
Increasing Forest Cover and Connectivity Both Inside and Outside of Protected Areas in Southwestern Costa Rica

 , , , and
, , , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. LULC Map Creation
2.3. NDVI and EVI Map Creation
2.4. LULC Map Validation
2.5. LULC Data Analysis
2.6. Landscape Metrics Analysis
3. Results
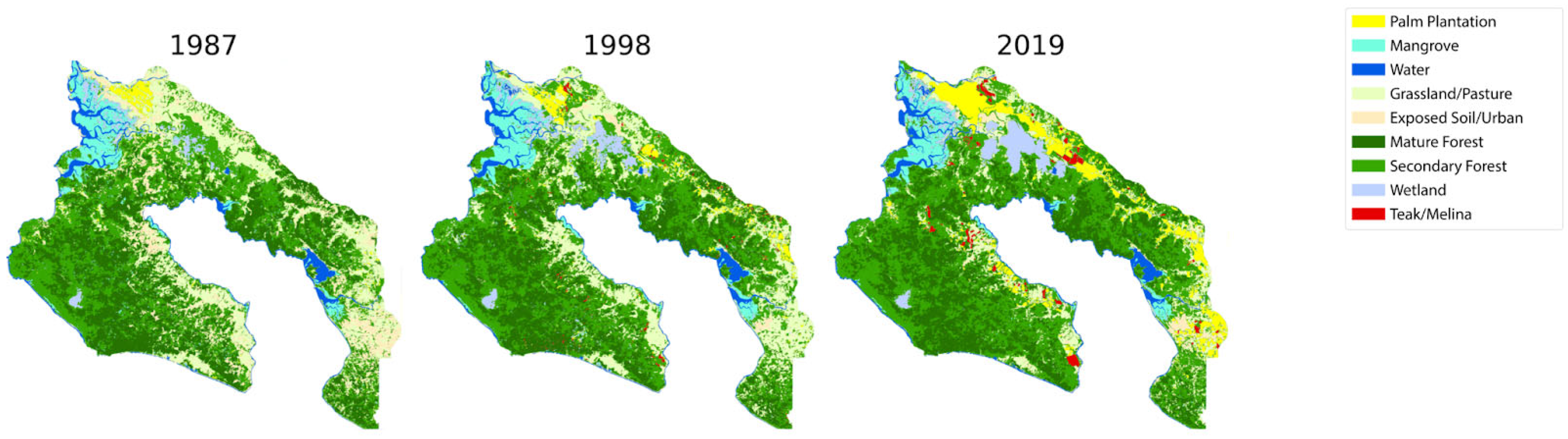
3.1. Overall LULC in the Study Area
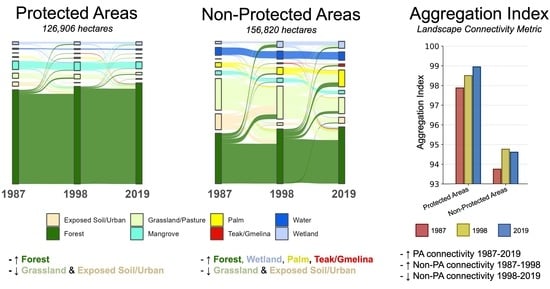
3.2. LULC Changes in PAs
3.3. LULC Changes in Riparian Zones, Roadsides, Towns, and PES Farms
3.4. LULC Accuracy
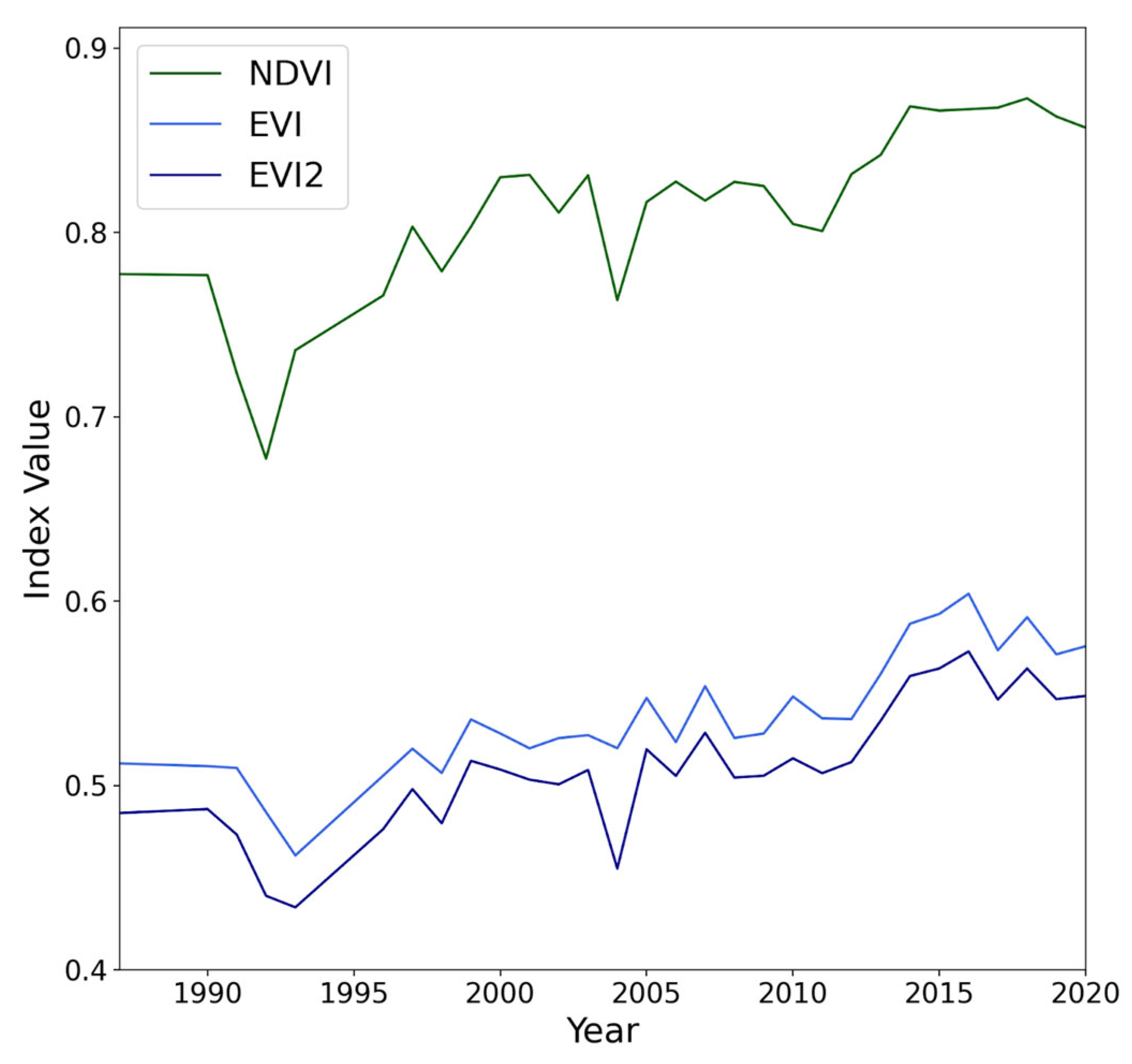
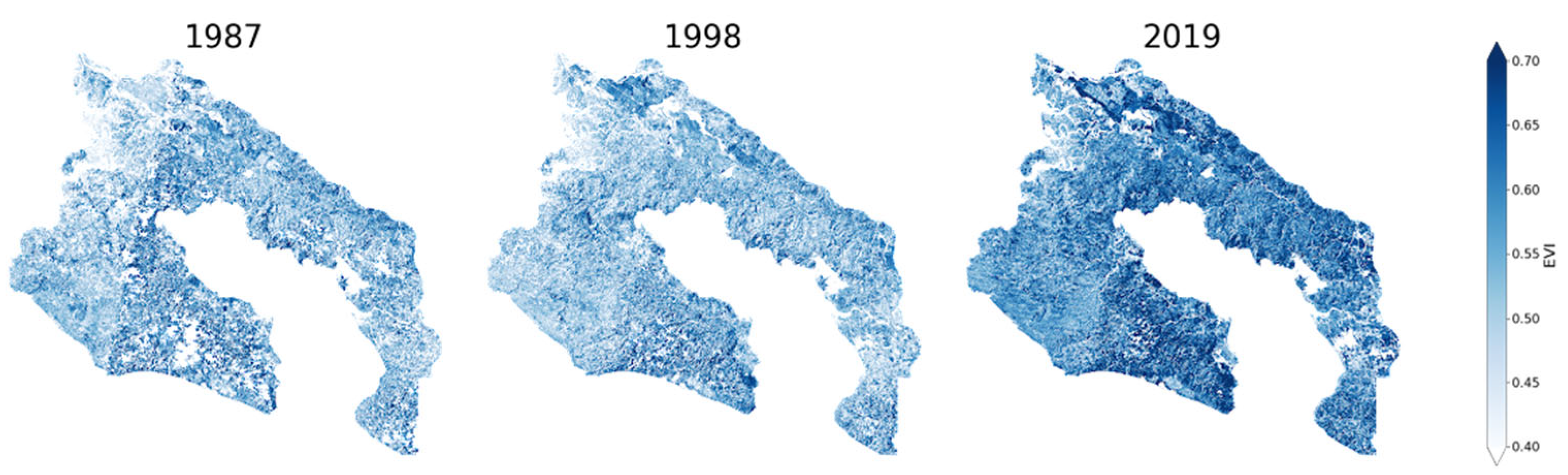
3.5. NDVI and EVI Change
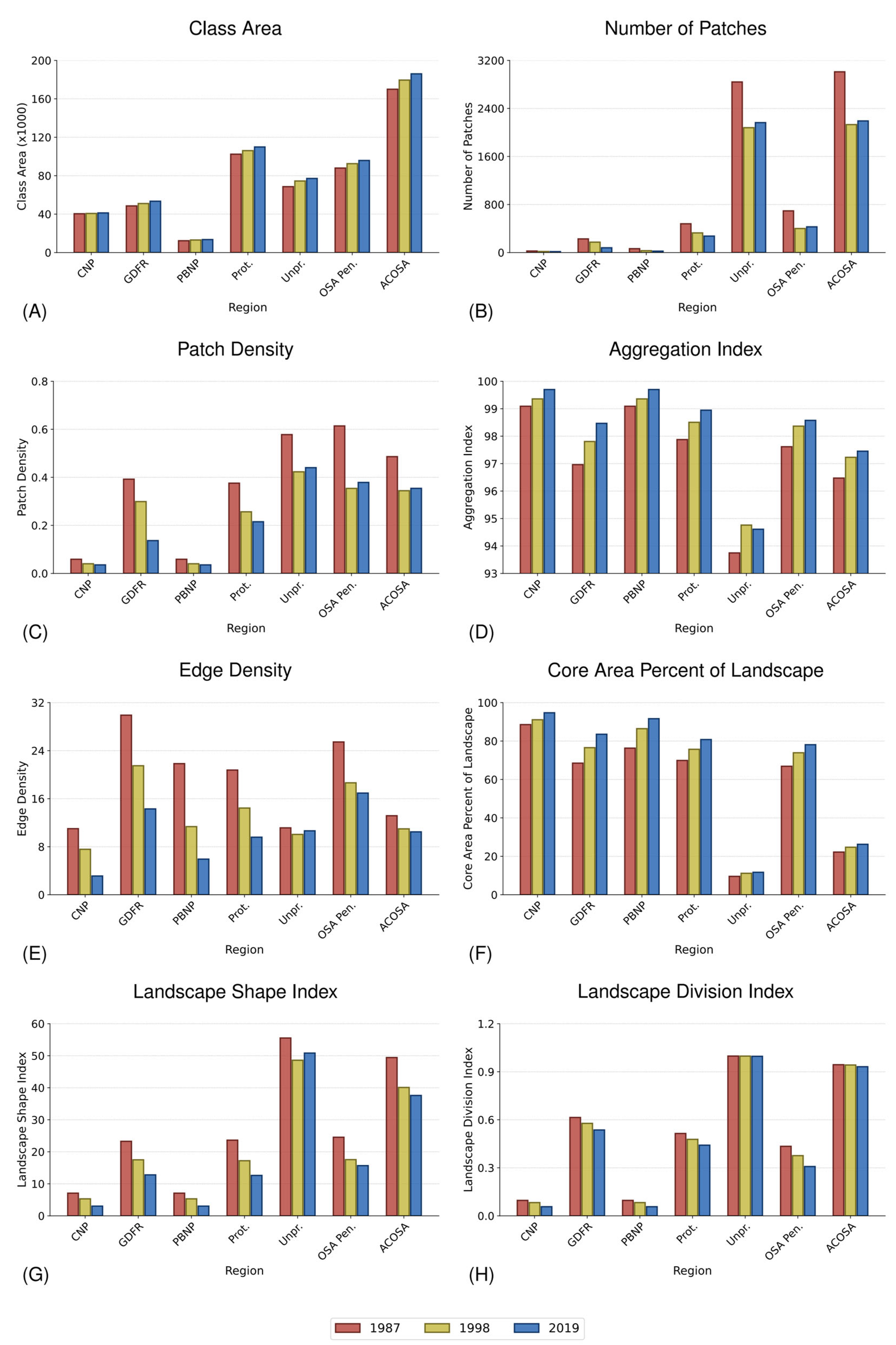
3.6. Landscape Metrics
4. Discussion
4.1. LULC Dynamics Reflect Osa’s Globalization Process
4.2. Forest Cover Increased in PAs, Indicating Almost No Encroachment, which Is Rare in the Tropics
4.3. Forest Cover Also Increased Outside of PAs, Suggesting Minimal Local Deforestation Leakage from PAs
4.4. Both Forest Conversion and Restoration Rates Decreased after 1996 Forest Law
4.5. Connectivity Increased across the Region, with PAs Having Higher Connectivity Than Non-PAs
4.6. Non-PAs Saw Increasing Forest Cover yet Slightly Decreasing Connectivity 1998–2019
4.7. Forest Cover Increased in Properties Receiving Payments for Ecosystem Services
4.8. Riparian Zones Had Low Forest Cover but Saw Larger Increases over the Study Period
4.9. Roadsides Saw the Largest Increase in Forest Cover
4.10. Study Limitations and Further Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LULC Class | Quantity | Accurate Count | Producer’s Accuracy |
|---|---|---|---|
| Palm | 73 | 57 | 78.08% |
| Mangrove | 87 | 80 | 91.95% |
| Water | 56 | 52 | 92.86% |
| Grassland | 66 | 45 | 68.18% |
| Exposed/urban | 50 | 41 | 82.00% |
| Mature forest | 112 | 73 | 65.18% |
| Secondary forest | 166 | 103 | 62.05% |
| Wetland | 67 | 65 | 97.01% |
| Teak/gmelina | 66 | 56 | 84.85% |
| Grand Total | 743 | 572 | 76.99% |
| LULC Class | Quantity | Accurate Count | Producer’s Accuracy |
|---|---|---|---|
| Palm | 73 | 57 | 78.08% |
| Mangrove | 87 | 80 | 91.95% |
| Water | 56 | 52 | 92.86% |
| Grassland | 66 | 45 | 68.18% |
| Exposed/urban | 50 | 41 | 82.00% |
| Forest | 278 | 276 | 99.28% |
| Wetland | 67 | 65 | 97.01% |
| Teak/gmelina | 66 | 56 | 84.85% |
| Grand Total | 743 | 672 | 90.44% |
| LULC MAP (PREDICTED) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Palm | Mangrove | Water | Grassland | Exposed/Urban | Old Growth Forest | Secondary Forest | Wetland | Teak/Gmelina | SUM | PRODUCER’S ACCURACY | ||
| VERIFICATION POINTS (ACTUAL) | Palm | 57 | 0 | 0 | 1 | 2 | 1 | 12 | 0 | 0 | 73 | 78.08% |
| Mangrove | 0 | 80 | 4 | 0 | 0 | 0 | 1 | 2 | 0 | 87 | 91.95% | |
| Water | 1 | 1 | 52 | 0 | 0 | 0 | 1 | 1 | 0 | 56 | 92.86% | |
| Grassland | 4 | 0 | 0 | 45 | 2 | 0 | 15 | 0 | 0 | 66 | 68.18% | |
| Exposed/urban | 0 | 0 | 0 | 9 | 41 | 0 | 0 | 0 | 0 | 50 | 82.00% | |
| Old growth forest | 0 | 0 | 0 | 0 | 0 | 73 | 39 | 0 | 0 | 112 | 65.18% | |
| Secondary forest | 1 | 0 | 0 | 0 | 1 | 61 | 103 | 0 | 0 | 166 | 62.05% | |
| Wetland | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 65 | 0 | 67 | 97.01% | |
| Teak/gmelina | 4 | 0 | 0 | 1 | 0 | 0 | 5 | 0 | 56 | 66 | 84.85% | |
| SUM | 67 | 82 | 56 | 56 | 46 | 135 | 177 | 68 | 56 | |||
| USER’S ACCURACY | 85.07% | 97.56% | 92.86% | 80.36% | 89.13% | 54.07% | 58.19% | 95.59% | 100.00% | |||
| LULC MAP (PREDICTED) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Palm | Mangrove | Water | Grassland | Exposed/Urban | Forest | Wetland | Teak/gmelina | SUM | PRODUCER’S ACCURACY | ||
| VERIFICATION POINTS (ACTUAL) | Palm | 57 | 0 | 0 | 1 | 2 | 13 | 0 | 0 | 73 | 78.08% |
| Mangrove | 0 | 80 | 4 | 0 | 0 | 1 | 2 | 0 | 87 | 91.95% | |
| Water | 1 | 1 | 52 | 0 | 0 | 1 | 1 | 0 | 56 | 92.86% | |
| Grassland | 4 | 0 | 0 | 45 | 2 | 15 | 0 | 0 | 66 | 68.18% | |
| Exposed/urban | 0 | 0 | 0 | 9 | 41 | 0 | 0 | 0 | 50 | 82.00% | |
| Forest | 0 | 0 | 0 | 0 | 0 | 276 | 0 | 0 | 276 | 100.00% | |
| Wetland | 0 | 1 | 0 | 0 | 0 | 1 | 65 | 0 | 67 | 97.01% | |
| Teak/gmelina | 4 | 0 | 0 | 1 | 0 | 5 | 0 | 56 | 66 | 84.85% | |
| SUM | 66 | 82 | 56 | 56 | 45 | 312 | 68 | 56 | |||
| USER’S ACCURACY | 86.36% | 97.56% | 92.86% | 80.36% | 91.11% | 88.46% | 95.59% | 100.00% | |||






References
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future Threats to Biodiversity and Pathways to Their Prevention. Nature 2017, 546, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Venter, O.; Fuller, R.A.; Allan, J.R.; Maxwell, S.L.; Negret, P.J.; Watson, J.E.M. One-Third of Global Protected Land Is under Intense Human Pressure. Science 2018, 360, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Dinerstein, E.; Vynne, C.; Sala, E.; Joshi, A.R.; Fernando, S.; Lovejoy, T.E.; Mayorga, J.; Olson, D.; Asner, G.P.; Baillie, J.E.M.; et al. A Global Deal for Nature: Guiding Principles, Milestones, and Targets. Sci. Adv. 2019, 5, eaaw2869. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F.; Carolina Useche, D.; Rendeiro, J.; Kalka, M.; Bradshaw, C.J.A.; Sloan, S.P.; Laurance, S.G.; Campbell, M.; Abernethy, K.; Alvarez, P.; et al. Averting Biodiversity Collapse in Tropical Forest Protected Areas. Nature 2012, 489, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Levi, T.; Ripple, W.J.; Zárrate-Charry, D.A.; Betts, M.G. A Forest Loss Report Card for the World’s Protected Areas. Nat. Ecol. Evol. 2021, 5, 520–529. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; With, K.A. Landscape Connectivity: A Return to the Basics. In Connectivity Conservation; Crooks, K.R., Sanjayan, M., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 29–43. ISBN 978-0-521-85706-2. [Google Scholar]
- Ford, S.A.; Jepsen, M.R.; Kingston, N.; Lewis, E.; Brooks, T.M.; MacSharry, B.; Mertz, O. Deforestation Leakage Undermines Conservation Value of Tropical and Subtropical Forest Protected Areas. Glob. Ecol. Biogeogr. 2020, 29, 2014–2024. [Google Scholar] [CrossRef]
- Rayfield, B.; Pelletier, D.; Dumitru, M.; Cardille, J.A.; Gonzalez, A. Multipurpose Habitat Networks for Short-Range and Long-Range Connectivity: A New Method Combining Graph and Circuit Connectivity. Methods Ecol. Evol. 2016, 7, 222–231. [Google Scholar] [CrossRef]
- Allen, K.E.; Padgett Vásquez, S. Forest Cover, Development, and Sustainability in Costa Rica: Can One Policy Fit All? Land Use Policy 2017, 67, 212–221. [Google Scholar] [CrossRef]
- Le Coq, J.-F.; Froger, G.; Pesche, D.; Legrand, T.; Saenz, F. Understanding the Governance of the Payment for Environmental Services Programme in Costa Rica: A Policy Process Perspective. Ecosyst. Serv. 2015, 16, 253–265. [Google Scholar] [CrossRef]
- Ward, M.; Saura, S.; Williams, B.; Ramírez-Delgado, J.P.; Arafeh-Dalmau, N.; Allan, J.R.; Venter, O.; Dubois, G.; Watson, J.E.M. Just Ten Percent of the Global Terrestrial Protected Area Network Is Structurally Connected via Intact Land. Nat. Commun. 2020, 11, 4563. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B.J. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure; Gen Tech Rep PNW-GTR-351; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1995; 122p. [CrossRef]
- Senior, R.A.; Hill, J.K.; Edwards, D.P. Global Loss of Climate Connectivity in Tropical Forests. Nat. Clim. Chang. 2019, 9, 623–626. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Hansen, M.C.; Wang, L.; Song, X.-P.; Tyukavina, A.; Turubanova, S.; Potapov, P.V.; Stehman, S.V. The Fate of Tropical Forest Fragments. Sci. Adv. 2020, 6, eaax8574. [Google Scholar] [CrossRef]
- Martinez Pardo, J.; Saura, S.; Insaurralde, A.; Di Bitetti, M.S.; Paviolo, A.; De Angelo, C. Much More than Forest Loss: Four Decades of Habitat Connectivity Decline for Atlantic Forest Jaguars. Landsc. Ecol. 2023, 38, 41–57. [Google Scholar] [CrossRef]
- Rueda, X.; Thomas, N.E.; Lambin, E.F. Eco-Certification and Coffee Cultivation Enhance Tree Cover and Forest Connectivity in the Colombian Coffee Landscapes. Reg. Environ. Chang. 2015, 15, 25–33. [Google Scholar] [CrossRef]
- Li, L.; Gou, M.; Wang, N.; La, L.; Liu, C. Do Ecological Restoration Programs Reduce Forest Fragmentation? Case Study of the Three Gorges Reservoir Area, China. Ecol. Eng. 2021, 172, 106410. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rivas, C.A.; Quinto, L.; Navarro, S.H.; Varo-Martínez, M.Á.; Palacios-Rodríguez, P. Afforestation on Agricultural Land in Southern Spain: An Important Driver to Improve Forest Landscape Connectivity. New For. 2023, 54, 1061–1084. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M.; Espelta, J.M.; Gordillo, J.; Pino, J. Changes in Forest Landscape Patterns Resulting from Recent Afforestation in Europe (1990–2012): Defragmentation of Pre-Existing Forest versus New Patch Proliferation. Ann. For. Sci. 2020, 77, 1–15. [Google Scholar] [CrossRef]
- Bray, D.B. Forest Cover Dynamics and Forest Transitions in Mexico and Central America: Towards a “Great Restoration”? In Reforesting Landscapes: Linking Pattern and Process; Nagendra, H., Southworth, J., Eds.; Landscape Series; Springer: Dordrecht, The Netherlands, 2010; pp. 85–120. ISBN 978-1-4020-9656-3. [Google Scholar]
- Pagiola, S. Payments for Environmental Services in Costa Rica. Ecol. Econ. 2008, 65, 712–724. [Google Scholar] [CrossRef]
- The Earthshot Prize: Republic of Costa Rica. Available online: https://earthshotprize.org/winners-finalists/costa-rica/ (accessed on 18 July 2023).
- Tafoya, K.A.; Brondizio, E.S.; Johnson, C.E.; Beck, P.; Wallace, M.; Quirós, R.; Wasserman, M.D. Effectiveness of Costa Rica’s Conservation Portfolio to Lower Deforestation, Protect Primates, and Increase Community Participation. Front. Environ. Sci. 2020, 8, 580724. [Google Scholar] [CrossRef]
- Forest Area (% of Land Area)—Costa Rica. Available online: https://data.worldbank.org/indicator/AG.LND.FRST.ZS?locations=CR (accessed on 19 July 2023).
- Hunt, C.A.; Durham, W.H.; Driscoll, L.; Honey, M. Can Ecotourism Deliver Real Economic, Social, and Environmental Benefits? A Study of the Osa Peninsula, Costa Rica. J. Sustain. Tour. 2015, 23, 339–357. [Google Scholar] [CrossRef]
- Zambrano, A.M.A.; Broadbent, E.N.; Durham, W.H. Social and Environmental Effects of Ecotourism in the Osa Peninsula of Costa Rica: The Lapa Rios Case. J. Ecotourism 2010, 9, 62–83. [Google Scholar] [CrossRef]
- Sanchez-Azofeifa, G.A.; Rivard, B.; Calvo, J.; Moorthy, I. Dynamics of Tropical Deforestation Around National Parks: Remote Sensing of Forest Change on the Osa Peninsula of Costa Rica. Mt. Res. Dev. 2002, 22, 352–358. [Google Scholar] [CrossRef]
- Fernandez-Vega, J.; Covey, K.R.; Ashton, M.S. Tamm Review: Large-Scale Infrequent Disturbances and Their Role in Regenerating Shade-Intolerant Tree Species in Mesoamerican Rainforests: Implications for Sustainable Forest Management. For. Ecol. Manag. 2017, 395, 48–68. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Ballesteros, E.; Breedy, O.; Naranjo-Elizondo, B.; Hernández, N.; Salinas-de-León, P.; Sala, E.; Cortés, J. Nearshore Marine Biodiversity of Osa Peninsula, Costa Rica: Where the Ocean Meets the Rainforest. PLoS ONE 2022, 17, e0271731. [Google Scholar] [CrossRef] [PubMed]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, Costa Rica, 1967; Available online: https://search.worldcat.org/title/life-zone-ecology/oclc/625212 (accessed on 6 February 2024).
- Buchs, D.M.; Baumgartner, P.O.; Baumgartner-Mora, C.; Bandini, A.N.; Jackett, S.-J.; Diserens, M.-O.; Stucki, J. Late Cretaceous to Miocene Seamount Accretion and Mélange Formation in the Osa and Burica Peninsulas (Southern Costa Rica): Episodic Growth of a Convergent Margin. In The Origin and Evolution of the Caribbean Plate; James, K.H., Lorente, M.A., Pindell, J.L., Eds.; Geological Society of London: London, UK, 2009; Volume 328, ISBN 978-1-86239-288-5. [Google Scholar]
- Taylor, P.; Asner, G.; Dahlin, K.; Anderson, C.; Knapp, D.; Martin, R.; Mascaro, J.; Chazdon, R.; Cole, R.; Wanek, W.; et al. Landscape-Scale Controls on Aboveground Forest Carbon Stocks on the Osa Peninsula, Costa Rica. PLoS ONE 2015, 10, e0126748. [Google Scholar] [CrossRef]
- Ley 7575. Available online: http://www.pgrweb.go.cr/scij/Busqueda/Normativa/Normas/nrm_texto_completo.aspx?nValor1=1&nValor2=41661 (accessed on 19 July 2023).
- Huarcaya, R.P.; Morales, M.L.; Álvarez-Alcázar, L.; Whitworth, A. The First Ex-Situ Germination and Dispersal Mechanisms of the Rare, Critically Endangered Tree, Pleodendron Costaricense. Trop. Conserv. Sci. 2022, 15, 1–12. [Google Scholar] [CrossRef]
- Lopez Gutierrez, B.; Almeyda Zambrano, A.M.; Mulder, G.; Ols, C.; Dirzo, R.; Almeyda Zambrano, S.L.; Quispe Gil, C.A.; Cruz Díaz, J.C.; Alvarez, D.; Valdelomar Leon, V.; et al. Ecotourism: The ‘Human Shield’ for Wildlife Conservation in the Osa Peninsula, Costa Rica. J. Ecotourism 2020, 19, 197–216. [Google Scholar] [CrossRef]
- Lorion, C.M.; Kennedy, B.P. Riparian Forest Buffers Mitigate the Effects of Deforestation on Fish Assemblages in Tropical Headwater Streams. Ecol. Appl. 2009, 19, 468–479. [Google Scholar] [CrossRef]
- Sánchez-Azofeifa, G.A.; Harriss, R.C.; Skole, D.L. Deforestation in Costa Rica: A Quantitative Analysis Using Remote Sensing Imagery1. Biotropica 2001, 33, 378–384. [Google Scholar] [CrossRef]
- Roy, D.P.; Kovalskyy, V.; Zhang, H.K.; Vermote, E.F.; Yan, L.; Kumar, S.S.; Egorov, A. Characterization of Landsat-7 to Landsat-8 Reflective Wavelength and Normalized Difference Vegetation Index Continuity. Remote Sens. Environ. 2016, 185, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS; NTRS—NASA Technical Reports Server. 1974. Available online: https://ntrs.nasa.gov/citations/19740022614 (accessed on 6 February 2024).
- Huete, A.; Justice, C.; van Leeuwen, W. MODIS Vegetation Index: Algorithm Theoretical Basis Document; 1999. Available online: https://modis.gsfc.nasa.gov/data/atbd/atbd_mod13.pdf (accessed on 6 February 2024).
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a Two-Band Enhanced Vegetation Index without a Blue Band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Whitworth, A.; Beirne, C.; Rowe, J.; Ross, F.; Acton, C.; Burdekin, O.; Brown, P. The Response of Faunal Biodiversity to an Unmarked Road in the Western Amazon. Biodivers. Conserv. 2015, 24, 1657–1670. [Google Scholar] [CrossRef]
- Algeet-Abarquero, N.; Sánchez-Azofeifa, A.; Bonatti, J.; Marchamalo, M. Land Cover Dynamics in Osa Region, Costa Rica: Secondary Forest Is Here to Stay. Reg. Environ. Chang. 2015, 15, 1461–1472. [Google Scholar] [CrossRef]
- Frazier, A.E.; Kedron, P. Comparing Forest Fragmentation in Eastern U.S. Forests Using Patch-Mosaic and Gradient Surface Models. Ecol. Inform. 2017, 41, 108–115. [Google Scholar] [CrossRef]
- Kupfer, J.A. Landscape Ecology and Biogeography: Rethinking Landscape Metrics in a Post-FRAGSTATS Landscape. Prog. Phys. Geogr. Earth Environ. 2012, 36, 400–420. [Google Scholar] [CrossRef]
- Gustafson, E.J. How Has the State-of-the-Art for Quantification of Landscape Pattern Advanced in the Twenty-First Century? Landsc. Ecol. 2019, 34, 2065–2072. [Google Scholar] [CrossRef]
- Wu, J.; Levin, S.A. A Patch-Based Spatial Modeling Approach: Conceptual Framework and Simulation Scheme. Ecol. Model. 1997, 101, 325–346. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape Ecology: What Is the State of the Science? Annu. Rev. Ecol. Evol. Syst. 2005, 36, 319–344. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Godron, M. Landscape Ecology, 1st ed.; John Wiley & Sons: New York, NY, USA, 1986; ISBN 978-0-471-87037-1. [Google Scholar]
- Wang, Y.; Brandt, M.; Zhao, M.; Xing, K.; Wang, L.; Tong, X.; Xue, F.; Kang, M.; Jiang, Y.; Fensholt, R. Do Afforestation Projects Increase Core Forests? Evidence from the Chinese Loess Plateau. Ecol. Indic. 2020, 117, 106558. [Google Scholar] [CrossRef]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An Open-Source R Tool to Calculate Landscape Metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef]
- Hijmans, R.J.; van Etten, J.; Sumner, M.; Cheng, J.; Baston, D.; Bevan, A.; Bivand, R.; Busetto, L.; Canty, M.; Fasoli, B.; et al. Raster: Geographic Data Analysis and Modeling 2023. Available online: https://cran.r-project.org/web/packages/raster/raster.pdf (accessed on 6 February 2024).
- McGarigal, K.; Cushman, S.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps; University of Massachusetts: Amherst, MA, USA, 2012. [Google Scholar]
- Huete, A.R.; Liu, H.; van Leeuwen, W.J.D. The Use of Vegetation Indices in Forested Regions: Issues of Linearity and Saturation. In Proceedings of the IGARSS’97, 1997 IEEE International Geoscience and Remote Sensing Symposium Proceedings, Remote Sensing—A Scientific Vision for Sustainable Development, Singapore, 3–8 August 1997; Volume 4, pp. 1966–1968. [Google Scholar]
- Vancutsem, C.; Achard, F.; Pekel, J.-F.; Vieilledent, G.; Carboni, S.; Simonetti, D.; Gallego, J.; Aragão, L.E.O.C.; Nasi, R. Long-Term (1990–2019) Monitoring of Forest Cover Changes in the Humid Tropics. Sci. Adv. 2021, 7, eabe1603. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Fahrig, L.; Watling, J.I.; Nowakowski, J.; Tabarelli, M.; Tischendorf, L.; Melo, F.P.L.; Santos, B.A.; Benchimol, M.; Morante-Filho, J.C.; et al. Preserving 40% Forest Cover Is a Valuable and Well-Supported Conservation Guideline: Reply to Banks-Leite et Al. Ecol. Lett. 2021, 24, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Berveglieri, A.; Imai, N.N.; Christovam, L.E.; Galo, M.L.B.T.; Tommaselli, A.M.G.; Honkavaara, E. Analysis of Trends and Changes in the Successional Trajectories of Tropical Forest Using the Landsat NDVI Time Series. Remote Sens. Appl. Soc. Environ. 2021, 24, 100622. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhu, B.; Tan, K. Forest Biomass Carbon Stocks in China over the Past 2 Decades: Estimation Based on Integrated Inventory and Satellite Data. J. Geophys. Res. Biogeosci. 2005, 110, 1–10. [Google Scholar] [CrossRef]
- Flatt, E.; Basto, A.; Pinto, C.; Ortiz, J.; Navarro, K.; Reed, N.; Brumberg, H.; Chaverri, M.H.; Whitworth, A. Arboreal Wildlife Bridges in the Tropical Rainforest of Costa Rica’s Osa Peninsula. Folia Primatol. 2022, 93, 419–435. [Google Scholar] [CrossRef]
- Newcombe, P.B.; Forsyth, A.; Brumberg, H.; Whitworth, A. Climate-Resilient Conservation Strategies for an Endemic Forest Bird, the Black-Cheeked Ant-Tanager. J. Field Ornithol. 2023, 94. [Google Scholar] [CrossRef]
- Arroyo-Mora, J.P.; Sánchez-Azofeifa, G.A.; Rivard, B.; Calvo, J.C.; Janzen, D.H. Dynamics in Landscape Structure and Composition for the Chorotega Region, Costa Rica from 1960 to 2000. Agric. Ecosyst. Environ. 2005, 106, 27–39. [Google Scholar] [CrossRef]
- Ritchie, H. Palm Oil. Our World Data. 2021. Available online: https://ourworldindata.org/palm-oil (accessed on 14 March 2024).
- Vijay, V.; Pimm, S.L.; Jenkins, C.N.; Smith, S.J. The Impacts of Oil Palm on Recent Deforestation and Biodiversity Loss. PLoS ONE 2016, 11, e0159668. [Google Scholar] [CrossRef]
- Kraenzel, M.; Castillo, A.; Moore, T.; Potvin, C. Carbon Storage of Harvest-Age Teak (Tectona Grandis) Plantations, Panama. For. Ecol. Manag. 2003, 173, 213–225. [Google Scholar] [CrossRef]
- Leiva, R.M.B.; Salas, A.C. Spatial Distribution of Mangrove Species and Their Association with the Substrate Sediment Types, Estuarine Sector Térraba-Sierpe National Wetlands, Costa Rica. Rev. Biol. Trop. 2015, 63, 47–60. [Google Scholar] [CrossRef]
- Geldmann, J.; Manica, A.; Burgess, N.D.; Coad, L.; Balmford, A. A Global-Level Assessment of the Effectiveness of Protected Areas at Resisting Anthropogenic Pressures. Proc. Natl. Acad. Sci. USA 2019, 116, 23209–23215. [Google Scholar] [CrossRef] [PubMed]
- Joppa, L.; Pfaff, A. Reassessing the Forest Impacts of Protection. Ann. N. Y. Acad. Sci. 2010, 1185, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Andam, K.S.; Ferraro, P.J.; Pfaff, A.; Sanchez-Azofeifa, G.A.; Robalino, J.A. Measuring the Effectiveness of Protected Area Networks in Reducing Deforestation. Proc. Natl. Acad. Sci. USA 2008, 105, 16089–16094. [Google Scholar] [CrossRef]
- Andam, K.S.; Ferraro, P.J.; Hanauer, M.M. The Effects of Protected Area Systems on Ecosystem Restoration: A Quasi-Experimental Design to Estimate the Impact of Costa Rica’s Protected Area System on Forest Regrowth. Conserv. Lett. 2013, 6, 317–323. [Google Scholar] [CrossRef]
- Fagan, M.E.; DeFries, R.S.; Sesnie, S.E.; Arroyo, J.P.; Walker, W.; Soto, C.; Chazdon, R.L.; Sanchun, A. Land Cover Dynamics Following a Deforestation Ban in Northern Costa Rica. Environ. Res. Lett. 2013, 8, 034017. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Beita, C.M.; Murillo, L.F.S.; Alvarado, L.D.A. Ecological Corridors in Costa Rica: An Evaluation Applying Landscape Structure, Fragmentation-Connectivity Process, and Climate Adaptation. Conserv. Sci. Pract. 2021, 3, e475. [Google Scholar] [CrossRef]
- Temple, S.A.; Cary, J.R. Modeling Dynamics of Habitat-Interior Bird Populations in Fragmented Landscapes. Conserv. Biol. 1988, 2, 340–347. [Google Scholar] [CrossRef]
- Reid, J.L.; Fagan, M.E.; Lucas, J.; Slaughter, J.; Zahawi, R.A. The Ephemerality of Secondary Forests in Southern Costa Rica. Conserv. Lett. 2019, 12, e12607. [Google Scholar] [CrossRef]
- Vargas Soto, J.S.; Beirne, C.; Whitworth, A.; Cruz Diaz, J.C.; Flatt, E.; Pillco-Huarcaya, R.; Olson, E.R.; Azofeifa, A.; Saborío-R, G.; Salom-Pérez, R.; et al. Human Disturbance and Shifts in Vertebrate Community Composition in a Biodiversity Hotspot. Conserv. Biol. 2022, 36, e13813. [Google Scholar] [CrossRef]
- Hansen, A.; di Castri, F. (Eds.) Landscape Boundaries; Ecological Studies; Springer: New York, NY, USA, 1992; Volume 92, ISBN 978-1-4612-7677-7. [Google Scholar]
- Gilroy, J.J.; Prescott, G.W.; Cardenas, J.S.; Castañeda, P.G.D.P.; Sánchez, A.; Rojas-Murcia, L.E.; Medina Uribe, C.A.; Haugaasen, T.; Edwards, D.P. Minimizing the Biodiversity Impact of Neotropical Oil Palm Development. Glob. Chang. Biol. 2015, 21, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Cushman, S.A.; McRae, B.; Adriaensen, F.; Beier, P.; Shirley, M.; Zeller, K. Biological Corridors and Connectivity. In Key Topics in Conservation Biology 2; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 384–404. ISBN 978-1-118-52017-8. [Google Scholar]
- Brownson, K.; Anderson, E.P.; Ferreira, S.; Wenger, S.; Fowler, L.; German, L. Governance of Payments for Ecosystem Ecosystem Services Influences Social and Environmental Outcomes in Costa Rica. Ecol. Econ. 2020, 174, 106659. [Google Scholar] [CrossRef]
- Arriagada, R.A.; Ferraro, P.J.; Sills, E.O.; Pattanayak, S.K.; Cordero-Sancho, S. Do Payments for Environmental Services Affect Forest Cover? A Farm-Level Evaluation from Costa Rica. Land Econ. 2012, 88, 382–399. [Google Scholar] [CrossRef]
- Robalino, J.; Pfaff, A.; Sandoval, C.; Sanchez-Azofeifa, G.A. Can We Increase the Impacts from Payments for Ecosystem Services? Impact Rose over Time in Costa Rica, yet Spatial Variation Indicates More Potential. For. Policy Econ. 2021, 132, 102577. [Google Scholar] [CrossRef]
- Brumberg, H.; Beirne, C.; Broadbent, E.N.; Almeyda Zambrano, A.M.; Almeyda Zambrano, S.L.; Quispe Gil, C.A.; Lopez Gutierrez, B.; Eplee, R.; Whitworth, A. Riparian Buffer Length Is More Influential than Width on River Water Quality: A Case Study in Southern Costa Rica. J. Environ. Manag. 2021, 286, 112132. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.P.; Cochrane, M.A.; Souza, C.M.; Laurance, W.F. Roads, Deforestation, and the Mitigating Effect of Protected Areas in the Amazon. Biol. Conserv. 2014, 177, 203–209. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G.W. Impacts of Roads and Linear Clearings on Tropical Forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef]









| Palm Plantation | Mangrove | Water | Grassland | Exposed Soil/Urban | Mature Forest | Secondary Forest | Wetland | Teak/Gmelina | Natural | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1987 | 2.48% | 5.19% | 4.17% | 16.93% | 9.16% | 27.47% | 32.34% | 2.23% | 0.02% | 71.40% |
| 1998 | 3.78% | 5.3% | 4.28% | 17.37% | 1.37% | 26.26% | 36.90% | 4.07% | 0.66% | 76.81% |
| 2019 | 8.16% | 4.34% | 4.48% | 8.61% | 3.16% | 25.47% | 39.98% | 4.55% | 1.27% | 78.82% |
| Palm Plantation | Mangrove | Water | Grassland | Exposed Soil/Urban | Mature Forest | Secondary Forest | Wetland | Teak/Gmelina | Natural | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1987 | 1.07% | 1.42% | 0.41% | 11.83% | 6.68% | 40.84% | 36.97% | 0.73% | 0.04% | 80.37% |
| 1998 | 1.05% | 1.29% | 0.38% | 12.58% | 0.66% | 39.36% | 42.51% | 1.5% | 0.67% | 85.04% |
| 2019 | 2.37% | 0.69% | 0.54% | 7.43% | 1.86% | 38.51% | 46.27% | 0.96% | 1.36% | 86.97% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brumberg, H.; Furey, S.; Bouffard, M.G.; Mata Quirós, M.J.; Murayama, H.; Neyestani, S.; Pauline, E.; Whitworth, A.; Madden, M. Increasing Forest Cover and Connectivity Both Inside and Outside of Protected Areas in Southwestern Costa Rica. Remote Sens. 2024, 16, 1088. https://doi.org/10.3390/rs16061088
Brumberg H, Furey S, Bouffard MG, Mata Quirós MJ, Murayama H, Neyestani S, Pauline E, Whitworth A, Madden M. Increasing Forest Cover and Connectivity Both Inside and Outside of Protected Areas in Southwestern Costa Rica. Remote Sensing. 2024; 16(6):1088. https://doi.org/10.3390/rs16061088
Chicago/Turabian StyleBrumberg, Hilary, Samuel Furey, Marie G. Bouffard, María José Mata Quirós, Hikari Murayama, Soroush Neyestani, Emily Pauline, Andrew Whitworth, and Marguerite Madden. 2024. "Increasing Forest Cover and Connectivity Both Inside and Outside of Protected Areas in Southwestern Costa Rica" Remote Sensing 16, no. 6: 1088. https://doi.org/10.3390/rs16061088
APA StyleBrumberg, H., Furey, S., Bouffard, M. G., Mata Quirós, M. J., Murayama, H., Neyestani, S., Pauline, E., Whitworth, A., & Madden, M. (2024). Increasing Forest Cover and Connectivity Both Inside and Outside of Protected Areas in Southwestern Costa Rica. Remote Sensing, 16(6), 1088. https://doi.org/10.3390/rs16061088