
Identifying the Factors behind Climate Diversification and Refugial Capacity in Mountain Landscapes: The Key Role of Forests



Abstract
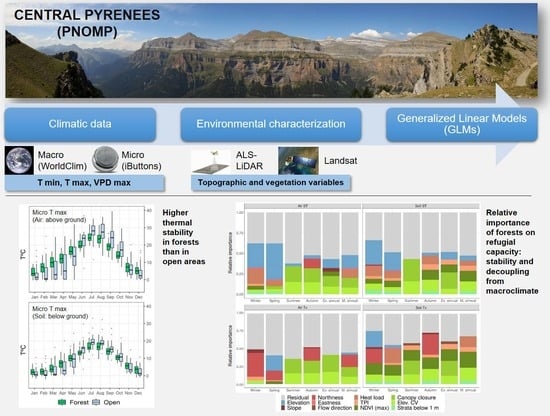
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Macro- and Microclimate Data Acquisition
2.3. Microclimatic Data-Processing
2.4. Environmental Variables
2.5. Statistical Modeling
3. Results
3.1. Climatic Diversity and Performance of Microclimatic Models
3.2. Variables behind Microclimatic Diversity (Tn, Tx, and VPDmax)
3.3. Open Versus Forested Microclimates
3.4. Factors Promoting Refugial Capacity
4. Discussion
4.1. Climatic Diversification: Beyond the Effect of Elevation
4.2. Refugial Capacity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peñuelas, J.; Sardans, J.; Estiarte, M.; Ogaya, R.; Carnicer, J.; Coll, M.; Barbeta, A.; Rivas-Ubach, A.; Llusià, J.; Garbulsky, M.; et al. Evidence of Current Impact of Climate Change on Life: A Walk from Genes to the Biosphere. Glob. Chang. Biol. 2013, 19, 2303–2338. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, B.R.; Meester, L.D.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The Broad Footprint of Climate Change from Genes to Biomes to People. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Continent-Wide Response of Mountain Vegetation to Climate Change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction Debt of High-Mountain Plants under Twenty-First-Century Climate Change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The Velocity of Climate Change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef]
- Sandel, B.; Arge, L.; Dalsgaard, B.; Davies, R.G.; Gaston, K.J.; Sutherland, W.J.; Svenning, J.C. The Influence of Late Quaternary Climate-Change Velocity on Species Endemism. Science 2011, 334, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Körner, C. Infra-red Thermometry of Alpine Landscapes Challenges Climatic Warming Projections. Glob. Ecol. Biogeogr. 2010, 16, 2602–2613. [Google Scholar] [CrossRef]
- Scherrer, D.; Körner, C. Topographically Controlled Thermal-Habitat Differentiation Buffers Alpine Plant Diversity against Climate Warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Suggitt, A.J.; Wilson, R.J.; Isaac, N.J.B.; Beale, C.M.; Auffret, A.G.; August, T.; Bennie, J.J.; Crick, H.Q.P.; Duffield, S.; Fox, R.; et al. Extinction Risk from Climate Change Is Reduced by Microclimatic Buffering. Nat. Clim. Chang. 2018, 8, 713–717. [Google Scholar] [CrossRef]
- Dobrowski, S.Z. A Climatic Basis for Microrefugia: The Influence of Terrain on Climate. Glob. Chang. Biol. 2011, 17, 1022–1035. [Google Scholar] [CrossRef]
- Ashcroft, M.B. Identifying Refugia from Climate Change. J. Biogeogr. 2010, 37, 1407–1413. [Google Scholar] [CrossRef]
- Gentili, R.; Baroni, C.; Caccianiga, M.; Armiraglio, S.; Ghiani, A.; Citterio, S. Potential Warm-Stage Microrefugia for Alpine Plants: Feedback between Geomorphological and Biological Processes. Ecol. Complex. 2015, 21, 87–99. [Google Scholar] [CrossRef]
- Brighenti, S.; Hotaling, S.; Finn, D.S.; Fountain, A.G.; Hayashi, M.; Herbst, D.; Saros, J.E.; Tronstad, L.M.; Millar, C.I. Rock Glaciers and Related Cold Rocky Landforms: Overlooked Climate Refugia for Mountain Biodiversity. Glob. Chang. Biol. 2021, 27, 1504–1517. [Google Scholar] [CrossRef]
- Boehnke, D. Exploring the Thermal Microcosms at the Forest Floor—A Case Study of a Temperate Forest. Atmos. Basel 2021, 12, 503. [Google Scholar] [CrossRef]
- Rita, A.; Bonanomi, G.; Allevato, E.; Borghetti, M.; Cesarano, G.; Mogavero, V.; Rossi, S.; Saulino, L.; Zotti, M.; Saracino, A. Topography Modulates Near-Ground Microclimate in the Mediterranean Fagus Sylvatica Treeline. Sci. Rep. UK 2021, 11, 8122. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, D.M.; Iversen, L.L.; Ehrlén, J.; Hylander, K. Changes in Forest Structure Drive Temperature Preferences of Boreal Understorey Plant Communities. J. Ecol. 2021, 110, 631–643. [Google Scholar] [CrossRef]
- Greiser, C.; Meineri, E.; Luoto, M.; Ehrlén, J.; Hylander, K. Monthly Microclimate Models in a Managed Boreal Forest Landscape. Agric. For. Meteorol. 2018, 250, 147–158. [Google Scholar] [CrossRef]
- Wolf, C.; Bell, D.M.; Kim, H.; Nelson, M.P.; Schulze, M.; Betts, M.G. Temporal Consistency of Undercanopy Thermal Refugia in Old-Growth Forest. Agric. For. Meteorol. 2021, 307, 108520. [Google Scholar] [CrossRef]
- Rull, V. Microrefugia. J. Biogeogr. 2009, 36, 481–484. [Google Scholar] [CrossRef]
- Reside, A.E.; Van Der Wal, J.; Phillips, B.L.; Shoo, L.P. Climate Change Refugia for Terrestrial Biodiversity. Austral Ecol. 2014, 39, 887–897. [Google Scholar] [CrossRef]
- Gavin, D.G.; Fitzpatrick, M.C.; Gugger, P.F.; Heath, K.D.; Sánchez, F.R.; DOBROWSKI, S.Z.; Hampe, A.; Hu, F.S.; Ashcroft, M.B.; Bartlein, P.J.; et al. Climate Refugia: Joint Inference from Fossil Records, Species Distribution Models and Phylogeography. New Phytol. 2014, 204, 37–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclean, I.M.D.; Hopkins, J.J.; Bennie, J.; Lawson, C.R.; Wilson, R.J. Microclimates Buffer the Responses of Plant Communities to Climate Change. Glob. Ecol. Biogeogr. 2015, 24, 1340–1350. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing Climate Change Refugia for Climate Adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keppel, G.; Wardell-Johnson, G.W. Refugia: Keys to Climate Change Management. Glob. Chang. Biol. 2012, 18, 2389–2391. [Google Scholar] [CrossRef]
- Keppel, G.; Mokany, K.; Wardell-Johnson, G.W.; Phillips, B.L.; Welbergen, J.A.; Reside, A.E. The Capacity of Refugia for Conservation Planning under Climate Change. Front. Ecol. Environ. 2015, 13, 106–112. [Google Scholar] [CrossRef]
- Morelli, T.L.; Barrows, C.W.; Ramirez, A.R.; Cartwright, J.M.; Ackerly, D.D.; Eaves, T.D.; Ebersole, J.L.; Krawchuk, M.A.; Letcher, B.H.; Mahalovich, M.F.; et al. Climate-Change Refugia: Biodiversity in the Slow Lane. Front. Ecol. Environ. 2020, 18, 228–234. [Google Scholar] [CrossRef]
- von Arx, G.; Pannatier, E.G.; Thimonier, A.; Rebetez, M. Microclimate in Forests with Varying Leaf Area Index and Soil Moisture: Potential Implications for Seedling Establishment in a Changing Climate. J. Ecol. 2013, 101, 1201–1213. [Google Scholar] [CrossRef]
- Ashcroft, M.B.; Gollan, J.R. Moisture, Thermal Inertia, and the Spatial Distributions of near-Surface Soil and Air Temperatures: Understanding Factors That Promote Microrefugia. Agric. For. Meteorol. 2013, 176, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Maclean, I.M.D.; Suggitt, A.J.; Wilson, R.J.; Duffy, J.P.; Bennie, J.J. Fine-Scale Climate Change: Modelling Spatial Variation in Biologically Meaningful Rates of Warming. Glob. Chang. Biol. 2017, 23, 256–268. [Google Scholar] [CrossRef]
- Davis, K.T.; Dobrowski, S.Z.; Holden, Z.A.; Higuera, P.E.; Abatzoglou, J.T. Microclimatic Buffering in Forests of the Future: The Role of Local Water Balance. Ecography 2019, 42, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Macek, M.; Kopecký, M.; Wild, J. Maximum Air Temperature Controlled by Landscape Topography Affects Plant Species Composition in Temperate Forests. Landscape Ecol. 2019, 34, 2541–2556. [Google Scholar] [CrossRef]
- Williamson, J.; Slade, E.M.; Luke, S.H.; Swinfield, T.; Chung, A.Y.C.; Coomes, D.A.; Heroin, H.; Jucker, T.; Lewis, O.T.; Vairappan, C.S.; et al. Riparian Buffers Act as Microclimatic Refugia in Oil Palm Landscapes. J. Appl. Ecol. 2020, 58, 431–442. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; van den Hoogen, J.; Aalto, J.; Ashcroft, M.B.; de Frenne, P.D.; Kemppinen, J.; Kopecký, M.; Luoto, M.; Maclean, I.M.D.; Crowther, T.W.; et al. Global Maps of Soil Temperature. Glob. Chang. Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, F.; de Frenne, P.D.; Lenoir, J.; Rocchini, D.; Coomes, D. Advances in Microclimate Ecology Arising from Remote Sensing. Trends Ecol. Evol. 2019, 34, 327–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, M.B.; Domingo, D.; Pizarro, M.; Font, X.; Gómez, D.; Ehrlén, J. Rocky Habitats as Microclimatic Refuges for Biodiversity. A Close-up Thermal Approach. Environ. Exp. Bot. 2020, 170, 103886. [Google Scholar] [CrossRef]
- George, A.D.; Thompson, F.R.; Faaborg, J. Using LiDAR and Remote Microclimate Loggers to Downscale Near-Surface Air Temperatures for Site-Level Studies. Remote Sens. Lett. 2015, 6, 924–932. [Google Scholar] [CrossRef]
- Keppel, G.; Robinson, T.P.; Wardell-Johnson, G.W.; Yates, C.J.; Niel, K.P.V.; Byrne, M.; Schut, A.G.T. A Low-Altitude Mountain Range as an Important Refugium for Two Narrow Endemics in the Southwest Australian Floristic Region Biodiversity Hotspot. Ann. Bot. 2017, 119, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Jucker, T.; Hardwick, S.R.; Both, S.; Elias, D.M.O.; Ewers, R.M.; Milodowski, D.T.; Swinfield, T.; Coomes, D.A. Canopy Structure and Topography Jointly Constrain the Microclimate of Human-Modified Tropical Landscapes. Glob. Chang. Biol. 2018, 24, 5243–5258. [Google Scholar] [CrossRef] [Green Version]
- Stickley, S.F.; Fraterrigo, J.M. Understory Vegetation Contributes to Microclimatic Buffering of Near-Surface Temperatures in Temperate Deciduous Forests. Landsc. Ecol. 2021, 36, 1197–1213. [Google Scholar] [CrossRef]
- Stewart, S.B.; Elith, J.; Fedrigo, M.; Kasel, S.; Roxburgh, S.H.; Bennett, L.T.; Chick, M.; Fairman, T.; Leonard, S.; Kohout, M.; et al. Climate Extreme Variables Generated Using Monthly Time-Series Data Improve Predicted Distributions of Plant Species. Ecography 2021, 44, 626–639. [Google Scholar] [CrossRef]
- Pardo, I.; Roquet, C.; Lavergne, S.; Olesen, J.M.; Gómez, D.; García, M.B. Spatial congruence between taxonomic, phylogenetic and functional hotspots: True pattern or methodological artefact? Divers. Distrib. 2017, 23, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, S.; Sistla, S.; Calheiros, M.D.; Kahraman, A.; Reznicek, A.A.; Rosenberg, R.; von Wettberg, E.J.B. Tracking Microhabitat Temperature Variation with IButton Data Loggers. Appl. Plant Sci. 2019, 7, e01237. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- McGaughey, R. FUSION/LDV: Software for LIDAR Data Analysis and Visualization 2009, Version 3.10; USDA Forest Service: Washington, DC, USA, 2014; pp. 1–212.
- Serrano-Notivoli, R.; Beguería, S.; Saz, M.Á.; Longares, L.A.; de Luis, M. SPREAD: A high-resolution daily gridded precipitation dataset for Spain—An extreme events frequency and intensity overview. Earth Syst. Sci. Data 2017, 9, 721–738. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Notivoli, R.; Beguería, S.; de Luis, M. STEAD: A high-resolution daily gridded temperature dataset for Spain, Earth Syst. Sci. Data 2019, 11, 1171–1188. [Google Scholar] [CrossRef] [Green Version]
- Montealegre, A.L.; Lamelas, M.T.; de la Riva, J. A Comparison of Open-Source LiDAR Filtering Algorithms in a Mediterranean Forest Environment. IEEE J. Sel. Top. Appl. 2015, 8, 4072–4085. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S.; Hudak, A.T. A Multiscale Curvature Algorithm for Classifying Discrete Return LiDAR in Forested Environments. IEEE Trans. Geosci. Remote 2007, 45, 1029–1038. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.1-13. 2020. Available online: https://CRAN.R-project-org/package=raster (accessed on 18 February 2022).
- Boehner, J.; Antonic, O. Land-surface parameters specific to topo-climatology. In Geomorphometry-Concepts, Software, Applications; Hengl, T., Reuter, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; Volume 33, pp. 195–226. [Google Scholar]
- Jennings, S.; Brown, N.; Sheil, D. Assessing Forest Canopies and Understorey Illumination: Canopy Closure, Canopy Cover and Other Measures. Int. J. Res. 1999, 72, 59–74. [Google Scholar] [CrossRef]
- Fox, J.F.; Wesberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.46.0. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 25 February 2022).
- Grömping, U. Relative Importance for Linear Regression in R : The Package Relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Gunton, R.M.; Polce, C.; Kunin, W.E. Predicting Ground Temperatures across European Landscapes. Methods Ecol. Evol. 2015, 6, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Ashcroft, M.B.; Gollan, J.R. Fine-Resolution (25 m) Topoclimatic Grids of near-Surface (5 Cm) Extreme Temperatures and Humidities across Various Habitats in a Large (200 × 300 Km) and Diverse Region. Int. J. Climatol. 2012, 90, 2134–2148. [Google Scholar] [CrossRef] [Green Version]
- Meineri, E.; Hylander, K. Fine-Grain, Large-Domain Climate Models Based on Climate Station and Comprehensive Topographic Information Improve Microrefugia Detection. Ecography 2017, 40, 1003–1013. [Google Scholar] [CrossRef]
- Gubler, M.; Henne, P.D.; Schwörer, C.; Boltshauser-Kaltenrieder, P.; Lotter, A.F.; Brönnimann, S.; Tinner, W. Microclimatic Gradients Provide Evidence for a Glacial Refugium for Temperate Trees in a Sheltered Hilly Landscape of Northern Italy. J. Biogeogr. 2018, 45, 2564–2575. [Google Scholar] [CrossRef]
- Frey, S.J.K.; Hadley, A.S.; Johnson, S.L.; Schulze, M.; Jones, J.A.; Betts, M.G. Spatial Models Reveal the Microclimatic Buffering Capacity of Old-Growth Forests. Sci. Adv. 2016, 2, e1501392. [Google Scholar] [CrossRef] [Green Version]
- Dobrowski, S.Z.; Abatzoglou, J.T.; Greenberg, J.A.; Schladow, S.G. How Much Influence Does Landscape-Scale Physiography Have on Air Temperature in a Mountain Environment? Agric. For. Meteorol. 2009, 149, 1751–1758. [Google Scholar] [CrossRef]
- Aalto, J.; Riihimäki, H.; Meineri, E.; Hylander, K.; Luoto, M. Revealing Topoclimatic Heterogeneity Using Meteorological Station Data. Int. J. Climatol. 2017, 37, 544–556. [Google Scholar] [CrossRef]
- de Frenne, P.D.; Lenoir, J.; Luoto, M.; Scheffers, B.R.; Zellweger, F.; Aalto, J.; Ashcroft, M.B.; Christiansen, D.M.; Decocq, G.; Pauw, K.D.; et al. Forest Microclimates and Climate Change: Importance, Drivers and Future Research Agenda. Glob. Chang. Biol. 2021, 27, 2279–2297. [Google Scholar] [CrossRef]
- McCullough, I.M.; Davis, F.W.; Dingman, J.R.; Flint, L.E.; Flint, A.L.; Serra-Diaz, J.M.; Syphard, A.D.; Moritz, M.A.; Hannah, L.; Franklin, J. High and Dry: High Elevations Disproportionately Exposed to Regional Climate Change in Mediterranean-Climate Landscapes. Landsc. Ecol. 2015, 31, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Reside, A.E.; Welbergen, J.A.; Phillips, B.L.; Wardell-Johnson, G.W.; Keppel, G.; Ferrier, S.; Williams, S.E.; Van Der Wal, J. Characteristics of Climate Change Refugia for Australian Biodiversity. Austral Ecol. 2014, 39, 887–897. [Google Scholar] [CrossRef]
- Schut, A.G.T.; Wardell-Johnson, G.W.; Yates, C.J.; Keppel, G.; Baran, I.; Franklin, S.E.; Hopper, S.D.; Niel, K.P.V.; Mucina, L.; Byrne, M. Rapid Characterisation of Vegetation Structure to Predict Refugia and Climate Change Impacts across a Global Biodiversity Hotspot. PLoS ONE 2014, 9, e82778. [Google Scholar] [CrossRef] [Green Version]
- Greiser, C.; Ehrlén, J.; Meineri, E.; Hylander, K. Hiding from the Climate: Characterizing Microrefugia for Boreal Forest Understory Species. Glob. Chang. Biol. 2019, 26, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zellweger, F.; de Frenne, P.D.; Lenoir, J.; Vangansbeke, P.; Verheyen, K.; Bernhardt-Römermann, M.; Baeten, L.; Hédl, R.; Berki, I.; Brunet, J.; et al. Forest Microclimate Dynamics Drive Plant Responses to Warming. Science 2020, 368, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Lombaerde, E.D.; Vangansbeke, P.; Lenoir, J.; Meerbeek, K.V.; Lembrechts, J.; Rodríguez-Sánchez, F.; Luoto, M.; Scheffers, B.; Haesen, S.; Aalto, J.; et al. Maintaining Forest Cover to Enhance Temperature Buffering under Future Climate Change. Sci. Total Environ. 2021, 810, 151338. [Google Scholar] [CrossRef]
- Norris, C.; Hobson, P.; Ibisch, P.L. Microclimate and Vegetation Function as Indicators of Forest Thermodynamic Efficiency. J. Appl. Ecol. 2012, 49, 562–570. [Google Scholar] [CrossRef]
- Zellweger, F.; Coomes, D.; Lenoir, J.; Depauw, L.; Maes, S.L.; Wulf, M.; Kirby, K.J.; Brunet, J.; Kopecký, M.; Máliš, F.; et al. Seasonal Drivers of Understorey Temperature Buffering in Temperate Deciduous Forests across Europe. Glob. Ecol. Biogeogr. 2019, 28, 1774–1786. [Google Scholar] [CrossRef] [Green Version]
- Oliver, T.; Roy, D.B.; Hill, J.K.; Brereton, T.; Thomas, C.D. Heterogeneous Landscapes Promote Population Stability. Ecol. Lett. 2010, 13, 473–484. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental Heterogeneity as a Universal Driver of Species Richness across Taxa, Biomes and Spatial Scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Hylander, K.; Ehrlén, J.; Luoto, M.; Meineri, E. Microrefugia: Not for Everyone. AMBIO 2015, 44, 60–68. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metric | Description |
|---|---|
| Winter | Mean of December, January, and February |
| Spring | Mean of March, April, and May |
| Summer | Mean of June, July, and August |
| Autumn | Mean of September, October, and November |
| Mean Annual | Mean of the annual monthly values |
| Extreme Annual | Highest and lowest annual monthly values |
| Variable Name | Type | Min.–Max. Values | Description of Min./Max. Values |
|---|---|---|---|
| Elevation | T | 587–3355 m a.s.l. | Lowest/Highest elevation |
| Slope | T | 0–87.98° | Flat terrain/Maximum steep slope |
| Northness | T | −1–1 | Southward/Northward aspect |
| Eastness | T | −1–1 | Westward/Eastward aspect |
| Flow direction | T | 1–128 | Minimum/Maximum water accumulation |
| Heat load | T | −1–1 | Minimum/Maximum solar exposure |
| TPI | T | −186–254 | Concave/Convex landforms |
| NDVImax | V | 0–1 | No intensity/Maximum intensity of vegetation |
| Canopy closure | V | 0–100% | 0%/100% of sky covered by vegetation from a single ground point |
| Elev. CV | V | 0–7.55 | Minimum/Maximum variability of vegetation heights |
| Strata below 1 m | V | 0–1 | No returns/All returns proportion below 1 m |
| Miroclimatic Metrics | Refugial Capacity Metrics | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stability | Offsets | |||||||||||
| Air Tn | Air Tx | Soil Tn | Soil Tx | VPDmax | Air ST | Soil ST | Air Tn | Air Tx | Soil Tn | Soil Tx | ||
| Winter | R2 | 0.39 | 0.76 | 0.71 | 0.77 | 0.64 | 0.62 | 0.66 | 0.76 | 0.49 | 0.86 | 0.76 |
| RMSE | 1.46 | 2.06 | 0.96 | 1.21 | 0.12 | 2.29 | 0.68 | 1.25 | 2.03 | 0.75 | 1.14 | |
| Spring | R2 | 0.87 | 0.84 | 0.93 | 0.87 | 0.86 | 0.62 | 0.51 | 0.72 | 0.41 | 0.51 | 0.56 |
| RMSE | 0.84 | 2.31 | 0.81 | 1.46 | 0.16 | 2.68 | 1.13 | 0.72 | 2.43 | 0.77 | 1.51 | |
| Summer | R2 | 0.86 | 0.42 | 0.81 | 0.58 | 0.49 | 0.38 | 0.43 | 0.35 | 0.36 | 0.62 | 0.58 |
| RMSE | 1.20 | 3.55 | 1.29 | 2.46 | 0.53 | 3.91 | 2.03 | 1.09 | 3.61 | 1.06 | 2.42 | |
| Autumn | R2 | 0.83 | 0.57 | 0.83 | 0.78 | 0.51 | 0.48 | 0.51 | 0.58 | 0.52 | 0.70 | 0.73 |
| RMSE | 1.13 | 2.70 | 1.12 | 1.42 | 0.31 | 2.89 | 1.19 | 1.00 | 2.70 | 0.80 | 1.36 | |
| Mean Annual | R2 | 0.83 | 0.75 | 0.87 | 0.81 | 0.63 | 0.48 | 0.47 | 0.69 | 0.45 | 0.76 | 0.68 |
| RMSE | 1.01 | 2.05 | 0.92 | 1.37 | 0.23 | 2.43 | 1.13 | 0.87 | 2.19 | 0.68 | 1.40 | |
| Extreme Annual | R2 | 0.34 | 0.40 | 0.68 | 0.58 | 0.39 | 0.43 | 0.52 | 0.40 | 0.41 | 0.73 | 0.61 |
| RMSE | 1.63 | 3.82 | 0.91 | 2.51 | 0.70 | 3.91 | 2.59 | 1.19 | 3.83 | 1.05 | 2.56 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffrén, R.; Miranda, H.; Pizarro, M.; Tejero, P.; García, M.B. Identifying the Factors behind Climate Diversification and Refugial Capacity in Mountain Landscapes: The Key Role of Forests. Remote Sens. 2022, 14, 1708. https://doi.org/10.3390/rs14071708
Hoffrén R, Miranda H, Pizarro M, Tejero P, García MB. Identifying the Factors behind Climate Diversification and Refugial Capacity in Mountain Landscapes: The Key Role of Forests. Remote Sensing. 2022; 14(7):1708. https://doi.org/10.3390/rs14071708
Chicago/Turabian StyleHoffrén, Raúl, Héctor Miranda, Manuel Pizarro, Pablo Tejero, and María B. García. 2022. "Identifying the Factors behind Climate Diversification and Refugial Capacity in Mountain Landscapes: The Key Role of Forests" Remote Sensing 14, no. 7: 1708. https://doi.org/10.3390/rs14071708
APA StyleHoffrén, R., Miranda, H., Pizarro, M., Tejero, P., & García, M. B. (2022). Identifying the Factors behind Climate Diversification and Refugial Capacity in Mountain Landscapes: The Key Role of Forests. Remote Sensing, 14(7), 1708. https://doi.org/10.3390/rs14071708