
Cyanobacteria Index as a Tool for the Satellite Detection of Cyanobacteria Blooms in the Baltic Sea

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
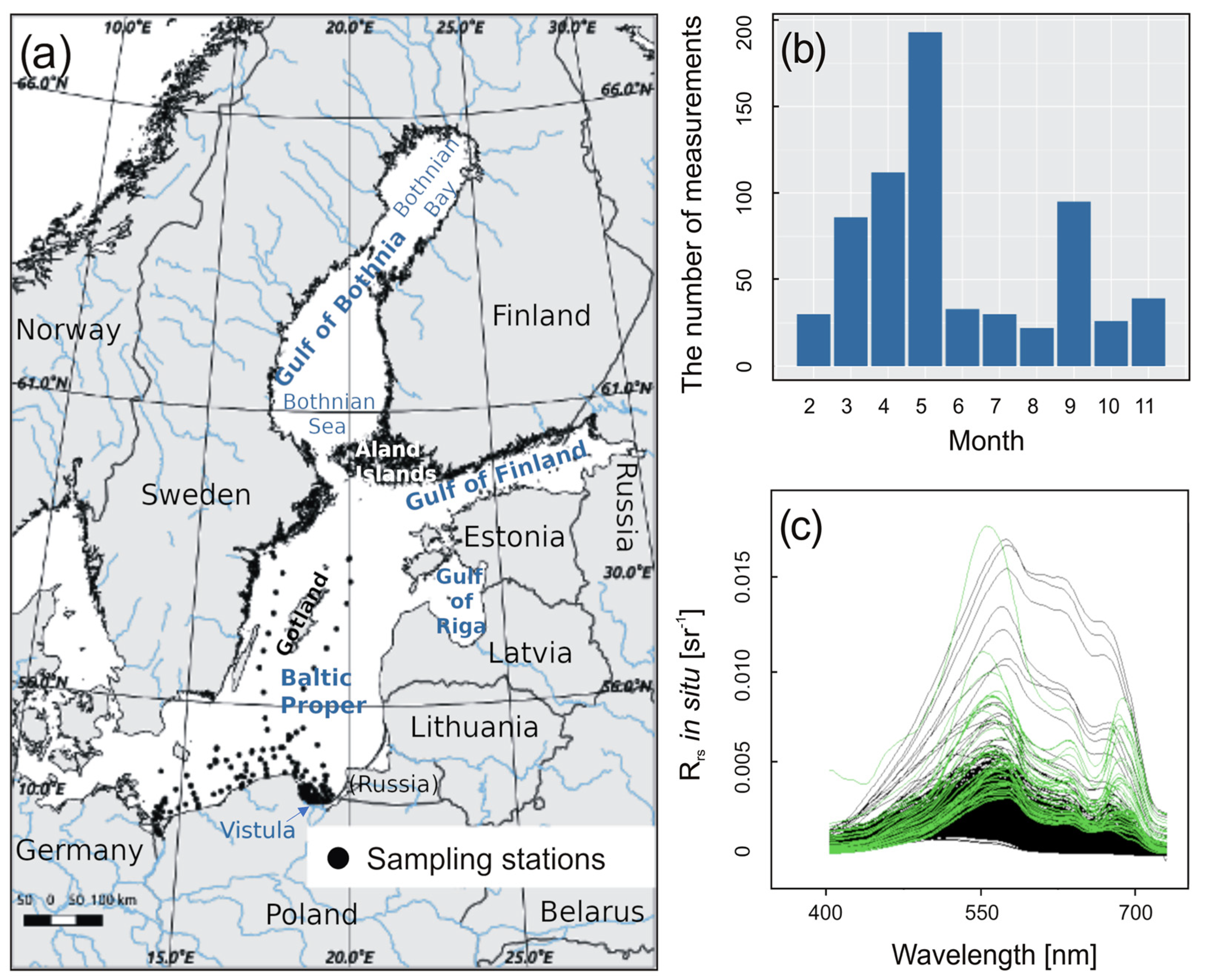
2.1. Study Area
2.2. In Situ Measurements
2.2.1. Pigment Concentrations
2.2.2. In Situ Remote Sensing Reflectance
2.3. Satellite Data
2.4. Cyanobacteria Index
2.5. Statistical Analyses
3. Results
3.1. CI Comparison for the Two Satellite Sensors
3.2. Bloom Identification Efficiency
3.3. Spatial and Temporal Changes of the Cyanobacteria Blooms in the Baltic Sea
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazur-Marzec, H.; Pliński, M. Do toxic cyanobacteria blooms pose a threat to the Baltic ecosystem? Oceanologia 2009, 51, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Reinart, A.; Kutser, T. Comparison of different satellite sensors in detecting cyanobacterial bloom events in the Baltic Sea. Remote Sens. Environ. 2006, 102, 74–85. [Google Scholar] [CrossRef]
- Ploug, H. Cyanobacterial surface blooms formed by Aphanizomenon sp. and Nodularia spumigena in the Baltic Sea: Small-scale fluxes, pH, and oxygen microenvironments. Limnol. Oceanogr. 2008, 53, 914–921. [Google Scholar] [CrossRef] [Green Version]
- Bresciani, M.; Adamo, M.; de Carolis, G.; Matta, E.; Pasquariello, G.; Vaičiūtė, D.; Giardino, C. Monitoring blooms and surface accumulation of cyanobacteria in the Curonian Lagoon by combining MERIS and ASAR data. Remote Sens. Environ. 2014, 146, 124–135. [Google Scholar] [CrossRef]
- Kahru, M.; Leppänen, J.-M.; Rud, O. Cyanobacterial blooms cause heating of the sea surface. Mar. Ecol. Prog. Ser. 1993, 101, 1–7. [Google Scholar] [CrossRef]
- Kanoshina, I.; Lips, U.; Leppänen, J.-M. The influence of weather conditions (temperature and wind) on cyanobacterial bloom development in the Gulf of Finland (Baltic Sea). Harmful Algae 2003, 2, 29–41. [Google Scholar] [CrossRef]
- Nebaeus, M. Algal water-blooms under ice-cover. Verh. Internat. Verein. Limnol. 1971, 22, 719–724. [Google Scholar] [CrossRef]
- Steidinger, K.A.; Garccés, E. Importance of Life Cycles in the Ecology of Harmful Microalgae. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-32210-8. [Google Scholar]
- Hollister, J.W.; Kreakie, B.J. Associations between chlorophyll a and various microcystin health advisory concentrations. F1000Research 2016, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Behrenfeld, M.J.; Westberry, T.K.; Boss, E.S.; O’Malley, R.T.; Siegel, D.A.; Wiggert, J.D.; Franz, B.A.; McClain, C.R.; Feldman, G.C.; Doney, S.C.; et al. Satellite-detected fluorescence reveals global physiology of ocean phytoplankton. Biogeosci. Discuss. 2008, 5, 4235–4270. [Google Scholar] [CrossRef] [Green Version]
- Naghdi, K.; Moradi, M.; Kabiri, K.; Rahimzadegan, M. The effects of cyanobacterial blooms on MODIS-L2 data products in the southern Caspian Sea. Oceanologia 2018, 60, 367–377. [Google Scholar] [CrossRef]
- Govindjee, G. Chlorophyll a Fluorescence: A Bit of Basics and History. In Chlorophyll Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, G., Eds.; Springer: Dordrecht, The Netherlands, 2004; ISBN 978-1-4020-3218-9. [Google Scholar]
- Dall’Olmo, G.; Gitelson, A.A. Effect of bio-optical parameter variability and uncertainties in reflectance measurements on the remote estimation of chlorophyll-a concentration in turbid productive waters: Modeling results. Appl. Opt. 2006, 45, 3577–3598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, S.; Grossman, A. Photoprotection in Cyanobacteria: Regulation of Light Harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Llewellyn, C.A.; Egeland, E.S.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2011; 890p, ISBN 978-1-107-00066-7. [Google Scholar]
- Wojtasiewicz, B.; Stoń-Egiert, J. Bio-optical characterization of selected cyanobacteria strains present in marine and freshwater ecosystems. J. Appl. Phycol. 2016, 28, 2299–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woźniak, M.; Bradtke, K.; Darecki, M.; Krężel, A. Empirical Model for Phycocyanin Concentration Estimation as an Indicator of Cyanobacterial Bloom in the Optically Complex Coastal Waters of the Baltic Sea. Remote Sens. 2016, 8, 212. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, M.; Klawonn, I.; Karlson, B. Nitrogen fixation estimates for the Baltic Sea indicate high rates for the previously overlooked Bothnian Sea. Ambio 2021, 50, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Rantajärvi, E. Alg@line in 2003: 10 Years of Innovative Plankton Monitoring and Research and Operational Information Service in the Baltic Sea; MERI—Report Series of the Finnish Institute of Marine Research; Merentutkimuslaitos: Helsinki, Finland, 2003; p. 48. ISBN 951-53-2507-2. [Google Scholar]
- Kahru, M.; Elmgren, R. Multidecadal time series of satellite-detected accumulations of cyanobacteria in the Baltic Sea. Biogeosciences 2014, 11, 3619–3633. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Elmgren, R.; Savchuk, O.P. Changing seasonality of the Baltic Sea. Biogeosciences 2016, 13, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Kutser, T. Passive optical remote sensing of cyanobacteria and other intense phytoplankton blooms in coastal and inland waters. Int. J. Remote Sens. 2009, 30, 4401–4425. [Google Scholar] [CrossRef]
- Philpot, W.D. The Derivative Ratio Algorithm: Avoiding Atmospheric Effects in Remote Sensing. IEEE Trans. Geosci. Remote Sens. 1991, 29, 350–357. [Google Scholar] [CrossRef]
- Schowengerdt, R.A. Remote Sensing: Models and Methods for Image Processing; Elsevier: Enschede, The Netherlands, 2006; ISBN 9780080480589. [Google Scholar]
- Stumpf, R.P.; Werdell, P.J. Adjustment of ocean color sensor calibration through multi-band statistics. Opt. Express 2010, 18, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Wynne, T.T.; Stumpf, R.P.; Briggs, T.O. Comparing MODIS and MERIS spectral shapes for cyanobacterial bloom detection. Int. J. Remote Sens. 2013, 34, 6668–6678. [Google Scholar] [CrossRef]
- Stumpf, R.P.; Davis, T.W.; Wynne, T.T.; Graham, J.L.; Loftin, K.A.; Johengen, T.H.; Gossiaux, D.; Palladino, D.; Burtner, A. Challenges for mapping cyanotoxin patterns from remote sensing of cyanobacteria. Harmful Algae 2016, 54, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Stumpf, R.P.; Schaeffer, B.A.; Werdell, J.P.; Loftin, K.A.; Meredith, A. Measurement of Cyanobacterial Bloom Magnitude using Satellite Remote Sensing. Sci. Rep. 2019, 9, 18310. [Google Scholar] [CrossRef] [Green Version]
- Leppäranta, M.; Myrberg, K. Physical Oceanography of the Baltic Sea; Springer Science & Business Media: Berlin, Germany, 2009; ISBN 978-3-540-79702-9. [Google Scholar]
- Andrén, E.; Snoeijs-Leijonmalm, P. Why is the Baltic Sea so special to live in? In Biological Oceanography of the Baltic Sea; Snoeijs-Leijonmalm, P., Schubert, H., Radziejewska, T., Eds.; Springer: Dordrecht, The Netherlands, 2017; ISBN 978-94-007-0667-5. [Google Scholar]
- Pastuszak, M.; Kowalkowski, T.; Kopiński, J.; Doroszewski, A.; Jurga, B.; Buszewski, B. Long-term changes in nitrogen and phosphorus emission into the Vistula and Oder catchments (Poland)—Modeling (MONERIS) studies. Environ. Sci. Pol. Res. 2018, 25, 29734–29751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur-Marzec, H.; Kężel, A.; Kobos, J.; Pliński, M. Toxic Nodularia spumigena blooms in the coastal waters of the Gulf of Gdańsk: A ten-year survey. Oceanologia 2006, 48, 255–273. [Google Scholar]
- Stoń-Egiert, J.; Ostrowska, M. Long-term changes in phytoplankton pigment contents in the Baltic Sea: Trends and spatial variability during 20 years of investigations. Cont. Shelf Res. 2022, 236, 104666. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S. Ecology of Harmful Cyanobacteria. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-32210-8. [Google Scholar]
- Stoń, J.; Kosakowska, A. Phytoplankton pigments designation–an application of RP-HPLC in qualitative and quantitative analysis. J. Appl. Phycol. 2002, 14, 205–210. [Google Scholar] [CrossRef]
- Stoń-Egiert, J.; Kosakowska, A. RP-HPLC determination of phytoplankton pigments—Comparison of calibration results for two columns. Mar. Biol. 2005, 147, 251–260. [Google Scholar] [CrossRef]
- Sobiechowska-Sasim, M.; Stoń-Egiert, J.; Kosakowska, A. Quantitative analysis of extracted phycobilin pigments in cyanobacteria—An assessment of spectrophotometric and spectrofluorometric methods. J. Appl. Phycol. 2014, 26, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Gordon, H.R.; Ding, K. Self-shading of in-water optical instruments. Limnol. Oceanogr. 1992, 37, 491–500. [Google Scholar] [CrossRef]
- Zibordi, G.; Ferrari, G.M. Instrument self-shading in underwater optical measurements: Experimental data. Appl. Opt. 1995, 34, 2750–2754. [Google Scholar] [CrossRef]
- Mueller, J.L.; Fargion, G.S.; McClain, C.R. Ocean Optics Protocols for Satellite Ocean Color Sensor Validation—Revision 4, Volume III: Radiometric Measurements and Data Analysis Protocols; NASA Technical Report; NASA/TM-2003-21621/Rev-Vol III; NASA: Greenbelt, MD, USA, 2003.
- Austin, R.W. Optical aspects of oceanography. In The Remote Sensing of Spectral Radiance from below the Ocean Surface; Jerlov, N.G., Nielsen, E.S., Eds.; Academic Press: New York, NY, USA, 1974; pp. 317–344. [Google Scholar]
- Pelloquin, C.; Nieke, J. Sentinel-3 OLCI and SLSTR Simulated Spectral Response Functions S3-TN-ESA-PL-316. In Proceedings of the Sentinel-3 OLCI/SLSTR and MERIS/(A)ATSR, 711(2), ESA Communications, Noordwijk, The Netherlands, 15–19 October 2012. [Google Scholar]
- Bailey, S.W.; Werdell, P.J. A multi-sensor approach for the on-orbit validation of ocean color satellite data products. Remote Sens. Environ. 2006, 102, 12–23. [Google Scholar] [CrossRef]
- Wynne, T.T.; Stumpf, R.P.; Tomlinson, M.C.; Dyble, J. Characterizing a cyanobacterial bloom in western Lake Erie using satellite imagery and meteorological data. Limnol. Oceanogr. 2010, 55, 2025–2036. [Google Scholar] [CrossRef] [Green Version]
- Chai, T.; Draxler, R.R. Root mean square error (RMSE) or mean absolute error (MAE)?—Arguments against avoiding RMSE in the literature. Geosci. Model Dev. 2014, 7, 1247–1250. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; R Core Team; et al. Caret: Classification and Regression Training. R package version 6.0-84. 2020. Available online: https://cran.r-project.org/web/packages/caret/index.html (accessed on 17 October 2022).
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Pawlik, M.; Ficek, D. Pine pollen grains in coastal waters of the Baltic Sea. Oceanol. Hydrobiol. Stud. 2016, 45, 35–41. [Google Scholar] [CrossRef]
- Schlüter, L.; Lauridsen, T.L.; Krogh, G.; Jørgensen, T. Identification and quantification of phytoplankton groups in lakes using new pigment ratios—A comparison between pigment analysis by HPLC and microscopy. Freshw. Biol. 2006, 51, 1474–1485. [Google Scholar] [CrossRef]
- Qi, L.; Hu, C.; Duan, H.; Cannizzaro, J.; Ma, R. A novel MERIS algorithm to derive cyanobacterial phycocyanin pigment concentrations in a eutrophic lake: Theoretical basis and practical considerations. Remote Sens. Environ. 2014, 154, 298–317. [Google Scholar] [CrossRef]
- Attila, S.; Fleming-Lehtinen, V.; Attila, J.; Junttila, S.; Alasalmi, H.; Hällfors, H.; Kervinen, M.; Koponen, S. A novel earth observation based ecological indicator for cyanobacterial blooms. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 145–155. [Google Scholar] [CrossRef]
- Gower, J.; King, S. On the importance of a band at 709 nm. In Proceedings of the 2nd MERIS (A)ATSR User Workshop, Frascati, Italy, 22–26 September 2008. [Google Scholar]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- McHugh, M.L. Interrater reliability: The kappa statistic. Biochem. Med. 2012, 22, 276–282. [Google Scholar] [CrossRef]
- Kahru, M.; Savchuk, O.P.; Elmgren, R. Satellite measurements of cyanobacterial bloom frequency in the Baltic Sea: Interannual and spatial variability. Mar. Ecol. Prog. Ser. 2007, 343, 15–23. [Google Scholar] [CrossRef]
- Kahru, M.; Elmgren, R.; Kaiser, J.; Wasmund, N.; Savchuk, O. Cyanobacterial Blooms in the Baltic Sea: Correlations with Environmental Factors. Harmful Algae 2020, 92, 101739. [Google Scholar] [CrossRef]
- Öberg, J. Cyanobacteria blooms in the Baltic Sea. HELCOM Baltic Sea Environment Fact Sheet 2014. Available online: https://helcom.fi/baltic-sea-trends/environment-fact-sheets/eutrophication/cyanobacteria-biomass/ (accessed on 30 December 2021).
- Copernicus. Early Algal Bloom in the Baltic Sea. Image of the Day Series, EU, Copernicus Senitnel-2 Imagery. 2021. Available online: https://www.copernicus.eu/en/media/image-day-gallery/early-algal-bloom-baltic-sea (accessed on 30 December 2021).
- Groetsch, P.M.M.; Simis, S.G.H.; Eleveld, M.A.; Peters, S.W.M. Cyanobacterial bloom detection based on coherence between ferrybox observations. J. Mar. Syst. 2014, 140, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Hu, C. A novel ocean color index to detect floating algae in the global oceans. Remote Sens. Environ. 2009, 113, 2118–2129. [Google Scholar] [CrossRef]
- Gilerson, A.; Zhou, J.; Hlaing, S.; Ioannou, I.; Gross, B.; Moshary, F.; Ahmed, S. Fluorescence component in the reflectance spectra from coastal waters. II. Performance of retrieval algorithms. Opt. Express 2008, 16, 2446–2460. [Google Scholar] [CrossRef] [PubMed]
- Kowalczuk, P.; Darecki, M.; Zabłocka, M.; Górecka, I. Validation of empirical and semi-analytical remote sensing algorithms for estimating absorption by Coloured Dissolved Organic Matter in the Baltic Sea from SeaWiFS and MODIS imagery. Oceanologia 2010, 52, 171–196. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Yoshida, T.; Kashima, A.; Takashima, Y.; Hosoda, N.; Nagasaki, K.; Hiroishi, S. Ecological Dynamics of the Toxic Bloom-Forming Cyanobacterium Microcystis aeruginosa and Its Cyanophages in Freshwater. Appl. Environ. Microbiol. 2008, 74, 3269–3273. [Google Scholar] [CrossRef] [Green Version]
- Śliwińska-Wilczewska, S.; Cieszyńska, A.; Konik, M.; Maculewicz, J.; Latała, A. Environmental drivers of bloom-forming cyanobacteria in the Baltic Sea: Effects of salinity, temperature, and irradiance. Estuar. Coast. Shelf Sci. 2019, 219, 139–150. [Google Scholar] [CrossRef]
- Wasmund, N.; Nausch, G.; Gerth, M.; Busch, S.; Burmeister, C.; Hansen, R.; Sadkowiak, B. Extension of the growing season of phytoplankton in the western Baltic Sea in response to climate change. Mar. Ecol. Prog. Ser. 2019, 622, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rolff, C.; Elfwing, T. Increasing nitrogen limitation in the Bothnian Sea, potentially caused by inflow of phosphate-rich water from the Baltic Proper. Ambio 2005, 44, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kownacka, J.; Busch, S.; Göbel, J.; Gromisz, S.; Hällfors, H.; Höglander, H.; Huseby, S.; Jaanus, A.; Jakobsen, H.H.; Johansen, M.; et al. Cyanobacteria biomass 1990–2018. HELCOM Baltic Sea Environment Fact Sheets 2018. Available online: https://helcom.fi/baltic-sea-trends/environment-fact-sheets/eutrophication/cyanobacteria-biomass/ (accessed on 2 September 2020).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of OLCI Pixels | ||||
|---|---|---|---|---|
| Class 0 (No Bloom) | Class 1 (Bloom) | Sum | ||
| Number of MODIS pixels | Class 0 (no bloom) | 1,368,696 | 22,976 | 1,391,672 |
| Class 1 (bloom) | 27,113 | 62,439 | 89,552 | |
| Total | 1,395,809 | 85,415 | κ = 0.70 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konik, M.; Bradtke, K.; Stoń-Egiert, J.; Soja-Woźniak, M.; Śliwińska-Wilczewska, S.; Darecki, M. Cyanobacteria Index as a Tool for the Satellite Detection of Cyanobacteria Blooms in the Baltic Sea. Remote Sens. 2023, 15, 1601. https://doi.org/10.3390/rs15061601
Konik M, Bradtke K, Stoń-Egiert J, Soja-Woźniak M, Śliwińska-Wilczewska S, Darecki M. Cyanobacteria Index as a Tool for the Satellite Detection of Cyanobacteria Blooms in the Baltic Sea. Remote Sensing. 2023; 15(6):1601. https://doi.org/10.3390/rs15061601
Chicago/Turabian StyleKonik, Marta, Katarzyna Bradtke, Joanna Stoń-Egiert, Monika Soja-Woźniak, Sylwia Śliwińska-Wilczewska, and Mirosław Darecki. 2023. "Cyanobacteria Index as a Tool for the Satellite Detection of Cyanobacteria Blooms in the Baltic Sea" Remote Sensing 15, no. 6: 1601. https://doi.org/10.3390/rs15061601
APA StyleKonik, M., Bradtke, K., Stoń-Egiert, J., Soja-Woźniak, M., Śliwińska-Wilczewska, S., & Darecki, M. (2023). Cyanobacteria Index as a Tool for the Satellite Detection of Cyanobacteria Blooms in the Baltic Sea. Remote Sensing, 15(6), 1601. https://doi.org/10.3390/rs15061601