
Developing the Role of Earth Observation in Spatio-Temporal Mosquito Modelling to Identify Malaria Hot-Spots

 , , ,
, , ,  , and
, and
Abstract
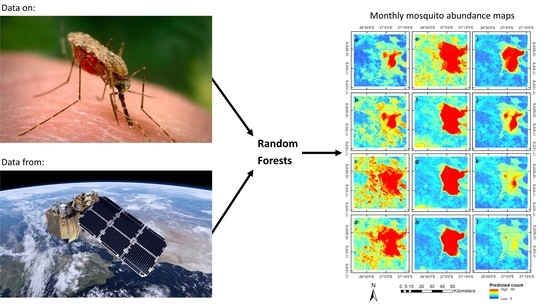
:
1. Introduction
2. Materials and Methods
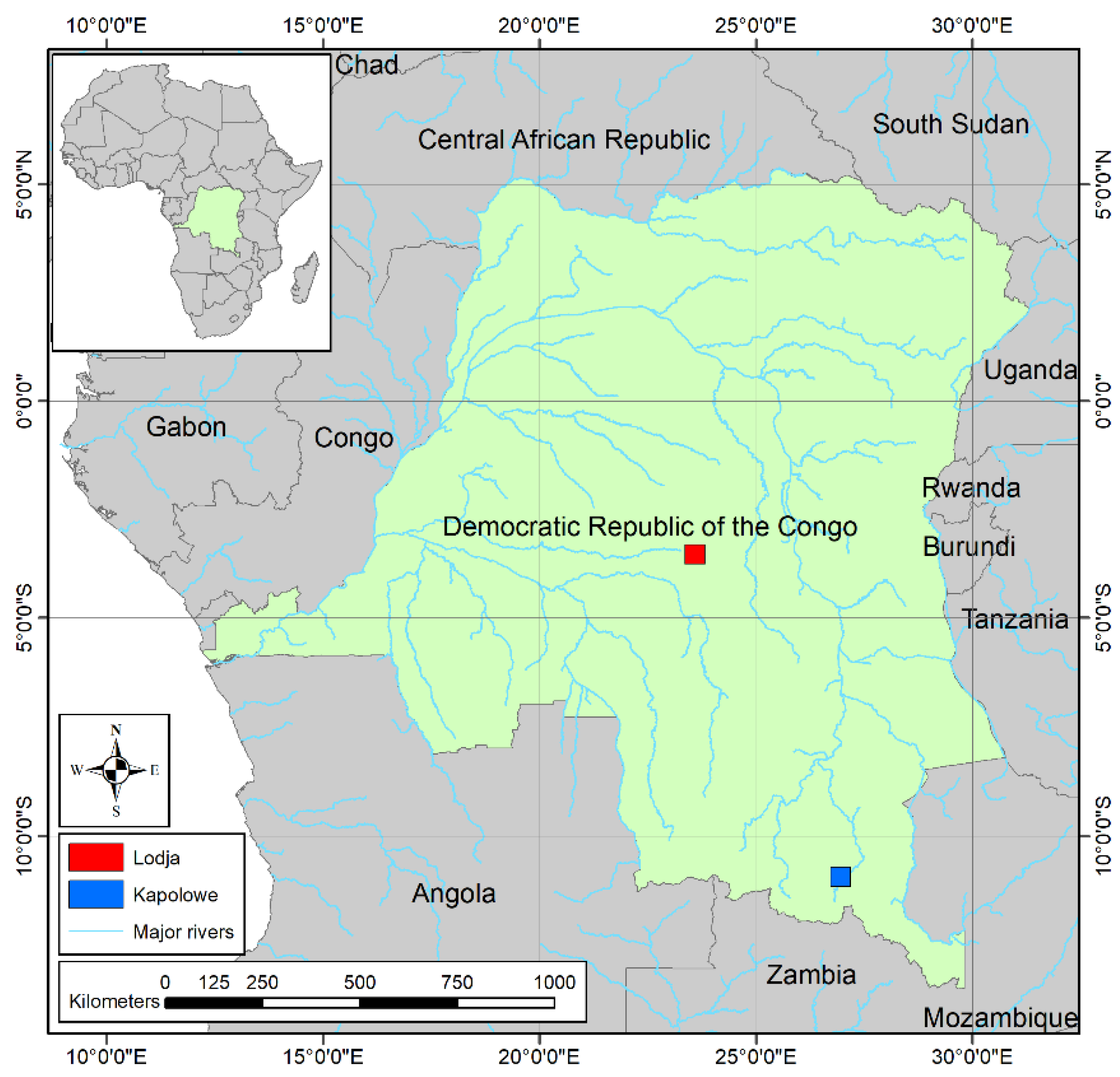
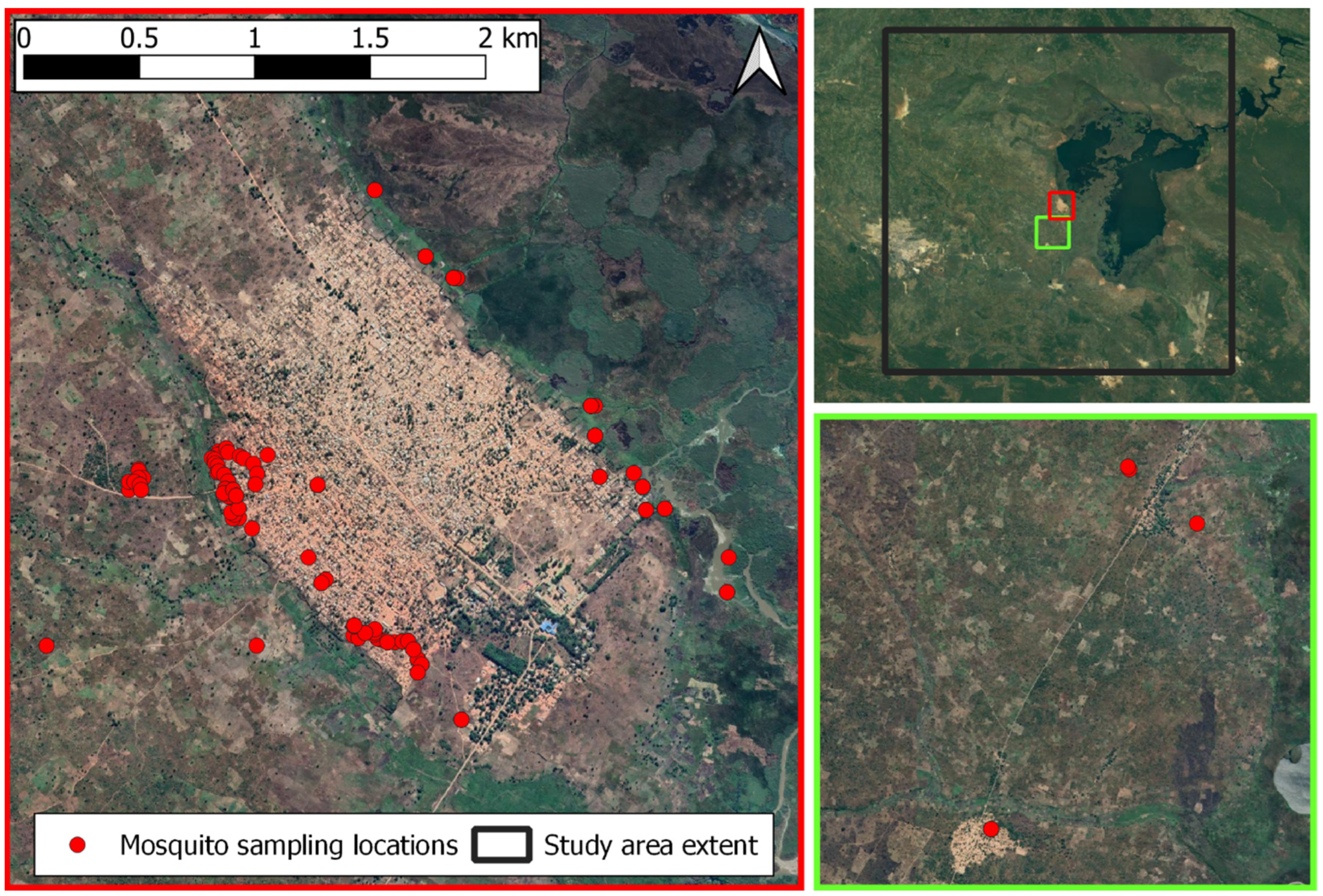
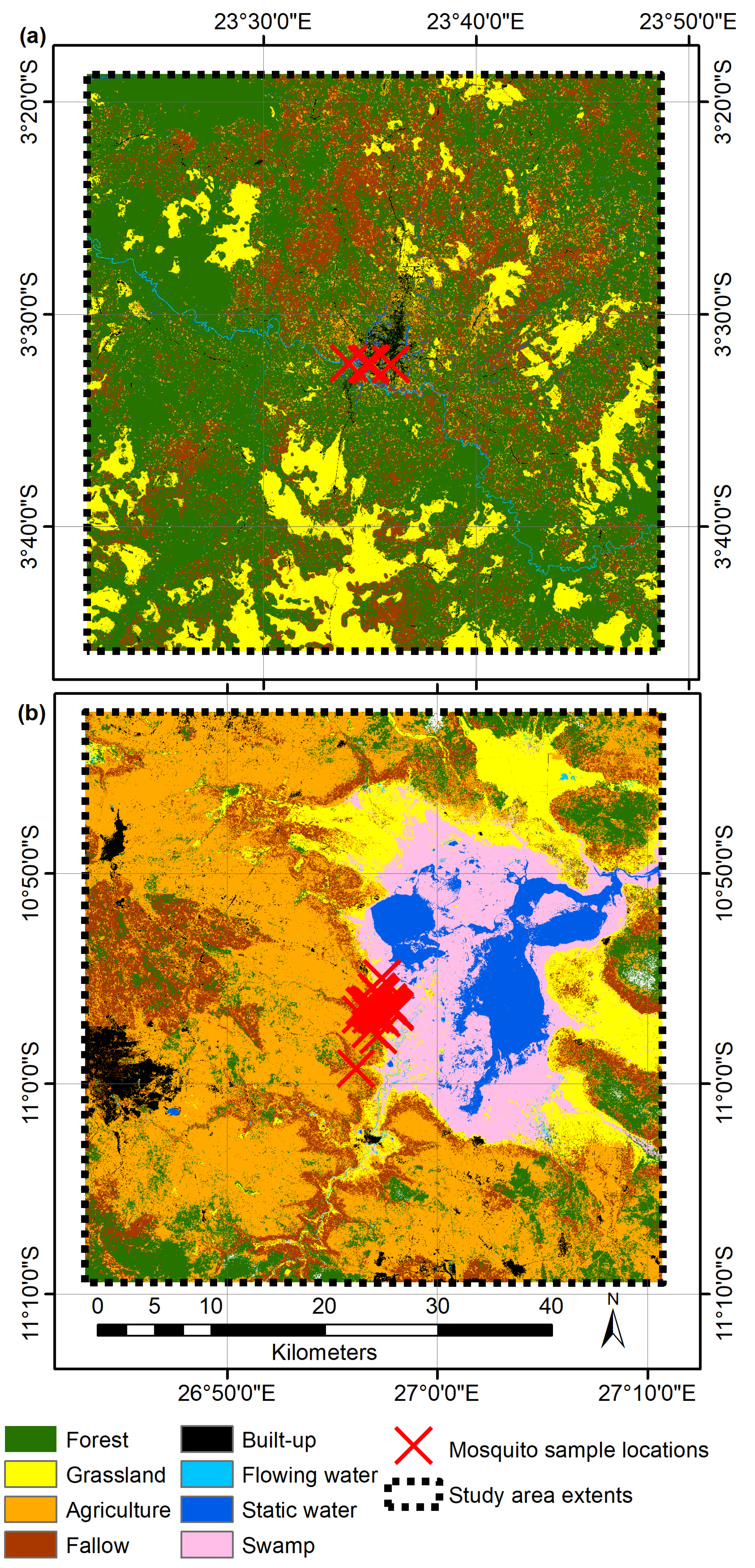
2.1. Study Area
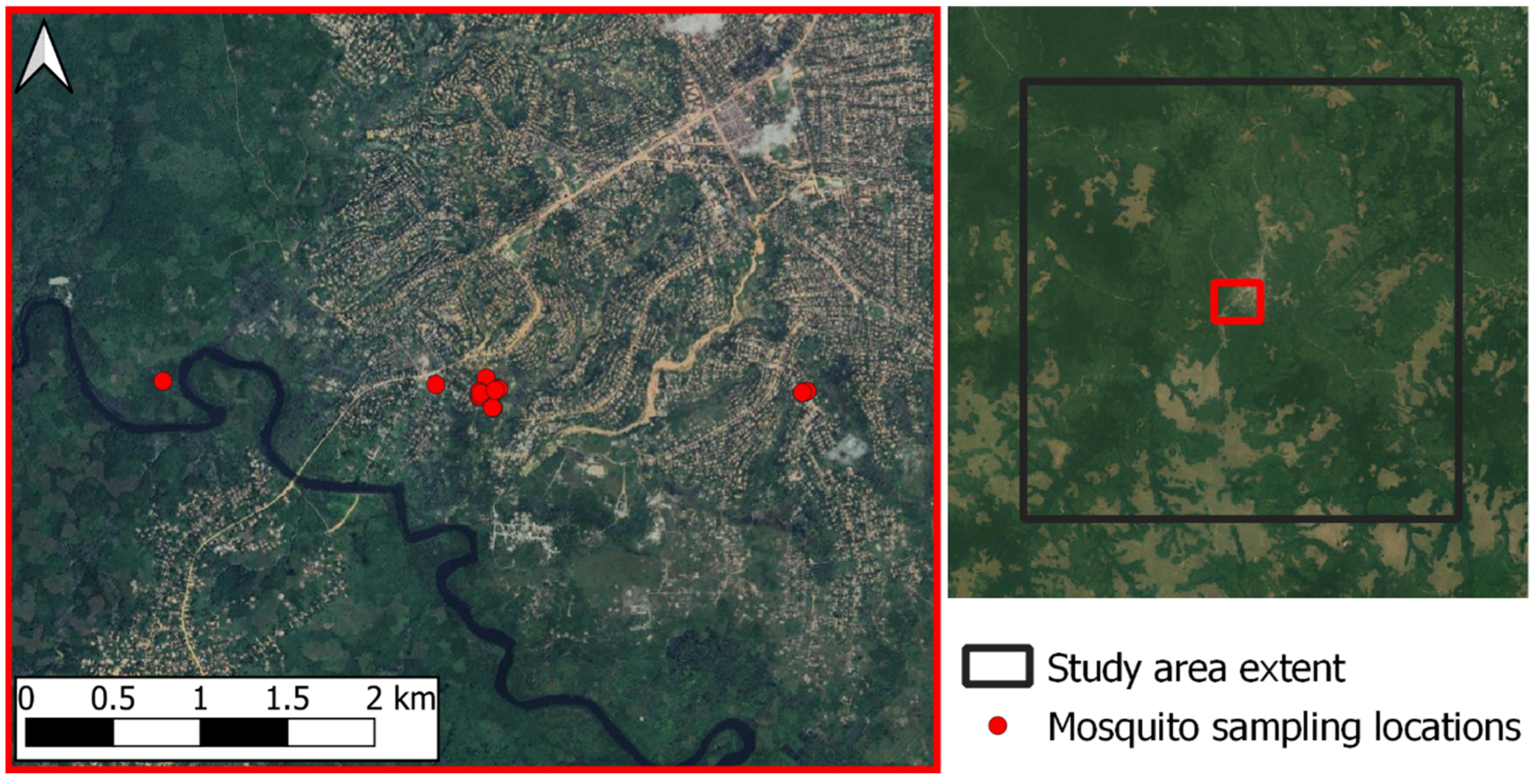
2.2. In Situ Mosquito Data Collection
2.3. Environmental Variables
2.4. Remote Sensing Analysis
2.4.1. Pre-Processing
2.4.2. Land Cover Classification
2.4.3. Vegetation and Water Indices
2.5. Climatic Data
2.6. Random Forest Analysis and Predictive Risk Modelling
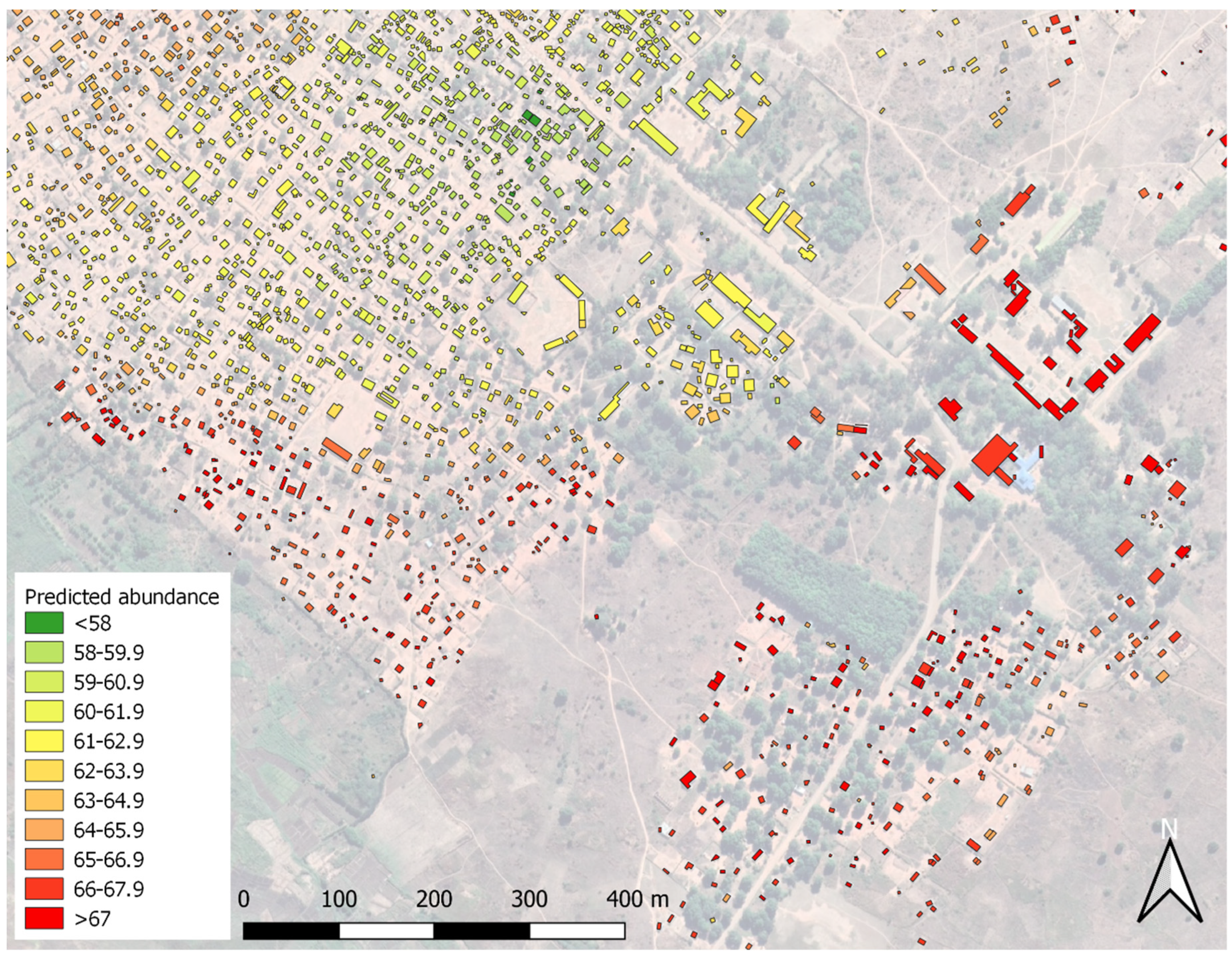
2.7. Open Buildings
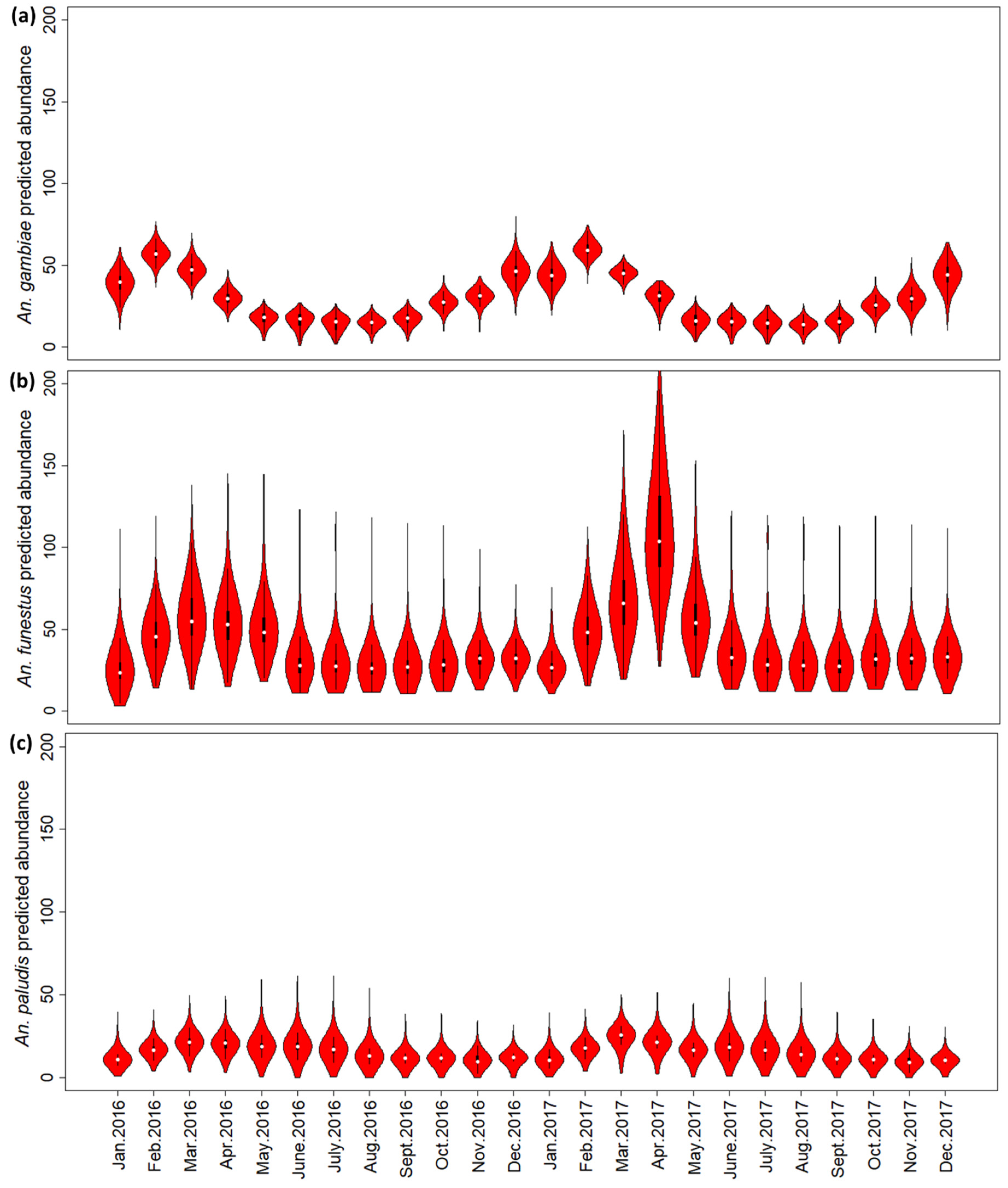
3. Results
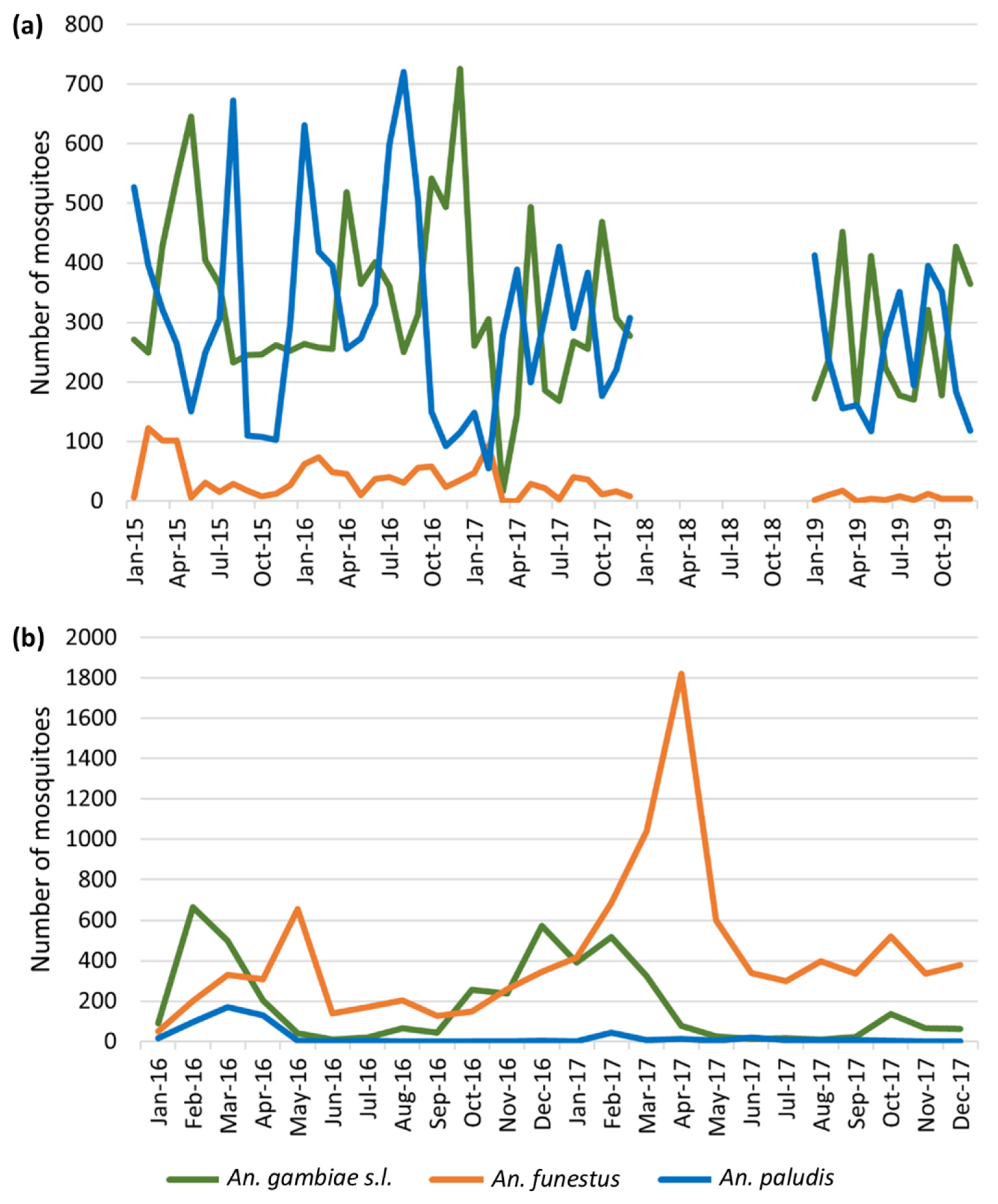
3.1. Mosquito Survey Data
3.2. Land Cover Classification
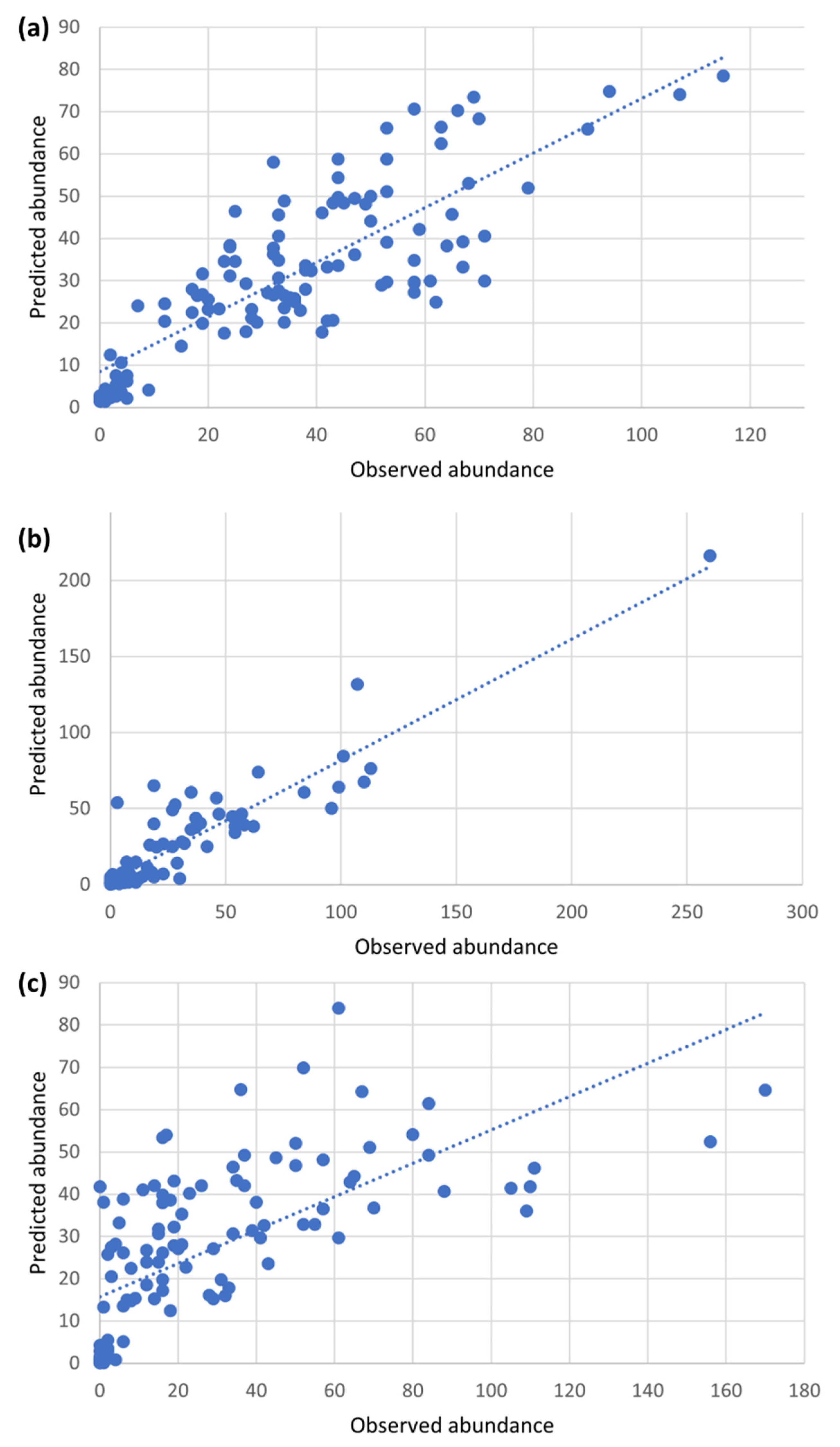
3.3. Random Forest Regression Analysis
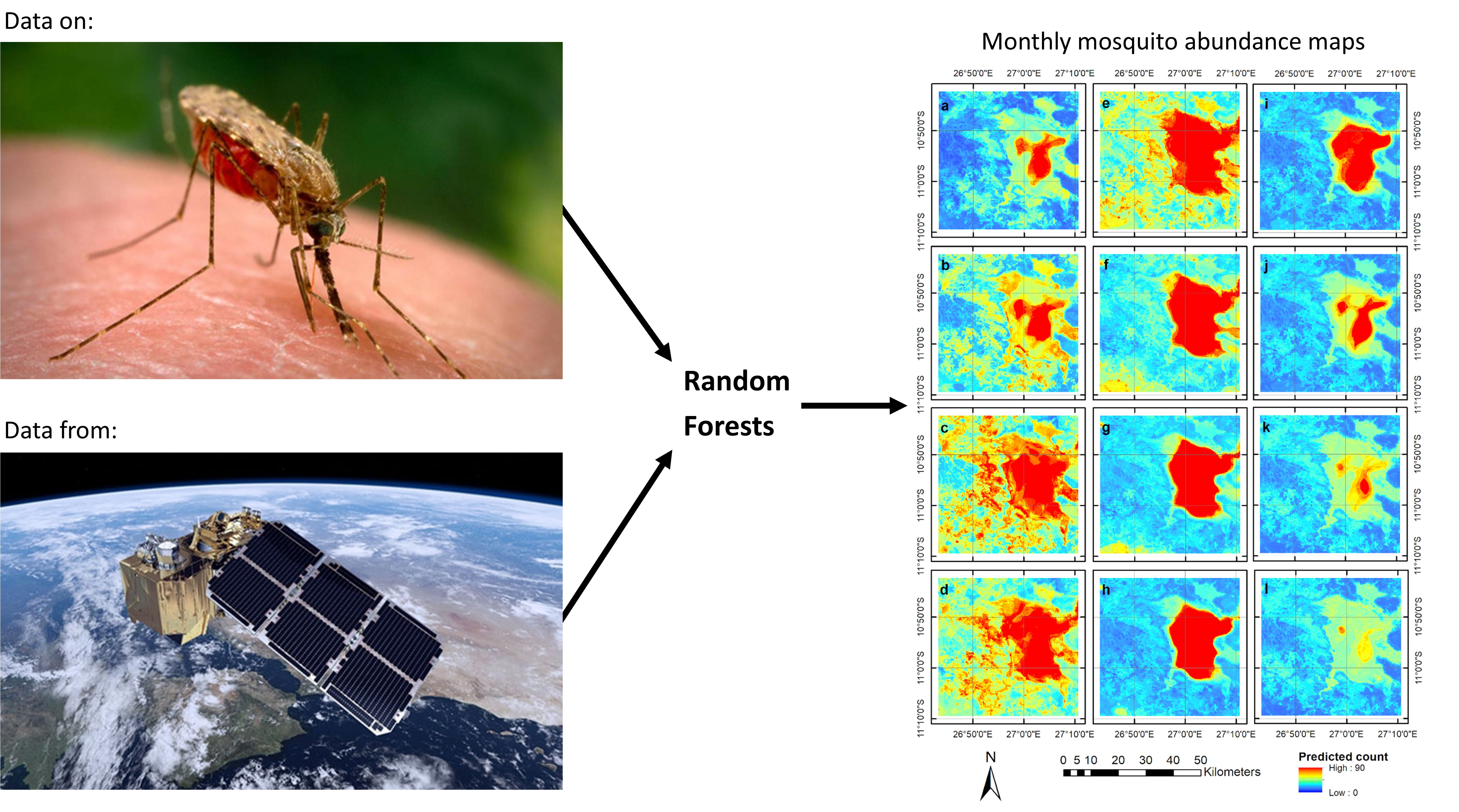
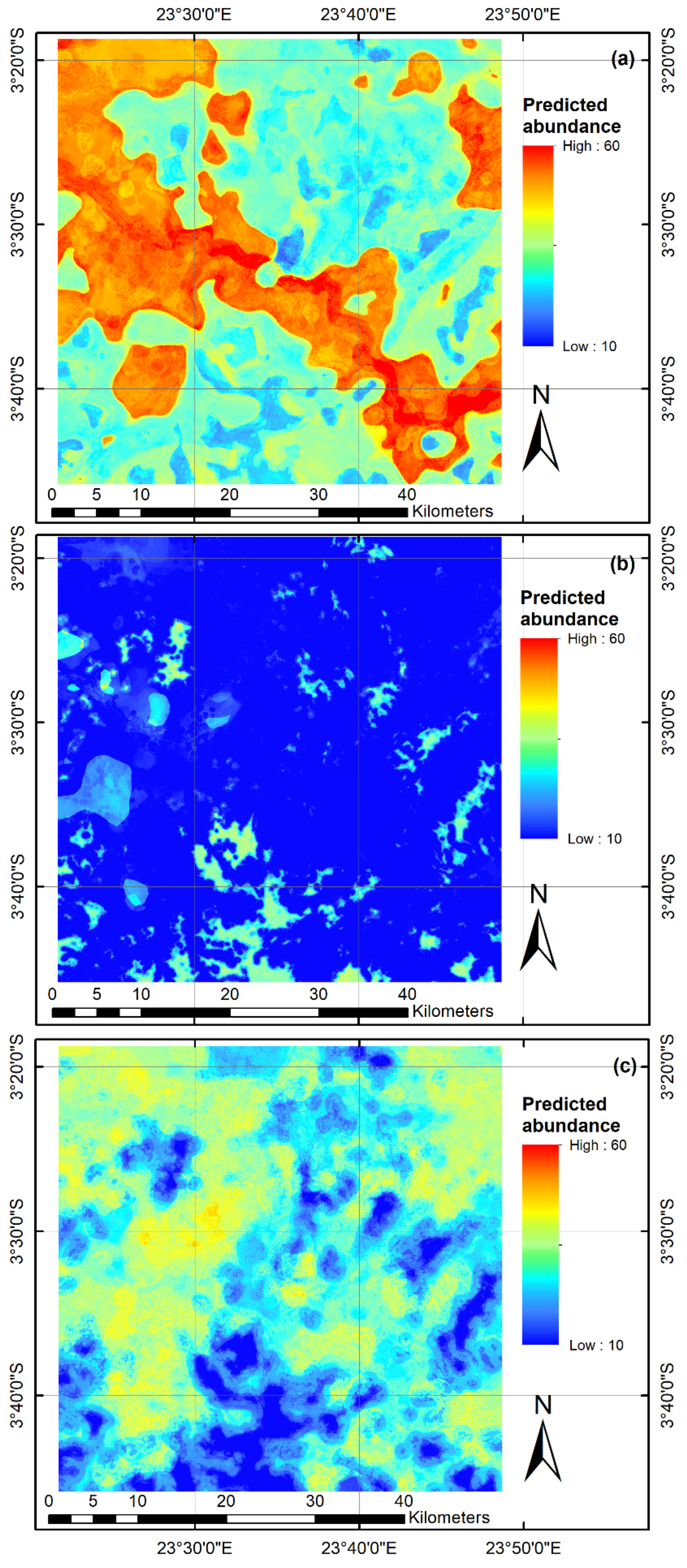
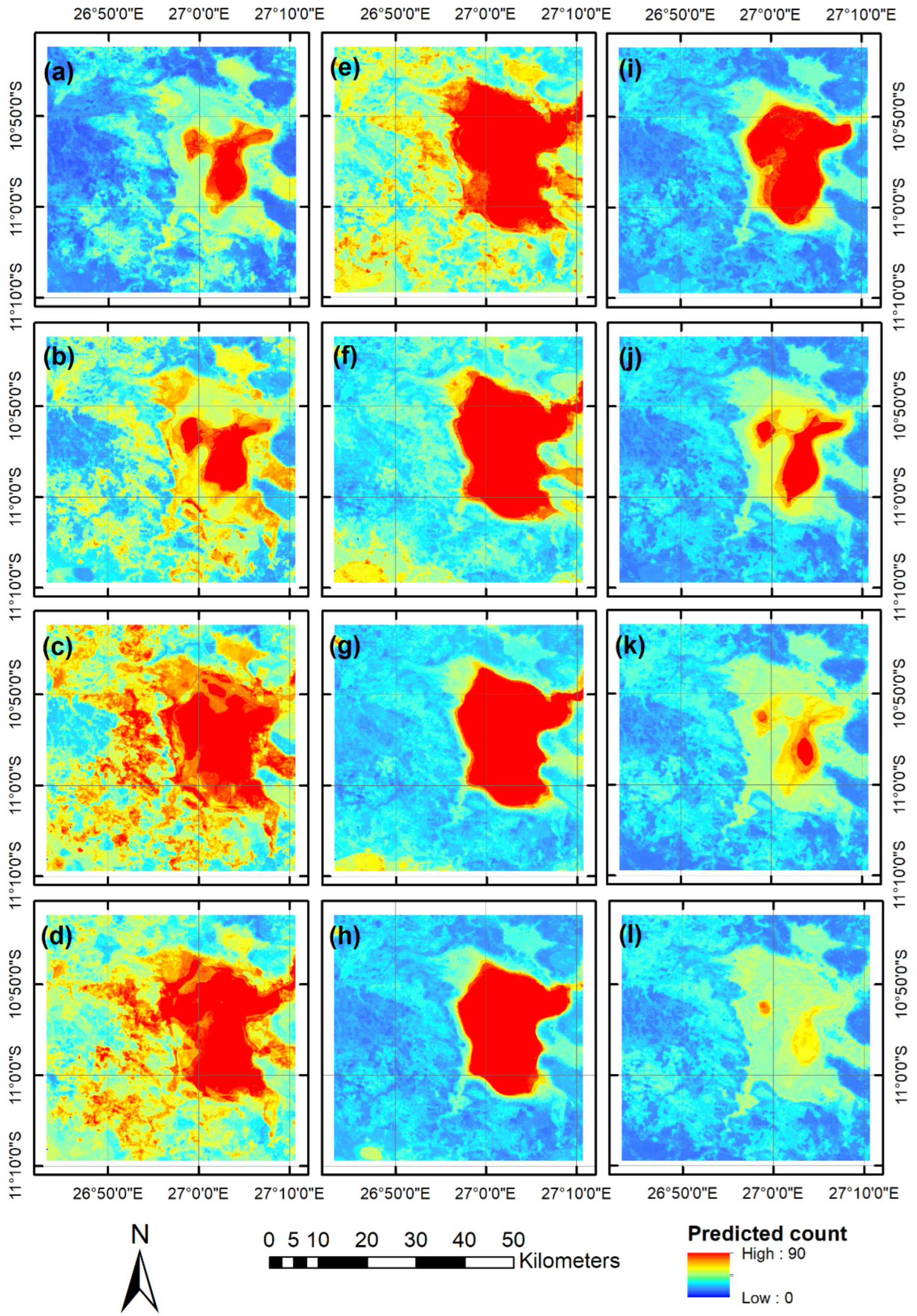
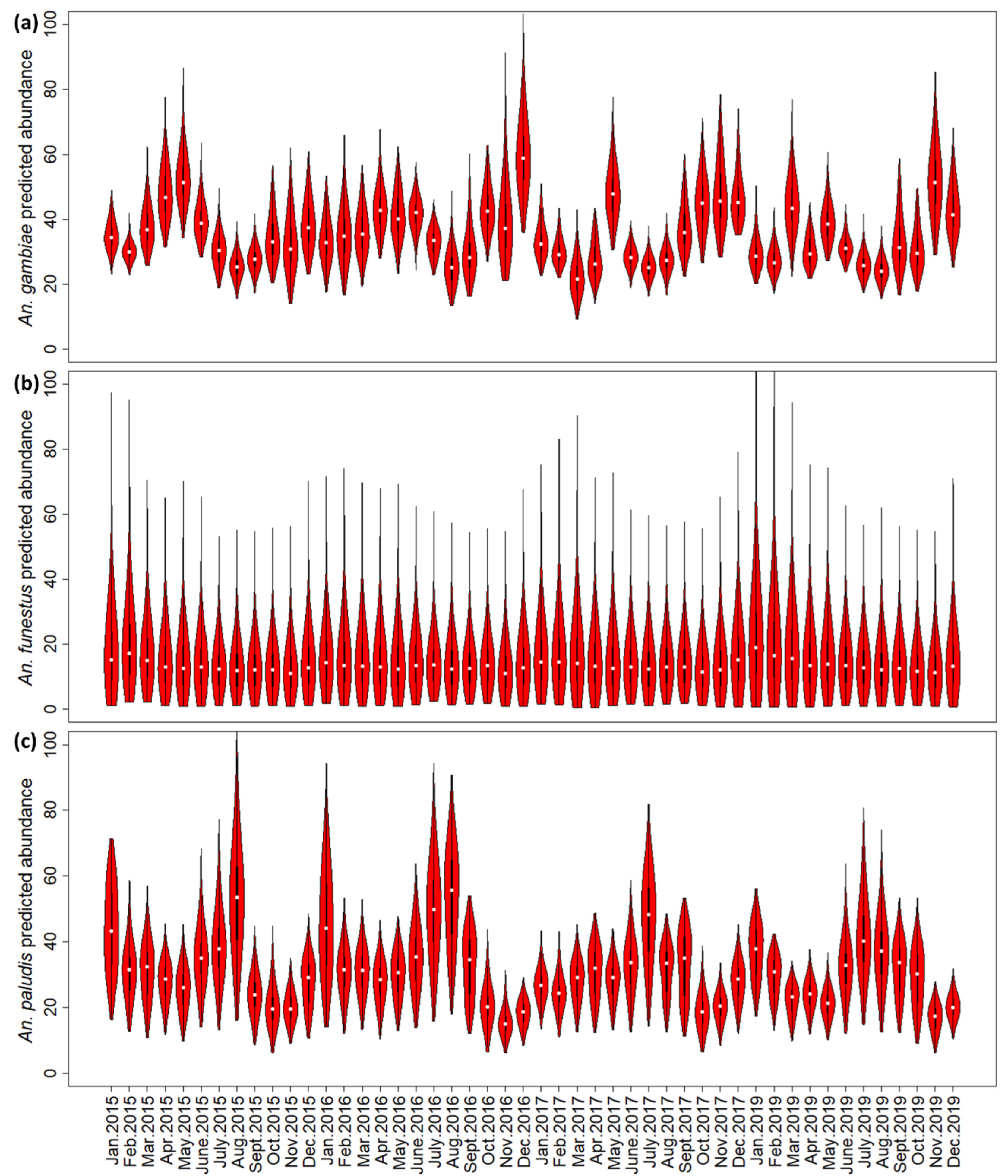
3.4. Predictive Risk Modelling
4. Discussion
4.1. Identifying Spatio-Temporal Drivers of Malaria Risk
4.2. Spatio-Temporal Risk Maps
4.3. Transferability of the Method
4.4. Future Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2021. 2021. Available online: https://www.who.int/publications/i/item/9789240040496 (accessed on 4 February 2022).
- Bhatt, S.; Weiss, D.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. World Malaria Report 2017. 2017. Available online: http://www.who.int/malaria/publications/world-malaria-report-2017/en/ (accessed on 15 March 2021).
- World Health Organization. Global Technical Strategy for Malaria 2016–2030. 2015. Available online: https://www.who.int/malaria/publications/atoz/9789241564991/en/ (accessed on 15 March 2021).
- World Health Organization. Larval Source Management: A Supplementary Measure for Malaria Vector Control: An Operational Manual. 2013. Available online: https://www.who.int/malaria/publications/atoz/9789241505604/en/ (accessed on 15 March 2021).
- Hardy, A.; Mageni, Z.; Dongus, S.; Killeen, G.; Macklin, M.G.; Majambare, S.; Ali, A.; Msellem, M.; Al-Mafazy, A.; Smith, M.; et al. Mapping hotspots of malaria transmission from pre-existing hydrology, geology and geomorphology data in the pre-elimination context of Zanzibar, United Republic of Tanzania. Parasites Vectors 2015, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, A.; Makame, M.; Cross, D.; Majambere, S.; Msellem, M. Using low-cost drones to map malaria vector habitats. Parasites Vectors 2017, 10, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, S.I.; Rogers, D.J.; Toomer, J.F.; Snow, R.W. Annual Plasmodium falciparum entomological inoculation rates (EIR) across Africa: Literature survey, Internet access and review. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 113–127. [Google Scholar] [CrossRef]
- Busula, A.O.; Takken, W.; Loy, D.E.; Hahn, B.H.; Mukabana, W.R.; Verhulst, N.O. Mosquito host preferences affect their response to synthetic and natural odour blends. Malar. J. 2015, 14, 133. [Google Scholar] [CrossRef] [Green Version]
- Fontenille, D.; Lochouarn, L. The complexity of the malaria vectorial system in Africa. Parassitologia 1999, 41, 267–271. [Google Scholar]
- Janko, M.M.; Irish, S.R.; Reich, B.J.; Peterson, M.; Doctor, S.M.; Mwandagalirwa, M.K.; Likwela, J.L.; Tshefu, A.K.; Meshnick, S.R.; Emch, M.E. The links between agriculture, Anopheles mosquitoes, and malaria risk in children younger than 5 years in the Democratic Republic of the Congo: A population-based, cross-sectional, spatial study. Lancet Planet Health 2018, 2, e74–e82. [Google Scholar] [CrossRef] [Green Version]
- Karch, S.; Mouchet, J. Anopheles paludis: Vecteur important du paludisme au Zaïre [Anopheles paludis: Important vector of malaria in Zaire]. Bull. Soc. Pathol. Exot. 1992, 85, 388–389. [Google Scholar]
- Li, Z.; Roux, E.; Dessay, N.; Girod, R.; Steefani, A.; Nacher, M.; Moiret, A.; Seyler, F. Mapping a knowledge-based Malaria hazard index related to landscape using remote sensing: Application to cross-border area between French Guiana and Brazil. Remote Sens. 2016, 8, 319. [Google Scholar] [CrossRef] [Green Version]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic precis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Olson, S.H.; Gangnon, R.; Silveira, G.A.; Patz, J.A. Deforestation and malaria in Mancio Lima county, Brazil. Emerg. Infect. Dis. 2010, 16, 1108–1115. [Google Scholar] [CrossRef]
- Kibret, S.; Lautze, J.; McCartney, M.; Wilson, G.G.; Nhamo, L. Malaria impact of large dams in sub-Saharan Africa: Maps, estimates and predictions. Malar. J. 2015, 14, 339. [Google Scholar] [CrossRef]
- Tonnang, H.E.; Kangalawe, R.Y.; Yanda, P.Z. Predicting and mapping malaria under climate change scenarios: The potential redistribution of malaria vectors in Africa. Malar. J. 2010, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Gleiser, R.M. Geoprocessing and expected distribution of diseases (including deforestation, global warming, and other changes). In Arthropod Borne Diseases; Marcones, C.B., Ed.; Springer: Cham, Switzerland, 2017; pp. 577–604. [Google Scholar] [CrossRef]
- Bousema, T.; Griffin, J.T.; Sauerwein, R.W.; Smith, D.L.; Churcher, T.S.; Takken, W.; Ghani, A.; Drakeley, C.; Gosling, R. Hitting hot-spots: Spatial targeting of malaria for control and elimination. PLoS Med. 2012, 9, e1001165. [Google Scholar] [CrossRef] [Green Version]
- Hamm, N.A.S.; Magalhães, R.J.S.; Clements, A.C.A. Earth Observation, Spatial Data Quality, and Neglected Tropical Diseases. PLoS Negl. Trop. Dis. 2015, 9, e0004164. [Google Scholar] [CrossRef]
- Ferrao, J.L.; Niquisse, S.; Mendes, J.M.; Painho, M. Mapping and modelling malaria risk areas using climate, socio-demographic and clinical variables in Chimoio, Mozambique. Int. J. Environ. Res. Public Health. 2018, 15, 795. [Google Scholar] [CrossRef] [Green Version]
- United Nations. Department of Economic and Social Affairs, Population Division. World Urbanization Prospects: The 2014 Revision. Highlights (ST/ESA/SER.A/352); United Nations: New York, NY, USA, 2015; Available online: https://population.un.org/wup/publications/files/wup2014-report.pdf (accessed on 3 May 2022).
- Robert, V.; Macintyre, K.; Keating, J.; Trape, J.F.; Duchemin, J.B.; Warren, M.; Beier, J.C. Malaria transmission in urban sub-Saharan Africa. Am. J. Trop. Med. Hyg. 2003, 68, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Sinka, M.E.; Pironon, S.; Massey, N.C.; Longbottom, J.; Hemingway, J.; Moyes, C.L.; Willis, K.J. A new malaria vector in Africa: Predicting the expansion range of Anopheles stephensi and identifying the urban populations at risk. Proc. Natl. Acad. Sci. USA 2020, 117, 24900–24908. [Google Scholar] [CrossRef]
- Kaindoa, E.W.; Mkandawile, G.B.; Lingamba, G.F.; Killeen, G.F.; Okumu, F.O. Longitudinal surveillance of disease-transmitting mosquitoes in rural Tanzania: Creating an entomological framework for evaluation. Lancet 2013, 381, S70. [Google Scholar] [CrossRef]
- Imbahale, S.S.; Paaijmans, K.P.; Mukabana, W.R.; van Lammeren, R.; Githeko, A.K.; Takken, W. A longitudinal study on Anopheles mosquito larval abundance in distinct geographical and environmental settings in western Kenya. Malar. J. 2011, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Olson, S.H.; Gangnon, R.; Elguero, E.; Durieux, L.; Guegan, J.F.; Foley, J.A.; Patz, J.A. Links between climate, malaria, and wetlands in the Amazon Basin. Emerg. Infect. Dis. 2009, 15, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Baak-Baak, C.M.; Moo-Llanes, D.A.; Cigarroa-Toledo, N.; Puerto, F.I.; Machain-Williams, C.; Reyes-Solis, G.; Nakazawa, Y.J.; Ulloa-Garcia, A.; Garcia-Rejon, J.E. Ecological Niche Model for Predicting Distribution of Disease-Vector Mosquitoes in Yucatán State, México. J. Med. Entomol. 2017, 54, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, N.B.; Caminade, C.; Beierkuhnlein, C.; Thomas, S.M. Mosquito-borne diseases: Advances in modelling climate-change impacts. Trends Parasitol. 2018, 34, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Marston, C.G.; Danson, F.M.; Armitage, R.P.; Giraudoux, P.; Pleydell, D.R.J.; Wang, Q.; Qui, J.; Craig, P.S. A random forest approach for predicting the presence of Echinococcus multilocularis intermediate host Ochotona spp. presence in relation to landscape characteristics in western China. Appl. Geogr. 2014, 55, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danson, F.M.; Armitage, R.P.; Marston, C.G. Spatial and temporal modelling for parasite transmission studies and risk assessment. Parasite 2008, 15, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Midekisa, A.; Senay, G.B.; Wimberly, M.C. Multi-sensor earth observations to characterize wetlands and malaria epidemiology in Ethiopia. Water Resour. Res. 2014, 50, 8791–8806. [Google Scholar] [CrossRef] [Green Version]
- Marston, C.G.; Giraudoux, P.; Armitage, R.P.; Danson, F.M.; Reynolds, S.; Wang, Q.; Qiu, J.; Craig, P.S. Vegetation phenology and habitat discrimination: Impacts for E. multilocularis transmission host modelling. Remote Sens. Environ. 2016, 176, 320–327. [Google Scholar] [CrossRef]
- Wimberly, M.C.; de Beurs, K.M.; Loboda, T.V.; Pan, W.K. Satellite Observations and Malaria: New Opportunities for Research and Applications. Trends Parasitol. 2021, 37, 525–537. [Google Scholar] [CrossRef]
- Adeola, A.M.; Botai, J.O.; Olwoch, J.M.; Rautenbach, H.C.J.W.; Kalumba, A.M.; Tsela, P.l.; Adisa, M.O.; Wasswa, N.F.; Mmtoni, P.; Ssentongo, A. Application of geographical information system and remote sensing in malaria research and control in South Africa: A review. S. Afr. J. Infect. Dis. 2015, 30, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Manica, M.; Filipponi, F.; D’Alessandro, A.; Screti, A.; Neteler, M.; Rosa, R.; Solimini, A.; della Torre, A.; Caputo, B. Spatial and Temporal Hot Spots of Aedes albopictus Abundance inside and outside a South European Metropolitan Area. PLoS Negl. Trop. Dis. 2016, 10, e0004758. [Google Scholar] [CrossRef] [Green Version]
- Moss, W.J.; Hamapumbu, H.; Kobayashi, T.; Shields, T.; Kamanga, A.; Clennon, J.; Mharakurwa, S.; Thuma, P.E.; Glass, G. Use of remote sensing to identify spatial risk factors for malaria in a region of declining transmission: A cross-sectional and longitudinal community survey. Malar. J. 2011, 10, 163. [Google Scholar] [CrossRef]
- Kabaria, C.W.; Molteni, F.; Mandike, R.; Chacky, F.; Noor, A.M.; Snow, R.W.; Linard, C. Mapping intra-urban malaria risk using high resolution satellite imagery: A case study of Dar es Salaam. Int. J. Health Geogr. 2016, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Clerici, N.; Calderón, C.A.V.; Posada, J.M. Fusion of Sentinel-1A and Sentinel-2A data for land cover mapping: A case study in the lower Magdalena region, Colombia. J. Maps. 2017, 13, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Marston, C.G.; Giraudoux, P. On the synergistic use of optical and SAR time-series satellite data for small mammal disease host mapping. Remote Sens. 2019, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.Z.; Qazi, W.; Aslam, N. A comparative study of ALOS-2 PALSAR and landsat-8 imagery for land cover classification using maximum likelihood classifier. Egypt. J. Remote Sens. Space Sci. 2018, 21, S29–S35. [Google Scholar] [CrossRef]
- Torbick, N.; Ledoux, L.; Salas, W.; Zhao, M. Regional mapping of plantation extent using multisensor imagery. Remote Sens. 2016, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Carrasco, L.; O’Neil, A.W.; Morton, R.D.; Rowland, C. Evaluating combinations of temporally aggregated Sentinel-1, Sentinel-2 and Landsat 8 for land cover mapping with Google Earth Engine. Remote Sens. 2019, 11, 288. [Google Scholar] [CrossRef] [Green Version]
- Marston, C.G.; Rowland, C.S.; O’Neil, A.W.; Irish, S.; Wat’senga, F.; Martin-Gallego, P.; Strode, C. Earth Observation for Malaria Modelling: A Practical Toolkit for Satellite-Based Prediction of Mosquito Distributions Using Google Earth Engine and R; UK Centre for Ecology and Hydrology: Lancaster, UK, 2022; 68p. [Google Scholar]
- President’s Malaria Initiative. FY 2018 Democratic Republic of the Congo Malaria Operational Plan. 2018. Available online: https://www.pmi.gov/where-we-work/democratic-republic-of-the-congo (accessed on 15 March 2021).
- Wat’senga, F.; Manzambi, E.Z.; Lunkula, A.; Mulumbu, R.; Mampangulu, T.; Lobo, N.; Hendershot, A.; Fornadel, C.; Jacob, D.; Niang, M.; et al. Nationwide insecticide resistance status and biting behaviour of malaria vector species in the Democratic Republic of Congo. Malar. J. 2018, 17, 129. [Google Scholar] [CrossRef]
- Coetzee, M.; Hunt, R.H.; Wilkerson, R.; della Torre, A.; Coulibaly, M.B.; Besansky, N.J. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa 2013, 3619, 246–274. [Google Scholar] [CrossRef] [Green Version]
- Hardy, A.J.; Gamarra, J.G.P.; Cross, D.E.; Macklin, M.G.; Smith, M.W.; Kihonda, J.; Killeenm, G.F.; Ling’ala, G.N.; Thomas, C.J. Habitat Hydrology and Geomorphology Control the Distribution of Malaria Vector Larvae in Rural Africa. PLoS ONE 2013, 8, e81931. [Google Scholar] [CrossRef] [PubMed]
- National Aeronautics and Space Administration. National Aeronautics and Space Administration (2021): MOD11A1-MODIS/Terra Land Surface Temperature/Emissivity Daily L3 Global 1 km SIN Grid. Available online: https://catalogue.ceda.ac.uk/uuid/35bb28eafbaa461db578e1218808c038 (accessed on 10 November 2022).
- Hay, S.I.; Snow, R.W.; Rogers, D.J. Predicting malaria seasons in Kenya using multi-temporal meteorological satellite sensor data. Trans. R. Soc. Trop. Med. Hyg. 1998, 92, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Snoeij, P.; Geudtner, D.; Bibby, D.; Davidson, M.; Attema, E.; Potin, P.; Rommen, B.; Floury, N.; Brown, M.; et al. GMES Sentinel-1 mission. Remote Sens. Environ. 2012, 120, 9–24. [Google Scholar] [CrossRef]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P.; et al. Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The climate hazards infrared precipitation with stations—a new environmental record for monitoring extremes. Sci. Data 2015, 2, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Hook, S.; Hulley, G. MODIS/Terra Land Surface Temperature/Emissivity Daily L3 Global 1 km SIN Grid V061. 2021, Distributed by NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/MODIS/MOD11A1.061 (accessed on 10 November 2022).
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-Filled SRTM for the Globe Version 4, Available from the CGIAR-CSI SRTM 90 m Database. 2008. Available online: https://srtm.csi.cgiar.org (accessed on 1 March 2021).
- Filipponi, F. Sentinel-1 GRD Preprocessing Workflow. Proceedings 2019, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Molinario, G.; Hansen, M.C.; Potapov, P.V.; Tyukavina, A.; Stehman, S.; Barker, B.; Humber, M. Quantification of land cover and land use within the rural complex of the Democratic Republic of Congo. Environ. Res. Lett. 2017, 12, 104001. [Google Scholar] [CrossRef] [Green Version]
- Duro, D.C.; Franklin, S.E.; Dube, M.G. Multi-scale object-based image analysis and feature selection of multi-sensor earth observation imagery using random forests. Int. J. Remote Sens. 2012, 33, 4502–4526. [Google Scholar] [CrossRef]
- Verdonschot, P.F.; Besse-Lototskaya, A.A. Flight distance of mosquitoes (Culicidae): A metadata analysis to support the management of barrier zones around rewetted and newly constructed wetlands. Limnologica 2014, 45, 69–79. [Google Scholar] [CrossRef]
- Kaufmann, C.; Briegel, H. Flight performance of the malaria vectors Anopheles gambiae and Anopheles atroparvus. J. Vector Ecol. 2004, 29, 140–153. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Deering, D.W.; Schell, J.A.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; E73-10693; NASA: Greenbelt, MD, USA, 1973; p. 112. [Google Scholar]
- Huete, A.R. A Soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1998, 25, 295–309. [Google Scholar] [CrossRef]
- McFeeters, S.K. The use of normalized difference water index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Ali, Z.; Hussain, I.; Faisal, M.; Elashkar, E.E.; Gani, S.; Shehzad, M.A. Selection of appropriate time scale with Boruta algorithm for regional drought monitoring using multi-scaler drought index. Tellus A Dyn. Meteorol. Oceanogr. 2019, 71, 1. [Google Scholar] [CrossRef] [Green Version]
- Kursa, M.B.; Rudnicki, W.R. Feature Selection with the Boruta Package. J. Stat. Softw. 2010, 36, 1–13. Available online: http://www.jstatsoft.org/v36/i11/ (accessed on 6 March 2021). [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Kurza, M.B. Robustness of Random Forest-based gene selection methods. BMC Bioinform. 2014, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Tran, M.; Siwabessy, J. Selecting Optimal Random Forest Predictive Models: A Case Study on Predicting the Spatial Distribution of Seabed Hardness. PLoS ONE 2016, 11, e0149089. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 April 2022).
- Sirko, W.; Kashubin, S.; Ritter, M.; Annkah, A.; Bouchareb, Y.S.E.; Dauphin, Y.; Keysers, D.; Neumann, M.; Cisse, M.; Quinn, J.A. Continental-scale building detection from high resolution satellite imagery. arXiv 2021, arXiv:2107.12283. [Google Scholar]
- Gillies, M.T.; de Meillon, B. The Anophelinae of Africa south of the Sahara (Ethiopian Zoogeographical Region). Publ. S. Afr. Inst. Med. Res. 1968, 54, 1–343. [Google Scholar]
- Tene Fossog, B.; Ayala, D.; Acevedo, P.; Kengne, P.; Ngomo Abeso Mebuy, I.; Makanga, B.; Magnus, J.; Awono-Ambene, P.; Njiokou, F.; Pombi, M.; et al. Habitat segregation and ecological character displacement in cryptic African malaria mosquitoes. Evol. Appl. 2015, 8, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayoh, M.N.; Lindsay, S.W. Temperature-related duration of aquatic stages of the Afrotropical malaria vector mosquito Anopheles gambiae in the laboratory. Med. Vet. Entomol. 2004, 18, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Lyons, C.L.; Coetzee, M.; Chown, S.L. Stable and fluctuating temperature effects on the development rate and survival of two malaria vectors, Anopheles arabiensis and Anopheles funestus. Parasites Vectors 2013, 6, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantur Juri, M.J.; Estallo, E.; Almirón, W.; Santana, M.; Sartor, P.; Lamfri, M.; Zaidenberg, M. Satellite-derived NDVI, LST, and climatic factors driving the distribution and abundance of Anopheles mosquitoes in a former malarious area in northwest Argentina. J. Vector Ecol. 2015, 40, 36–45. [Google Scholar] [CrossRef]
- Shapiro, L.L.M.; Whitehead, S.A.; Thomas, M.B. Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLoS Biol. 2017, 15, e2003489. [Google Scholar] [CrossRef] [Green Version]
- McMahon, A.; Mihretie, A.; Ahmed, A.A.; Lake, M.; Awoke, W.; Wimberley, M.C. Remote sensing of environmental risk factors for malaria in different geographic contexts. Int. J. Health Geogr. 2021, 20, 28. [Google Scholar] [CrossRef]
- Christiansen-Jucht, C.; Parham, P.E.; Saddler, A.; Koella, J.C.; Basáñez, M.G. Temperature during larval development and adult maintenance influences the survival of Anopheles gambiae s.s. Parasites Vectors 2014, 7, 489. [Google Scholar] [CrossRef]
- Gimnig, J.E.; Ombok, M.; Otieno, S.; Kaufman, M.G.; Vulule, J.M.; Walker, E.D. Density-dependent development of Anopheles gambiae (Diptera: Culicidae) larvae in artificial habitats. J. Med. Entomol. 2002, 39, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Yaro, A.S.; Dao, A.; Adamou, A.; Crawford, J.E.; Ribeiro, J.M.; Gwadz, R.; Traoré, S.F.; Lehmann, T. The distribution of hatching time in Anopheles gambiae. Malar. J. 2006, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, F.M.; Manin, B.O.; Cooper, A.; Daim, S.; Homathevi, R.; Jelip, J.; Husin, T.; Chua, T.H. Vector compositions change across forested to deforested ecotones in emerging areas of zoonotic malaria transmission in Malaysia. Sci. Rep. 2019, 9, 13312. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. High Burden to High Impact: A Targeted Malaria Response. 2019. Available online: https://www.who.int/malaria/publications/atoz/high-impact-response/en/) (accessed on 15 April 2021).
- Larsen, D.A.; Martin, A.; Pollard, D.; Nielsen, C.F.; Hamainza, B.; Burns, M.; Stevenson, J.; Winters, A. Leveraging risk maps of malaria vector abundance to guide control efforts reduces malaria incidence in Eastern Province, Zambia. Sci. Rep. 2020, 10, 10307. [Google Scholar] [CrossRef]
- Wanzirah, H.; Tusting, L.S.; Arinaitwe, E.; Katureebe, A.; Maxwell, K.; Rek, J.; Bottomley, C.; Staedke, S.G.; Kamya, M.; Dorsey, G.; et al. Mind the gap: House structure and the risk of malaria in Uganda. PLoS ONE 2015, 10, e0117396. [Google Scholar]
- Hardy, A.; Ettritch, G.; Cross, D.; Bunting, P.; Liywalii, F.; Sakala, J.; Silumesii, A.; Singini, D.; Smith, M.; Willis, T.; et al. Automatic Detection of Open and Vegetated Water Bodies Using Sentinel 1 to Map African Malaria Vector Mosquito Breeding Habitats. Remote Sens. 2019, 11, 593. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Data Source | Reference |
|---|---|---|---|
| Topographic | |||
| Altitude | Elevation values (m). | SRTM DEM. | [21,32,37,49] |
| Slope | Slope values (degrees). | SRTM DEM. | [21,32,37,49] |
| Aspect | Aspect values (degrees). | SRTM DEM. | [21,32,37,49] |
| Topographical Position Index | The elevation of a pixel minus the mean elevation of the surrounding 15-pixel radius area. | SRTM DEM. | [37] |
| Climatic | |||
| Precipitation | Monthly mean precipitation (mean mm/day). | CHIRPS | [21,32,49] |
| Land surface temperature | Monthly mean land surface temperature (Kelvin) | MODIS | [50] |
| Land cover | |||
| Land cover | Proportional coverage of each land cover class, derived from a land cover classification of the study area. The area over which land cover proportions was calculated differed for the different mosquito species based on their established flight distances. Proportional coverage of forest and fallow classes combined was also calculated. | Sentinel-1, Sentinel-2, and SRTM DEM. | [13,20,21,49] |
| Proximity to forest edge | Distance from pixel centroid to the nearest patch of forest, fallow, and forest and fallow classes combined. | Derived from land cover classification. | [37] |
| Proximity to water body | Distance from pixel centroid to the nearest patch of flowing water, static water, and swamp. | Derived from land cover classification. | [6,19] |
| Vegetation Indices | |||
| Normalised Difference Vegetation Index (NDVI), Soil Adjusted Vegetation Index (SAVI), Normalised Difference Water Index (NDWI), and Modified Normalised Difference Water Index (MNDWI). | Median VI values calculated from a collection of cloud-free imagery acquired from the period of mosquito data collection in each study area. | Sentinel-2 | [21,51] |
| Land Cover Class | Description |
|---|---|
| Forest | A forest stand with over 60% tree cover. |
| Grassland | Natural grassland areas. |
| Agriculture | A field where crops are currently grown. |
| Fallow | A field left fallow, often grassy with increasing shrub encroachment. |
| Built-up | Roads, paths, settlements, communal areas along roads, buildings, and huts. |
| Flowing water | Rivers and streams. |
| Static water | Ponds, lakes, and rice paddies, without emergent vegetation. |
| Swamp | Swamp with surface vegetation. |
| Site | Year | An. gambiae s.l. | An. funestus | An. paludis | Total |
|---|---|---|---|---|---|
| Lodja | 2015 | 4143 (51.0%) | 477 (5.9%) | 3503 (43.1%) | 8123 |
| 2016 | 4748 (48.7%) | 520 (5.3%) | 4486 (46.0%) | 9754 | |
| 2017 | 3153 (47.5%) | 305 (4.6%) | 3186 (47.9%) | 6644 | |
| 2019 | 3295 (52.1%) | 69 (1.1%) | 2955 (46.8%) | 6319 | |
| All years | 15339 (49.7%) | 1371 (4.5%) | 14130 (45.8%) | 30840 | |
| Kapolowe | 2016 | 2702 (44.6%) | 2936 (48.4%) | 423 (7.0%) | 6061 |
| 2017 | 1656 (18.5%) | 7171 (80.2%) | 116 (1.3%) | 8943 | |
| All years | 4358 (29.0%) | 10107 (67.4%) | 539 (3.6%) | 15004 |
| Land Cover Class | Lodja | Kapolowe |
|---|---|---|
| Forest | 61.67% (1554.92 km2) | 12.53% (320.40 km) |
| Fallow | 16.48% (415.64 km2) | 13.93% (355.98 km2) |
| Grassland | 12.80% (322.71 km2) | 11.30% (288.93 km2) |
| Agriculture | 7.00% (176.46 km2) | 41.59% (1063.07 km2) |
| Built-up | 0.99% (25.05 km2) | 3.15% (80.51 km2) |
| Swamp | 0.00% (0.00 km2) | 11.74% (300.02 km2) |
| Static water | 0.81% (20.51 km2) | 5.52% (141.14 km2) |
| Flowing water | 0.25% (6.24 km2) | 0.25% (6.33 km2) |
| An. gambiae s.l. | An. funestus | An. paludis | ||||
|---|---|---|---|---|---|---|
| Importance Ranking | Variable | Importance | Variable | Importance | Variable | Importance |
| 1 | LST-4 | 13.57 | rainfall-2 | 11.61 | rainfall-1 | 11.11 |
| 2 | rainfall0 | 12.93 | LST-5 | 9.91 | rainfall0 | 9.57 |
| 3 | rainfall-1 | 11.83 | LST-1 | 9.54 | rainfall-2 | 9.21 |
| 4 | LST-3 | 10.51 | rainfall-3 | 8.56 | LST-1 | 8.90 |
| 5 | rainfall-5 | 10.16 | PCforfal | 7.85 | LST0 | 8.77 |
| 6 | rainfall-2 | 9.70 | PCforest | 7.57 | LST-4 | 8.53 |
| 7 | rainfall-3 | 9.49 | distforest | 7.53 | rainfall-5 | 7.74 |
| 8 | LST-2 | 9.42 | PCswamp | 7.46 | PCgrass | 7.26 |
| 9 | rainfall-4 | 9.34 | distforfal | 7.29 | distsw | 7.24 |
| 10 | LST-5 | 8.40 | PCgrass | 7.28 | rainfall-4 | 7.19 |
| 11 | PCgrass | 8.03 | distswamp | 7.26 | LST-3 | 6.93 |
| 12 | PCswamp | 6.84 | LST0 | 6.81 | PCforest | 6.78 |
| 13 | LST0 | 6.73 | elevation | 6.78 | distswamp | 6.76 |
| 14 | LST-1 | 6.60 | distsw | 6.70 | elevation | 6.66 |
| 15 | distforest | 6.59 | rainfall-1 | 6.55 | distforest | 6.59 |
| 16 | PCforfal | 6.11 | LST-4 | 6.29 | PCsw | 6.55 |
| 17 | distswamp | 5.93 | PCsw | 5.46 | LST-2 | 6.16 |
| 18 | distforfal | 4.97 | rainfall-5 | 5.33 | NDWI | 5.82 |
| 19 | PCsw | 4.95 | distfal | 5.13 | LST-5 | 5.67 |
| 20 | PCforest | 4.80 | PCag | 4.92 | rainfall-3 | 5.53 |
| 21 | PCbu | 4.74 | LST-3 | 4.81 | NDVI | 5.33 |
| 22 | distsw | 4.60 | rainfall0 | 4.72 | PCswamp | 5.31 |
| 23 | PCfw | 4.58 | rainfall-4 | 4.72 | PCforfal | 4.83 |
| 24 | elevation | 4.48 | PCfal | 4.48 | distforfal | 4.53 |
| 25 | NDVI | 4.41 | MNDWI | 4.41 | distfw | 3.24 |
| 26 | distfw | 3.52 | LST-2 | 3.73 | PCfal | 3.09 |
| 27 | NDWI | 3.44 | NDVI | 3.35 | ||
| 28 | distfal | 3.43 | NDWI | 3.34 | ||
| 29 | PCfal | 3.39 | aspect | 2.97 | ||
| 30 | PCag | 3.01 | distfw | 2.80 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marston, C.; Rowland, C.; O’Neil, A.; Irish, S.; Wat’senga, F.; Martín-Gallego, P.; Aplin, P.; Giraudoux, P.; Strode, C. Developing the Role of Earth Observation in Spatio-Temporal Mosquito Modelling to Identify Malaria Hot-Spots. Remote Sens. 2023, 15, 43. https://doi.org/10.3390/rs15010043
Marston C, Rowland C, O’Neil A, Irish S, Wat’senga F, Martín-Gallego P, Aplin P, Giraudoux P, Strode C. Developing the Role of Earth Observation in Spatio-Temporal Mosquito Modelling to Identify Malaria Hot-Spots. Remote Sensing. 2023; 15(1):43. https://doi.org/10.3390/rs15010043
Chicago/Turabian StyleMarston, Christopher, Clare Rowland, Aneurin O’Neil, Seth Irish, Francis Wat’senga, Pilar Martín-Gallego, Paul Aplin, Patrick Giraudoux, and Clare Strode. 2023. "Developing the Role of Earth Observation in Spatio-Temporal Mosquito Modelling to Identify Malaria Hot-Spots" Remote Sensing 15, no. 1: 43. https://doi.org/10.3390/rs15010043
APA StyleMarston, C., Rowland, C., O’Neil, A., Irish, S., Wat’senga, F., Martín-Gallego, P., Aplin, P., Giraudoux, P., & Strode, C. (2023). Developing the Role of Earth Observation in Spatio-Temporal Mosquito Modelling to Identify Malaria Hot-Spots. Remote Sensing, 15(1), 43. https://doi.org/10.3390/rs15010043