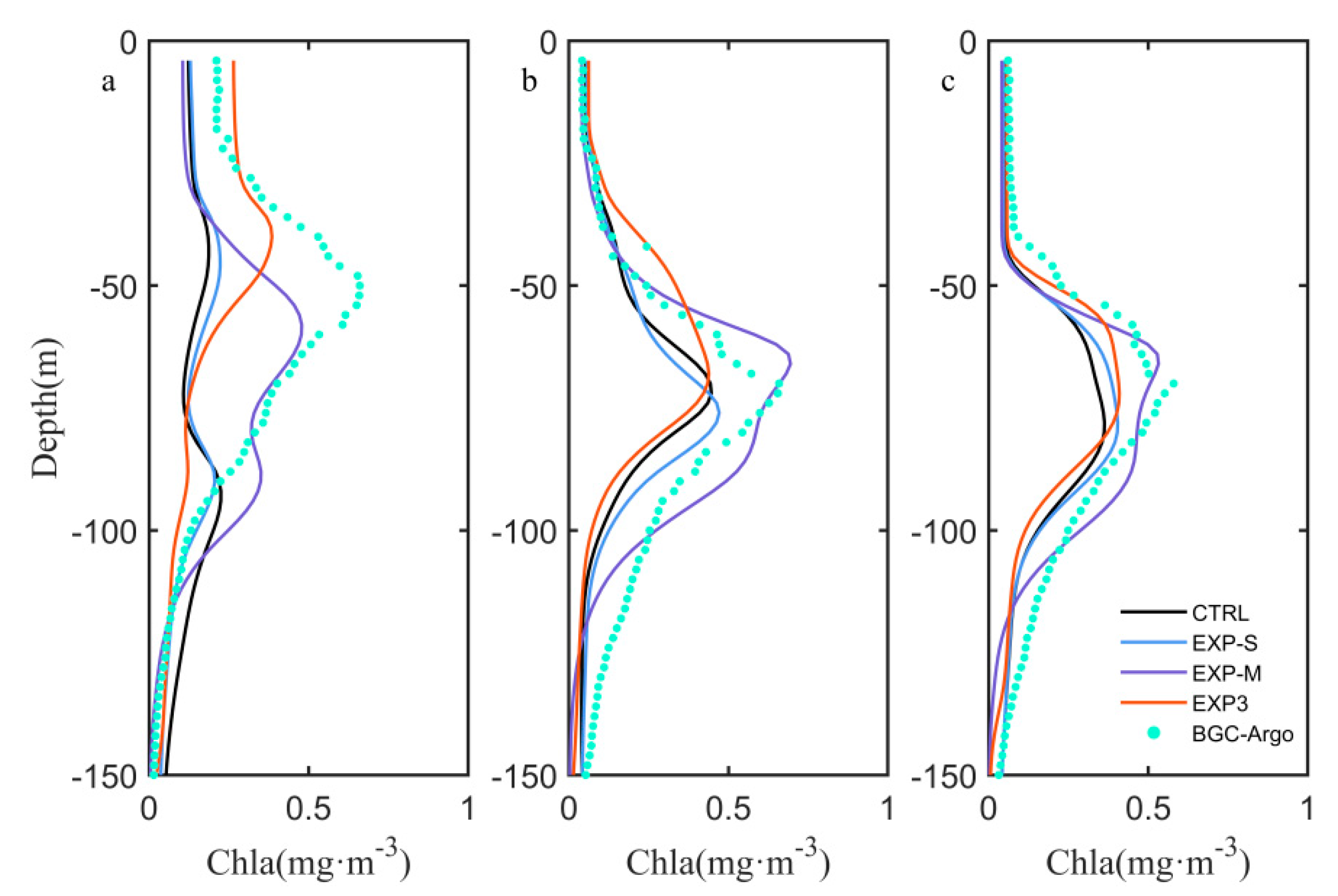
4.1. Influence of Sampling Frequency of Float Data
4.2. Effects of Biological Parameter on Vertical Chlorophyll-a Structure
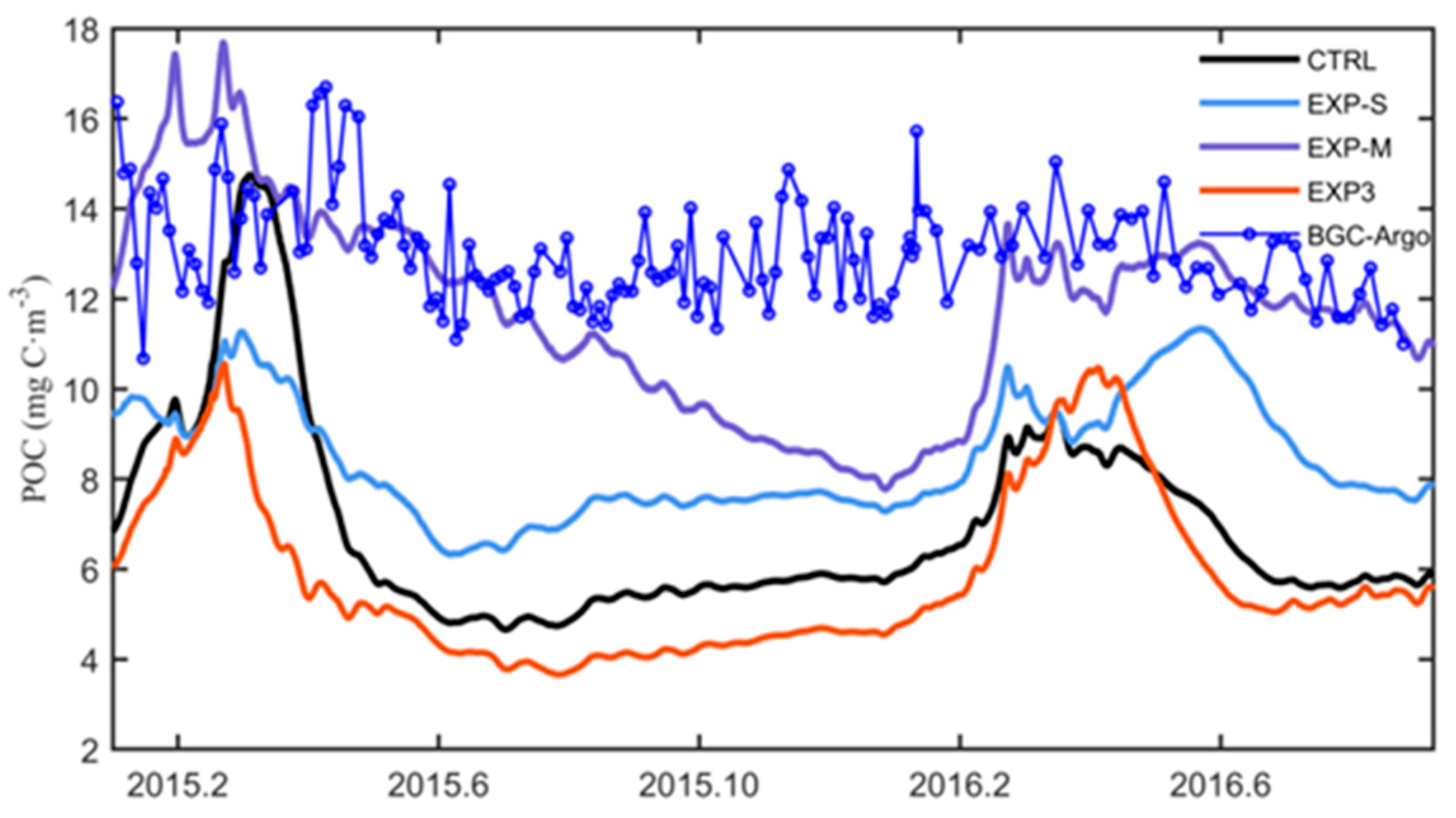
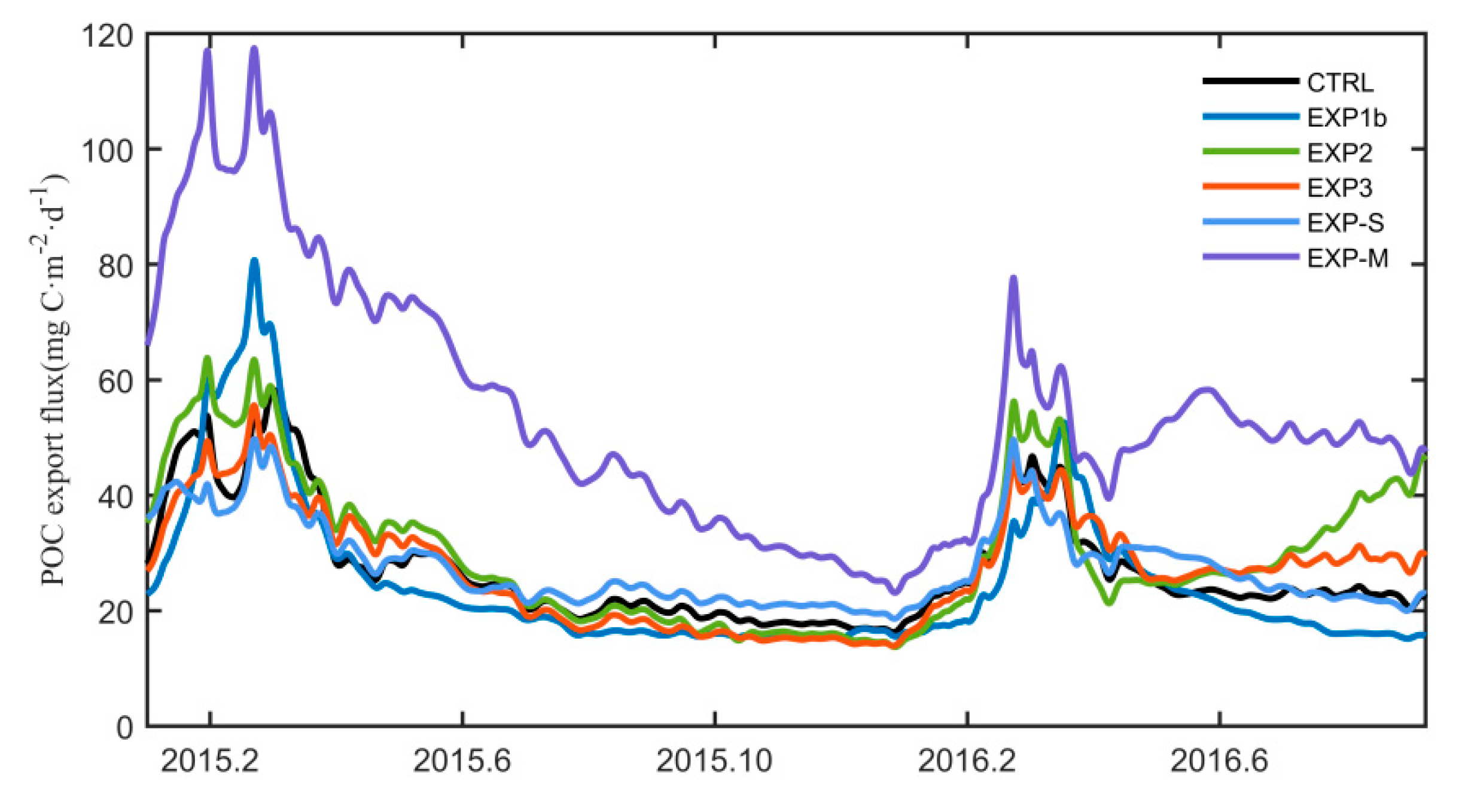
4.3. Impacts on Subsurface POC and Export Flux
5. Summary and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B
References
- Friedrichs, M.A.M.; Hood, R.R.; Wiggert, J.D. Ecosystem model complexity versus physical forcing: Quantification of their relative impact with assimilated Arabian Sea data. Deep Sea Res. Part II Top. Stud. Oceanogr. 2006, 53, 576–600. [Google Scholar] [CrossRef]
- Bisson, K.M.; Siegel, D.A.; Devries, T. How Data Set Characteristics Influence Ocean Carbon Export Models. Glob. Biogeochem. Cycles 2018, 32, 1312–1328. [Google Scholar] [CrossRef]
- Fennel, K.; Losch, M.; Schröter, J.; Wenzel, M. Testing a marine ecosystem model: Sensitivity analysis and parameter optimization. J. Mar. Syst. 2001, 28, 45–63. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, H.; Kishi, M.J. A data assimilation technique applied to estimate parameters for the NEMURO marine ecosystem model. Ecol. Model. 2004, 172, 69–85. [Google Scholar] [CrossRef]
- Dowd, M. Estimating parameters for a stochastic dynamic marine ecological system. Environmetrics 2011, 22, 501–515. [Google Scholar] [CrossRef]
- Mattern, J.P.; Dowd, M.; Fennel, K. Particle filter-based data assimilation for a three-dimensional biological ocean model and satellite observations. J. Geophys. Res. Ocean. 2013, 118, 2746–2760. [Google Scholar] [CrossRef]
- Xiao, Y.; Friedrichs, M.A.M. The assimilation of satellite-derived data into a one-dimensional lower trophic level marine ecosystem model. J. Geophys. Res. Ocean. 2014, 119, 2691–2712. [Google Scholar] [CrossRef] [Green Version]
- Gharamti, M.E.; Samuelsen, A.; Bertino, L.; Simon, E.; Korosov, A.; Daewel, U. Online tuning of ocean biogeochemical model parameters using ensemble estimation techniques: Application to a one-dimensional model in the North Atlantic. J. Mar. Syst. 2017, 168, 1–16. [Google Scholar] [CrossRef]
- Wang, B.; Fennel, K.; Yu, L.; Gordon, C. Assessing the value of biogeochemical argo profiles versus ocean color observations for biogeochemical model optimization in the Gulf of Mexico. Biogeosciences 2020, 17, 4059–4074. [Google Scholar] [CrossRef]
- Xue, H.; Chai, F.; Pettigrew, N.; Xu, D.; Shi, M.; Xu, J. Kuroshio intrusion and the circulation in the South China Sea. J. Geophys. Res. Ocean. 2004, 109, C02017. [Google Scholar] [CrossRef] [Green Version]
- Su, J. Overview of the South China Sea circulation and its influence on the coastal physical oceanography outside the Pearl River Estuary. Cont. Shelf Res. 2004, 24, 1745–1760. [Google Scholar] [CrossRef]
- Liu, K.K.; Chao, S.Y.; Shaw, P.T.; Gong, G.C.; Chen, C.C.; Tang, T.Y. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: Observations and a numerical study. Deep Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 1387–1412. [Google Scholar] [CrossRef]
- Ning, X.; Chai, F.; Xue, H.; Cai, Y.; Liu, C.; Zhu, G.; Shi, J. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea. J. Geophys. Res. Ocean 2004, 109, C10005. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Leptoukh, G.G.; Acker, J.G.; Yu, Z.; Kempler, S.J. Seasonal Variations of Chlorophyll a Concentration in the Northern South China Sea. IEEE Geosci. Remote Sens. Lett. 2008, 5, 315–319. [Google Scholar] [CrossRef]
- Tang, S.; Liu, F.; Chen, C. Seasonal and intraseasonal variability of surface chlorophyll a concentration in the South China Sea. Aquat. Ecosyst. Health Manag. 2014, 17, 242–251. [Google Scholar] [CrossRef]
- Geng, B.X.; Xiu, P.; Shu, C.; Zhang, W.Z.; Chai, F.; Li, S.; Wang, D. Evaluating the roles of wind- and buoyancy flux-induced mixing on phytoplankton dynamics in the northern and central South China Sea. J. Geophys. Res. Ocean. 2019, 124, 680–702. [Google Scholar] [CrossRef]
- Gong, X.; Shi, J.; Gao, H. Modeling seasonal variations of subsurface chlorophyll maximum in South China Sea. J. Ocean. Univ. China 2014, 13, 561–571. [Google Scholar] [CrossRef]
- Wang, S.; Li, S.; Hu, J.; Geng, B. Experiments in optimizing simulations of the subsurface chlorophyll maximum in the South China Sea. J. Mar. Syst. 2016, 156, 1–15. [Google Scholar] [CrossRef]
- Gong, X.; Jiang, W.; Wang, L.; Gao, H.; Boss, E.; Yao, X.; Kao, S.J.; Shi, J. Analytical solution of the nitracline with the evolution of subsurface chlorophyll maximum in stratified water columns. Biogeosciences 2017, 14, 2371–2386. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Hardman-Mountford, N.J.; Brewin, R.J.W. Synoptic relationships between surface Chlorophyll-a and diagnostic pigments specific to phytoplankton functional types. Biogeosciences 2011, 8, 311–327. [Google Scholar] [CrossRef] [Green Version]
- Sathyendranath, S.; Aiken, J.; Alvain, S. Phytoplankton Functional Types from Space; International Ocean-Colour Coordinating Group (IOCCG): Dartmouth, NS, Canada, 2014; p. 15. [Google Scholar]
- Kramer, S.J.; Roesler, C.S.; Sosik, H.M. Bio-optical discrimination of diatoms from other phytoplankton in the surface ocean: Evaluation and refinement of a model for the Northwest Atlantic. Remote Sens. Environ. 2018, 217, 126–143. [Google Scholar] [CrossRef]
- Kramer, S.J.; Siegel, D.A. How Can Phytoplankton Pigments Be Best Used to Characterize Surface Ocean Phytoplankton Groups for Ocean Color Remote Sensing Algorithms? J. Geophys. Res. Ocean. 2019, 124, 7557–7574. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Cao, W.; Wang, G.; Hu, S. Satellite-observed variability of phytoplankton size classes associated with a cold eddy in the South China Sea. Mar. Pollut. Bull. 2014, 83, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Brewin, R.J.W.; Stefano, C.; Shubha, S.; Thomas, J.; Gavin, T.; Kieran, C.; Airs, R.L.; Denise, C.; Vanda, B.; Emanuele, O. Uncertainty in ocean-Color estimates of chlorophyll for phytoplankton groups. Front. Mar. Sci. 2017, 4, 104. [Google Scholar] [CrossRef] [Green Version]
- Ciavatta, S.; Brewin, R.J.W.; Skákala, J.; Polimene, L.; De Mora, L.; Artioli, Y.; Allen, J.I. Assimilation of ocean-color plankton functional types to improve marine ecosystem simulations. J. Geophys. Res. Ocean. 2018, 123, 834–854. [Google Scholar] [CrossRef] [Green Version]
- Skákala, J.; Ford, D.; Brewin, R.J.W.; McEwan, R.; Kay, S.; Taylor, B.; de Mora, L.; Ciavatta, S. The assimilation of phytoplankton functional types for operational forecasting in the northwest European shelf. J. Geophys. Res. Ocean. 2018, 123, 5230–5247. [Google Scholar] [CrossRef] [Green Version]
- Ciavatta, S.; Kay, S.; Brewin, R.J.W.; Cox, R.; Di Cicco, A.; Nencioli, F.; Polimene, L.; Sammartino, M.; Santoleri, R.; Skákala, J. Ecoregions in the Mediterranean Sea through the reanalysis of phytoplankton functional types and carbon fluxes. J. Geophys. Res. Ocean. 2019, 124, 6737–6759. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, H.K.; Völker, C.; Losa, S.N.; Bracher, A.; Nerger, L. Global assimilation of ocean-color data of phytoplankton functional types: Impact of different data sets. J. Geophys. Res. Ocean. 2020, 125, e2019JC015586. [Google Scholar] [CrossRef] [Green Version]
- Hoshiba, Y.; Hirata, T.; Shigemitsu, M.; Nakano, H.; Hashioka, T.; Masuda, Y.; Yamanaka, Y. Biological data assimilation for parameter estimation of a phytoplankton functional type model for the western North Pacific. Ocean. Sci. 2018, 14, 371–386. [Google Scholar] [CrossRef]
- Kaufman, D.E.; Friedrichs, M.A.M.; Hemmings, J.C.P.; Smith, W.O., Jr. Assimilating bio-optical glider data during a phytoplankton bloom in the southern Ross Sea. Biogeosciences 2018, 15, 73–90. [Google Scholar] [CrossRef] [Green Version]
- Shchepetkin, A.F.; Mcwilliams, J.C. The regional oceanic modeling system (roms): A split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean. Model. 2005, 9, 347–404. [Google Scholar] [CrossRef]
- Chai, F.; Dugdale, R.C.; Peng, T.H.; Wilkerson, F.P.; Barber, R.T. One-dimensional ecosystem model of the equatorial Pacific upwelling system. Part I: Model development and silicon and nitrogen cycle. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 2713–2745. [Google Scholar] [CrossRef]
- Ma, W.; Xiu, P.; Chai, F.; Li, H. Seasonal variability of the carbon export in the central South China Sea. Ocean. Dyn. 2019, 69, 955–966. [Google Scholar] [CrossRef]
- Geider, R.J.; Macintyre, H.L.; Kana, T.M. Dynamic model of phytoplankton growth and acclimation: Responses of the balanced growth rate and the chlorophyll a:carbon ratio to light, nutrient-limitation and temperature. Mar. Ecol. Prog. Ser. 1997, 148, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Xiu, P.; Chai, F. Modeled biogeochemical responses to mesoscale eddies in the South China Sea. J. Geophys. Res. Ocean. 2011, 116, C10006. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Xiu, P.; Chai, F.; Xue, H.; Wang, D.; Sun, J. Enhanced chlorophyll concentrations induced by Kuroshio intrusion fronts in the northern South China Sea. Geophys. Res. Lett. 2017, 44, 11565–11572. [Google Scholar] [CrossRef] [Green Version]
- Geng, B.X.; Xiu, P.; Liu, N.; He, X.Q.; Chai, F. Biological response to the interaction of a mesoscale eddy and the river plume in the northern South China Sea. J. Geophys. Res. Ocean. 2021, 126, e2021JC017244. [Google Scholar] [CrossRef]
- Shaw, P.T.; Chao, S.Y.; Liu, K.K.; Pai, S.C.; Liu, C.T. Winter upwelling off Luzon in the northeastern South China Sea. J. Geophys. Res. Ocean. 1996, 101, 16435–16448. [Google Scholar] [CrossRef]
- Gan, J.; Li, L.; Wang, D.; Guo, X. Interaction of a river plume with coastal upwelling in the northeastern South China Sea. Cont. Shelf Res. 2009, 29, 728–740. [Google Scholar] [CrossRef]
- Jing, Z.Y.; Qi, Y.Q.; Hua, Z.L.; Zhang, H. Numerical study on the summer upwelling system in the northern continental shelf of the South China Sea. Cont. Shelf Res. 2009, 29, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Wong, G.T.F.; Tseng, C.M.; Wen, L.S.; Chung, S.W. Nutrient dynamics and N-anomaly at the SEATS station. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1528–1545. [Google Scholar] [CrossRef]
- Xiu, P.; Chai, F.; Shi, L.; Xue, H.; Chao, Y. A census of eddy activities in the South China Sea during 1993-2007. J. Geophys. Res. Ocean. 2010, 115, C03012. [Google Scholar] [CrossRef] [Green Version]
- Evans, G.T.; Parslow, J.S. A model of annual plankton cycles. Biol. Oceanogr. 1985, 3, 327–347. [Google Scholar] [CrossRef]
- Fasham, M.J.R. Variations in the seasonal cycle of biological production in subarctic oceans: A model sensitivity analysis. Deep Sea Res. Part I Oceanogr. Res. Pap. 1995, 42, 1111–1149. [Google Scholar] [CrossRef]
- Kishi, M.J.; Kashiwai, M.; Ware, D.M.; Megrey, B.A.; Eslinger, D.L.; Werner, F.E.; Noguchi-Aita, M.; Azumaya, T.; Fujii, M.; Hashimoto, S.; et al. NEMURO—a lower trophic level model for the North Pacific marine ecosystem. Ecol. Model. 2007, 202, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Hemmings, J.C.P.; Challenor, P.G.; Yool, A. Mechanistic site-based emulation of a global ocean biogeochemical model (MEDUSA 1.0) for parametric analysis and calibration: An application of the Marine Model Optimization Testbed (MarMOT 1.1). Geosci. Model. Dev. 2015, 8, 697–731. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Liu, G.; Gao, S.; Wang, H. Parameter sensitivity study of the biogeochemical model in the China coastal seas. Acta Oceanol. Sin. 2015, 34, 51–60. [Google Scholar] [CrossRef]
- Sankar, S.; Polimene, L.; Marin, L.; Menon, N.N.; Ciavatta, S. Sensitivity of the simulated Oxygen Minimum Zone to biogeochemical processes at an oligotrophic site in the Arabian Sea. Ecol. Model. 2018, 372, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Saltelli, A.; Ratto, M.; Andres, T.; Campolongo, F.; Tarantola, S. Global Sensitivity Analysis the Primer; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Schmitt, L.M. Fundamental study theory of genetic algorithms. Theor. Comput. Sci. 2001, 259, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Sathyendranath, S.; Brewin, R.; Brockmann, C.; Brotas, V.; Platt, T. An Ocean-Colour Time Series for Use in Climate Studies: The Experience of the Ocean-Colour Climate Change Initiative (OC-CCI). Sensors 2019, 19, 4285. [Google Scholar] [CrossRef] [Green Version]
- Boss, E.; Swift, D.; Taylor, L.; Brickley, P.; Zaneveld, R.; Riser, S.; Perry, M.J.; Strutton, P.G. Observations of pigment and particle distributions in the western North Atlantic from an autonomous float and ocean color satellite. Limnol. Oceanogr. 2008, 53, 2112–2122. [Google Scholar] [CrossRef] [Green Version]
- Cullen, J.J. The deep chlorophyll maximum: Comparing vertical profiles of chlorophyll a. Can. J. Fish. Aquat. Sci. 1982, 39, 791–803. [Google Scholar] [CrossRef]
- Xing, X.; Qiu, G.; Boss, E.; Wang, H. Temporal and vertical variations of particulate and dissolved optical properties in the South China Sea. J. Geophys. Res. Ocean. 2019, 124, 3779–3795. [Google Scholar] [CrossRef] [Green Version]
- Bisson, K.M.; Boss, E.; Westberry, T.K.; Behrenfeld, M.J. Evaluating satellite estimates of particulate backscatter in the global open ocean using autonomous profiling floats. Opt. Express 2019, 27, 30191–30203. [Google Scholar] [CrossRef] [PubMed]
- Haëntjens, N.; Della Penna, A.; Briggs, N.; Karp-Boss, L.; Gaube, P.; Claustre, H.; Boss, E. Detecting mesopelagic organisms using biogeochemical-Argo floats. Geophys. Res. Lett. 2020, 47, e2019GL086088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stramski, D.; Reynolds, R.A.; Babin, M.; Kaczmarek, S.; Lewis, M.R.; Röttgers, R.; Sciandra, A.; Stramska, M.; Twardowski, M.S.; Franz, B.A.; et al. Relationships between the surface concentration of particulate organic carbon and optical properties in the eastern South Pacific and eastern Atlantic Oceans. Biogeosciences 2008, 5, 171–201. [Google Scholar] [CrossRef] [Green Version]
- Qiu, G.; Xing, X.; Boss, E.S.; Yan, X.H.; Wang, H. Relationships between optical backscattering, particulate organic carbon, and phytoplankton carbon in the oligotrophic South China Sea basin. Opt. Express 2021, 29, 15159–15176. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.M.; Wong, G.T.F.; Lin, I.I.; Wu, C.R.; Liu, K.K. A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea. Geophys. Res. Lett. 2005, 32, L08608. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.Z.; Wang, H.; Chai, F.; Qiu, G. Physical drivers of chlorophyll variability in the open South China Sea. J. Geophys. Res. Ocean. 2016, 121, 7123–7140. [Google Scholar] [CrossRef]
- Bisson, K.M.; Boss, E.; Werdell, P.J. Seasonal bias in global ocean color observations. Appl. Opt. 2021, 60, 6978–6988. [Google Scholar] [CrossRef]
- Park, M.-S.; Lee, S.; Ahn, J.-H.; Lee, S.-J.; Choi, J.-K.; Ryu, J.-H. Decadal Measurements of the First Geostationary Ocean Color Satellite (GOCI) Compared with MODIS and VIIRS Data. Remote Sens. 2022, 14, 72. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, H.-Y.; Karl, D.M.; Takahashi, M. Nitrogen modulates phytoplankton growth in spring in the South China Sea. Cont. Shelf Res. 2004, 24, 527–541. [Google Scholar] [CrossRef]
- Chen, C.-C.; Shiah, F.-K.; Chung, S.-W.; Liu, K.-K. Winter phytoplankton blooms in the shallow mixed layer of the South China Sea enhanced by upwelling. J. Mar. Syst. 2006, 59, 97–110. [Google Scholar] [CrossRef]
- Huang, B.; Hu, J.; Xu, H.; Cao, Z.; Wang, D. Phytoplankton community at warm eddies in the northern South China Sea in winter 2003/2004. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1792–1798. [Google Scholar] [CrossRef]
- Beckmann, A.; Hense, I. Beneath the surface: Characteristics of oceanic ecosystems under weak mixing conditions—A theoretical investigation. Prog. Oceanogr. 2007, 75, 771–796. [Google Scholar] [CrossRef]
- De La Rocha, C.L. The Biological Pump. In Treatise on Geochemistry; Elsevier: Oxford, UK, 2007; pp. 1–29. [Google Scholar]
- Zhou, K.; Dai, M.; Maiti, K.; Chen, W.; Xie, Y. Impact of physical and biogeochemical forcing on particle export in the South China Sea. Prog. Oceanogr. 2020, 187, 102403. [Google Scholar] [CrossRef]
- Cai, P.; Zhao, D.; Wang, L.; Huang, B.; Dai, M. Role of particle stock and phytoplankton community structure in regulating particulate organic carbon export in a large marginal sea. J. Geophys. Res. Ocean. 2015, 120, 2063–2095. [Google Scholar] [CrossRef]
- Mouw, C.B.; Barnett, A.; McKinley, G.A.; Gloege, L.; Pilcher, D. Phytoplankton size impact on export flux in the global ocean. Glob. Biogeochem. Cycles 2016, 30, 1542–1562. [Google Scholar] [CrossRef] [Green Version]
- Siegel, D.A.; Buesseler, K.O.; Doney, S.C.; Sailley, S.F.; Behrenfeld, M.J.; Boyd, P.W. Global assessment of ocean carbon export by combining satellite observations and food-web models. Glob. Biogeochem. Cycles 2014, 28, 181–196. [Google Scholar] [CrossRef]
- Garcia-Gorriz, E.; Hoepffner, N.; Ouberdous, M. Assimilation of SeaWiFS data in a coupled physical–biological model of the Adriatic Sea. J. Mar. Syst. 2003, 40, 233–252. [Google Scholar] [CrossRef]
- Tjiputra, J.F.; Polzin, D.; Winguth, A.M.E. Assimilation of seasonal chlorophyll and nutrient data into an adjoint three-dimensional ocean carbon cycle model: Sensitivity analysis and ecosystem parameter optimization. Glob. Biogeochem. Cycles 2007, 21, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Lv, X. Data assimilation in a simple marine ecosystem model based on spatial biological parameterizations. Ecol. Model. 2009, 220, 1997–2008. [Google Scholar] [CrossRef]
- Xiao, Y.; Friedrichs, M.A.M. Using biogeochemical data assimilation to assess the relative skill of multiple ecosystem models in the Mid-Atlantic Bight: Effects of increasing the complexity of the planktonic food web. Biogeosciences 2014, 11, 3015–3030. [Google Scholar] [CrossRef] [Green Version]
- Brewin, R.J.W.; Sathyendranath, S.; Hirata, T.; Lavender, S.J.; Barciela, R.M.; HardmanMountford, N.J. A three-component model of phytoplankton size class for the Atlantic Ocean. Ecol. Model. 2010, 221, 1472–1483. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Observation Data |
|---|---|
| CTRL | - |
| EXP1a | satellite sea surface chlorophyll-a |
| EXP1b | satellite-derived PFT data |
| EXP2 | BGC-Argo profiles of chlorophyll-a |
| EXP3 | PFT data and BGC-Argo profiles of chlorophyll-a |
| EXP-S | PFT data and seasonal averaged BGC-Argo profiles of chlorophyll-a |
| EXP-M | PFT data and monthly averaged BGC-Argo profiles of chlorophyll-a |
| Parameter | Description | Initial Value | Minimum | Maximum | Unit | r2 |
|---|---|---|---|---|---|---|
| reg1 | Z1 excretion rate to ammonium | 0.1 | 0.05 | 0.2 | day−1 | 0.083 |
| gmaxs1 | maximum specific growth rate of S1 | 2.0 | 1.0 | 4.0 | day−1 | 0.018 |
| beta1 | Z1 maximum grazing rate | 0.8 | 0.4 | 1.0 | day−1 | 0.099 |
| beta2 | Z2 maximum grazing rate | 0.4 | 0.2 | 0.8 | day−1 | 0.047 |
| akz2 | half saturation for Z2 grazing | 0.25 | 0.125 | 0.5 | mmol N m−3 | 0.025 |
| amaxs1 | initial slope of P-I curve of S1 | 0.025 | 0.0125 | 0.05 | (W m−2 day)−1 | 0.075 |
| akno3s2 | half saturation of nitrate uptake by S2 | 2.0 | 1.0 | 4.0 | mmol N m−3 | 0.046 |
| bgamma1 | grazing efficiency of Z1 | 0.75 | 0.375 | 1.0 | day−1 | 0.139 |
| Chl2cs2_m | maximum chlorophyll-a to carbon ratio for S2 | 0.065 | 0.03 | 0.08 | mg Chla (mg C)−1 | 0.056 |
| Item | Fs1 | Fs2 | Fv | Fscm |
|---|---|---|---|---|
| CTRL | 0.1945 | 0.0333 | 3.2583 | 0.0542 |
| EXP1a | 0.1782 | 0.0293 | 3.5846 | 0.0477 |
| EXP1b | 0.1424 | 0.0169 | 3.3784 | 0.0516 |
| EXP2 | 0.2020 | 0.0279 | 2.5939 | 0.0368 |
| EXP3 | 0.1600 | 0.0155 | 2.8763 | 0.0404 |
| Item | Fv | Fscm | Fv-s | Fv-m |
|---|---|---|---|---|
| CTRL | 3.2583 | 0.0542 | 1.0583 | 0.9097 |
| EXP-S | 3.0348 | 0.0419 | 0.9812 | 0.8065 |
| EXP-M | 2.6264 | 0.0292 | 0.7438 | 0.5916 |
| EXP3 | 2.8763 | 0.0404 | 0.5826 | 0.6877 |
| Parameter | reg1 | gmaxs1 | beta1 | beta2 | akz2 | amaxs1 | akno3s2 | bgamma1 | Chl2cs2_m |
|---|---|---|---|---|---|---|---|---|---|
| CTRL | 0.1 | 2.0 | 0.8 | 0.4 | 0.25 | 0.025 | 2.0 | 0.75 | 0.065 |
| EXP1a | 0.096 | 2.412 | 0.573 | 0.605 | 0.436 | 0.024 | 1.536 | 0.539 | 0.041 |
| EXP1b | 0.124 | 3.027 | 0.518 | 0.423 | 0.395 | 0.016 | 1.371 | 0.912 | 0.045 |
| EXP2 | 0.118 | 2.303 | 0.679 | 0.649 | 0.384 | 0.026 | 1.647 | 0.821 | 0.057 |
| EXP3 | 0.12 | 1.914 | 0.702 | 0.672 | 0.372 | 0.021 | 1.372 | 0.874 | 0.056 |
| EXP-S | 0.151 | 1.937 | 0.916 | 0.405 | 0.414 | 0.027 | 2.587 | 0.876 | 0.05 |
| EXP-M | 0.092 | 3.471 | 0.624 | 0.468 | 0.305 | 0.048 | 1.05 | 0.743 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, C.; Xiu, P.; Xing, X.; Qiu, G.; Ma, W.; Brewin, R.J.W.; Ciavatta, S. Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea. Remote Sens. 2022, 14, 1297. https://doi.org/10.3390/rs14051297
Shu C, Xiu P, Xing X, Qiu G, Ma W, Brewin RJW, Ciavatta S. Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea. Remote Sensing. 2022; 14(5):1297. https://doi.org/10.3390/rs14051297
Chicago/Turabian StyleShu, Chan, Peng Xiu, Xiaogang Xing, Guoqiang Qiu, Wentao Ma, Robert J. W. Brewin, and Stefano Ciavatta. 2022. "Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea" Remote Sensing 14, no. 5: 1297. https://doi.org/10.3390/rs14051297
APA StyleShu, C., Xiu, P., Xing, X., Qiu, G., Ma, W., Brewin, R. J. W., & Ciavatta, S. (2022). Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea. Remote Sensing, 14(5), 1297. https://doi.org/10.3390/rs14051297