

Species Abundance Modelling of Arctic-Boreal Zone Ducks Informed by Satellite Remote Sensing
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
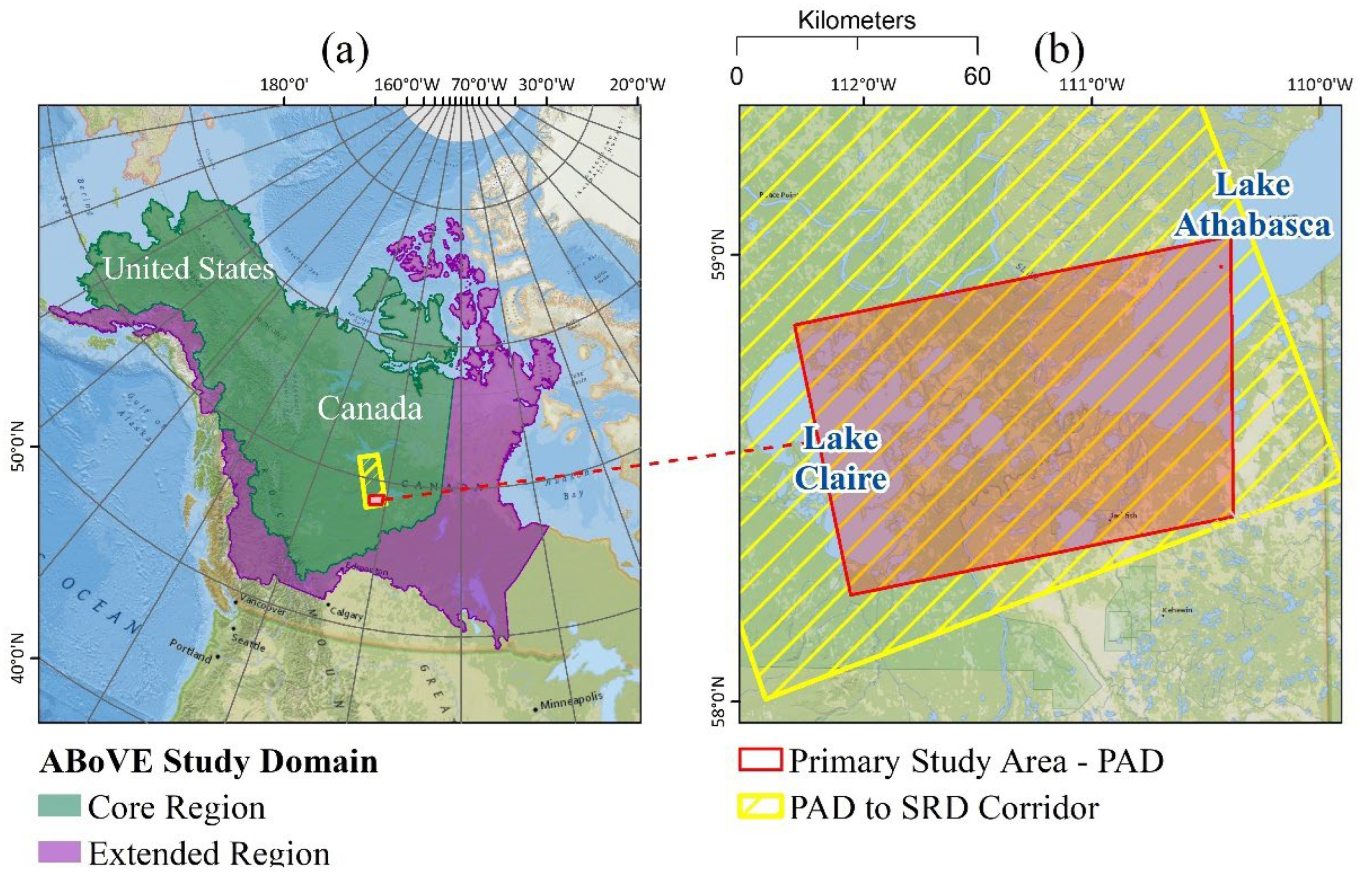
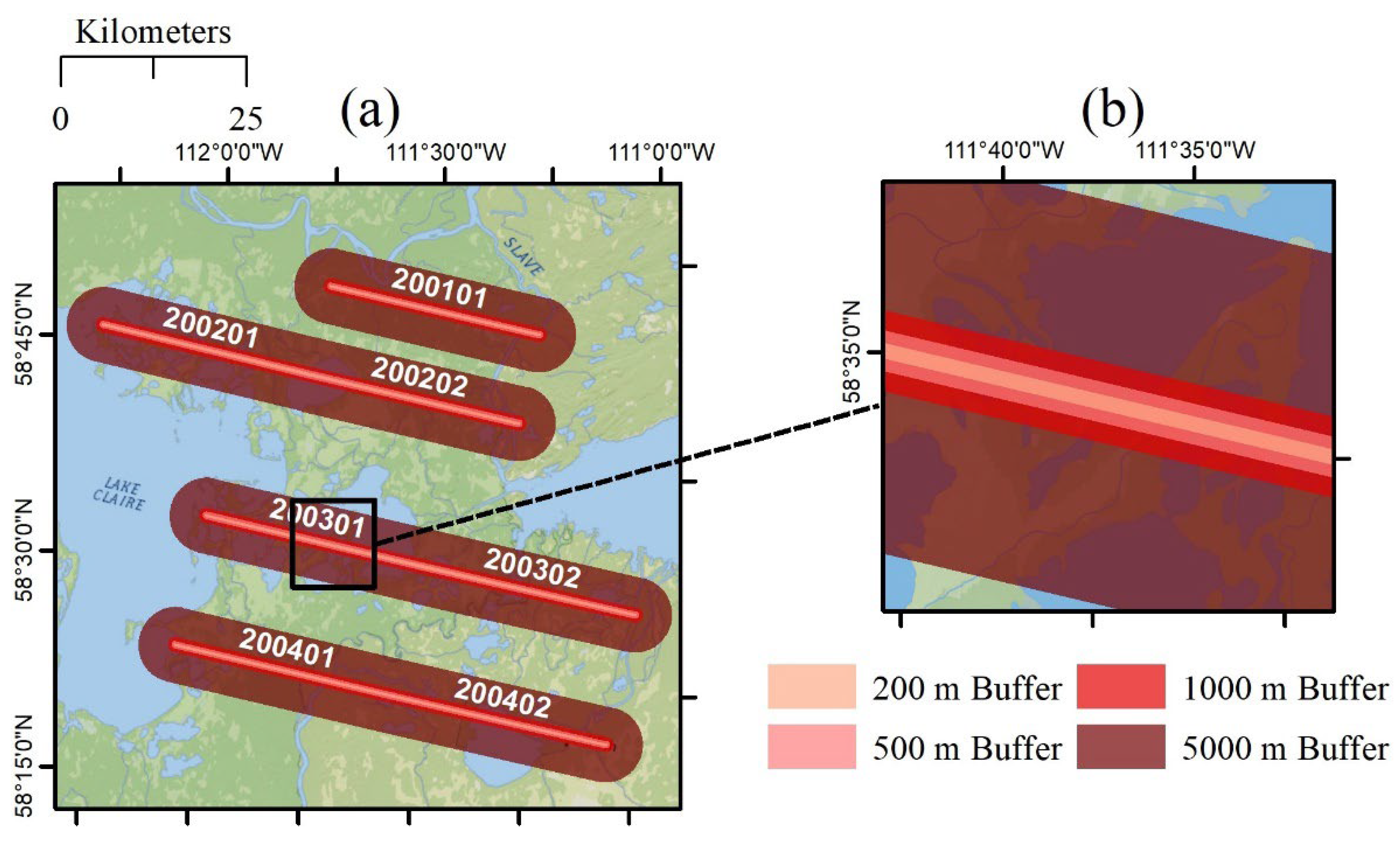
2.1. Study Area
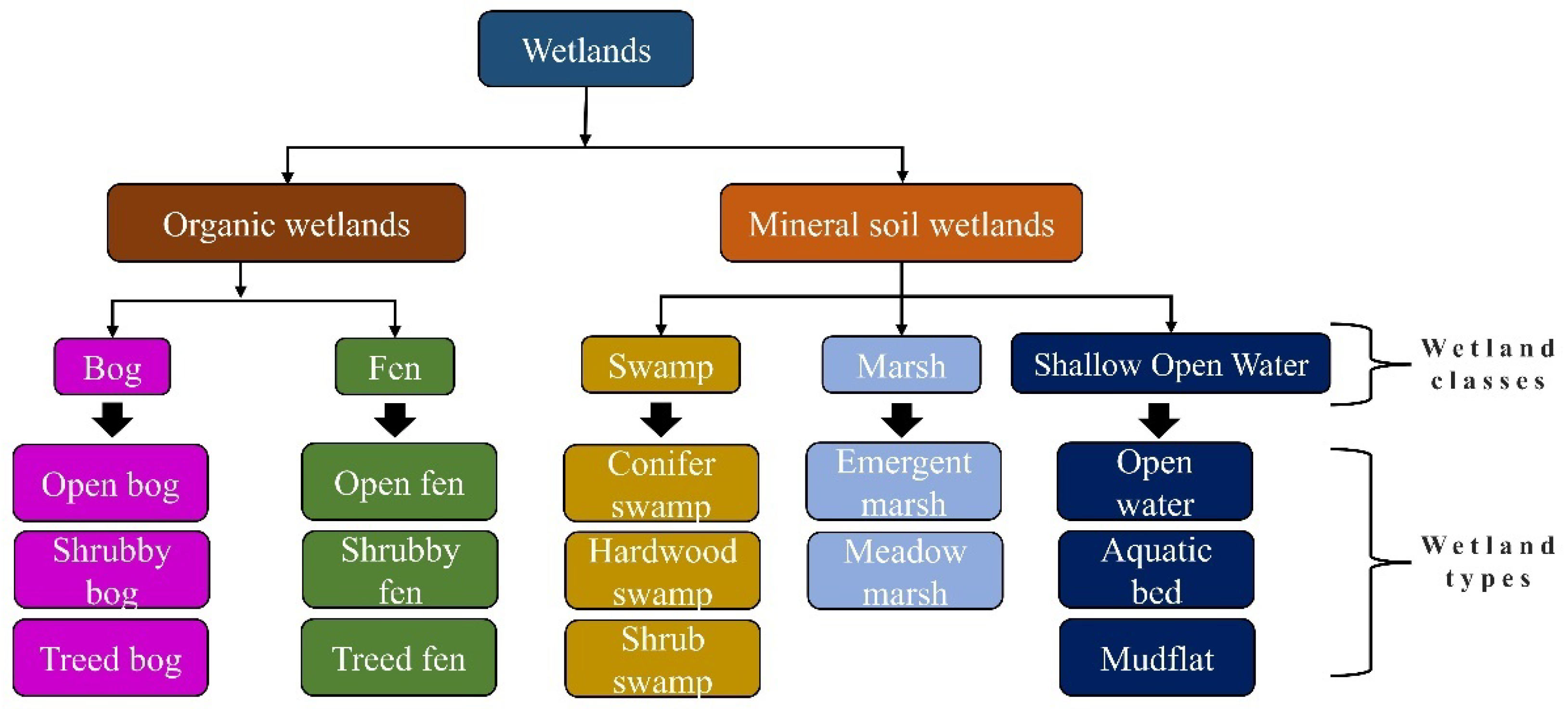
2.2. Wetland Type Mapping
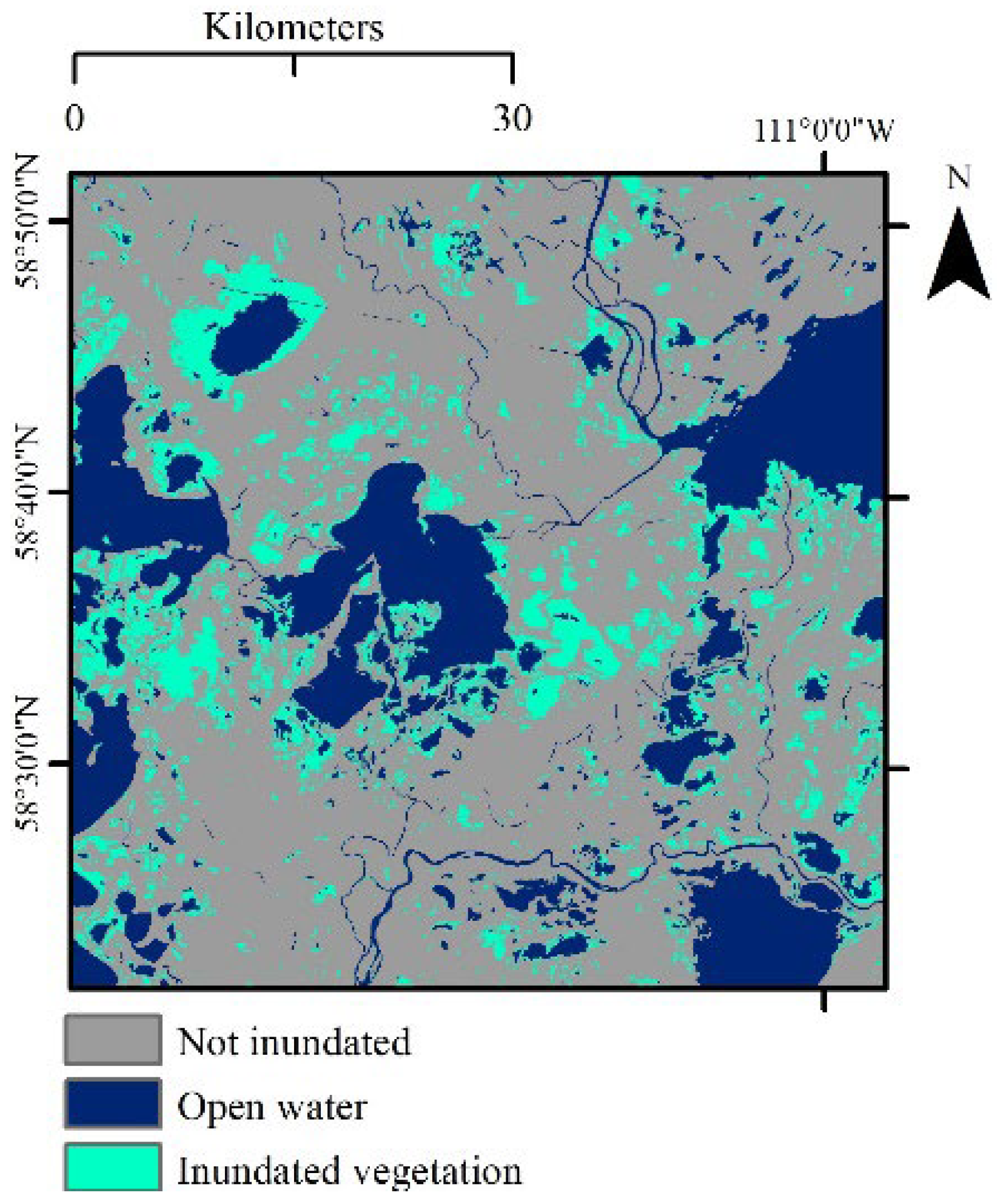
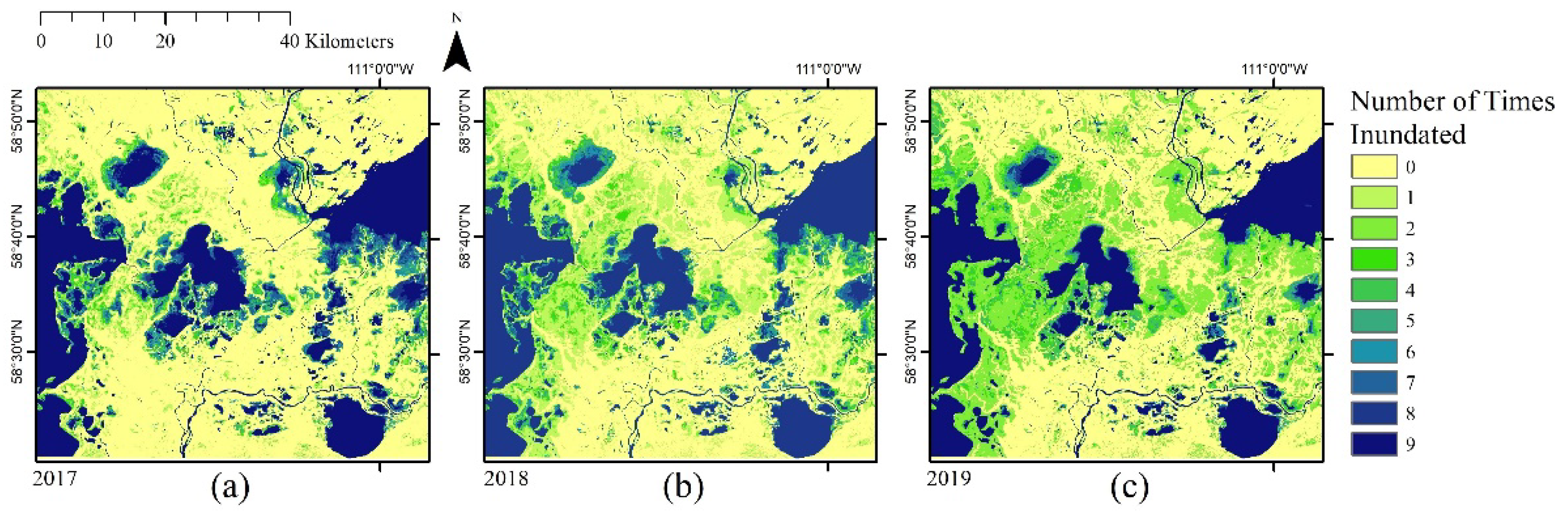
2.3. Wetland Inundation Mapping
2.4. Duck Abundance Modelling
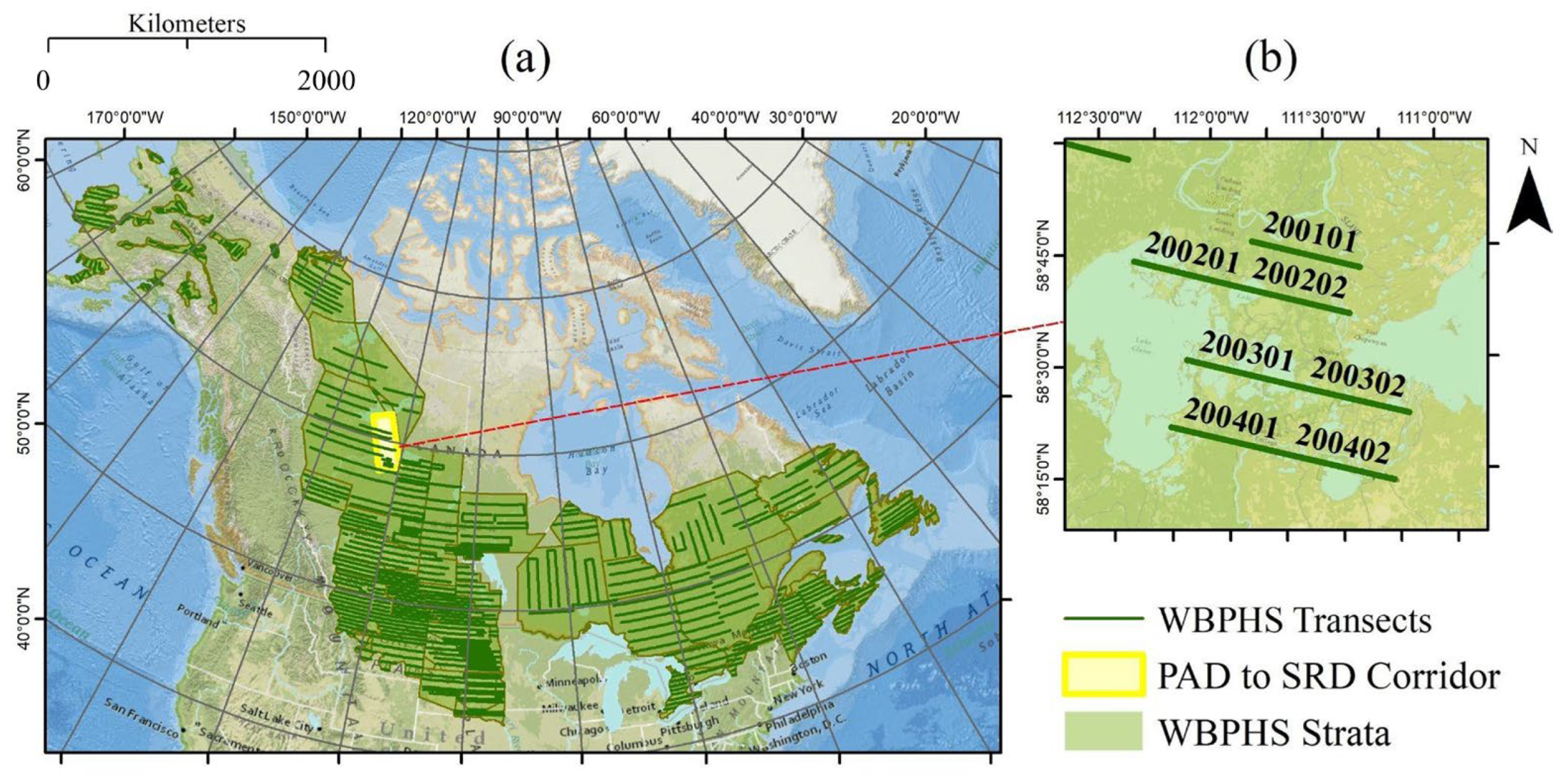
2.4.1. Duck Survey Data from the WBPHS
2.4.2. Calculation of Spatial Statistics
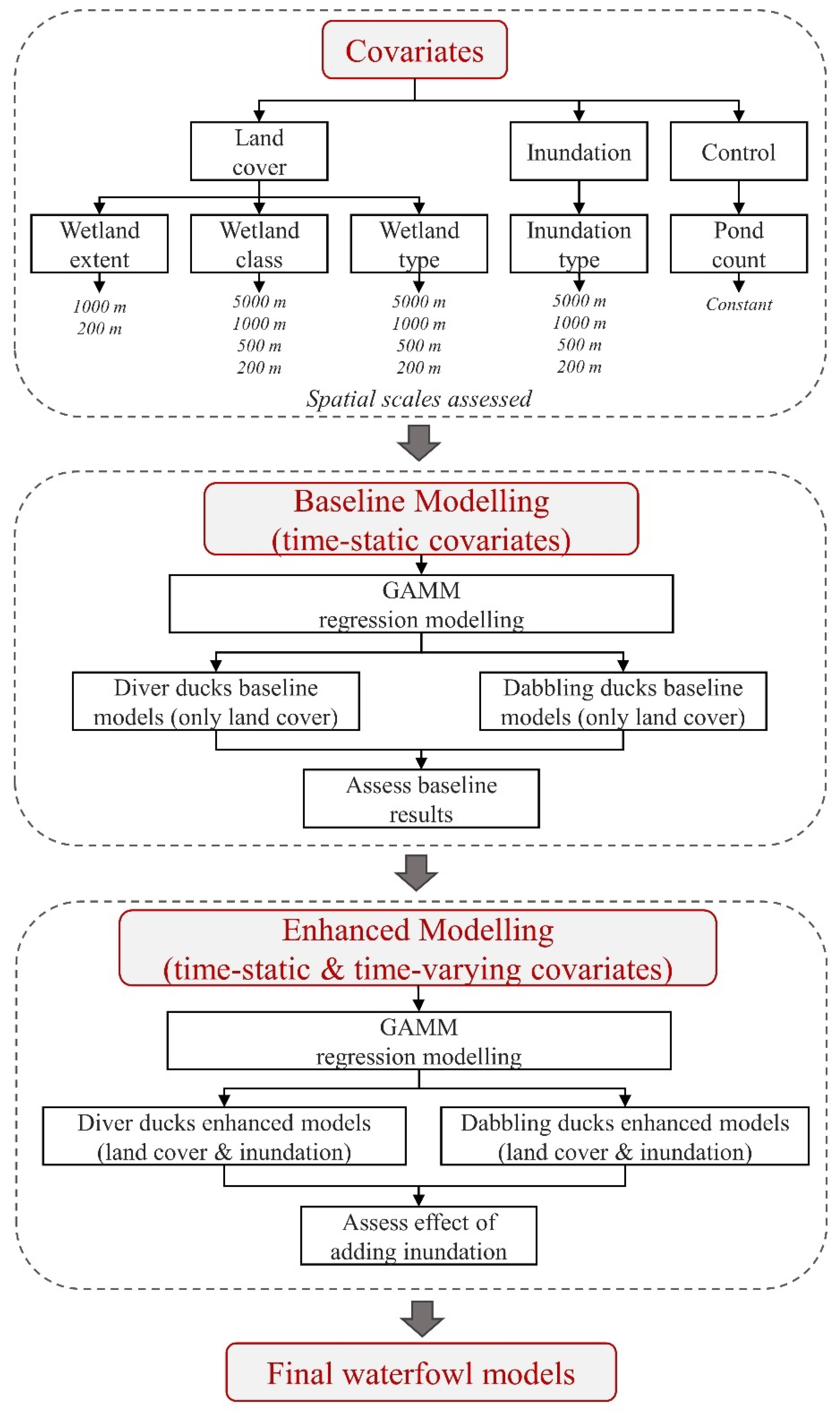
2.4.3. Covariates Used for SAMs
2.4.4. Development of Duck SAMs
3. Results
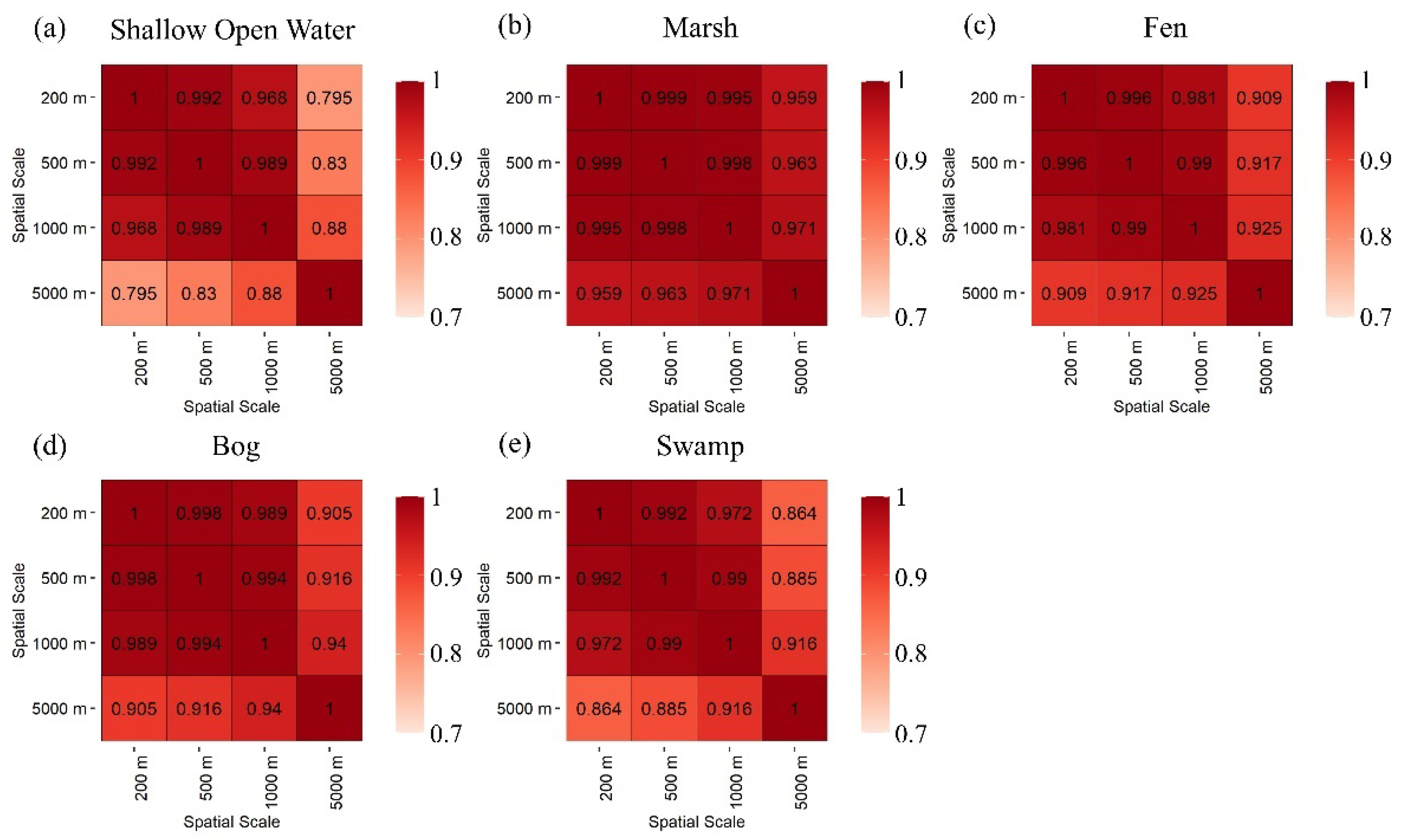
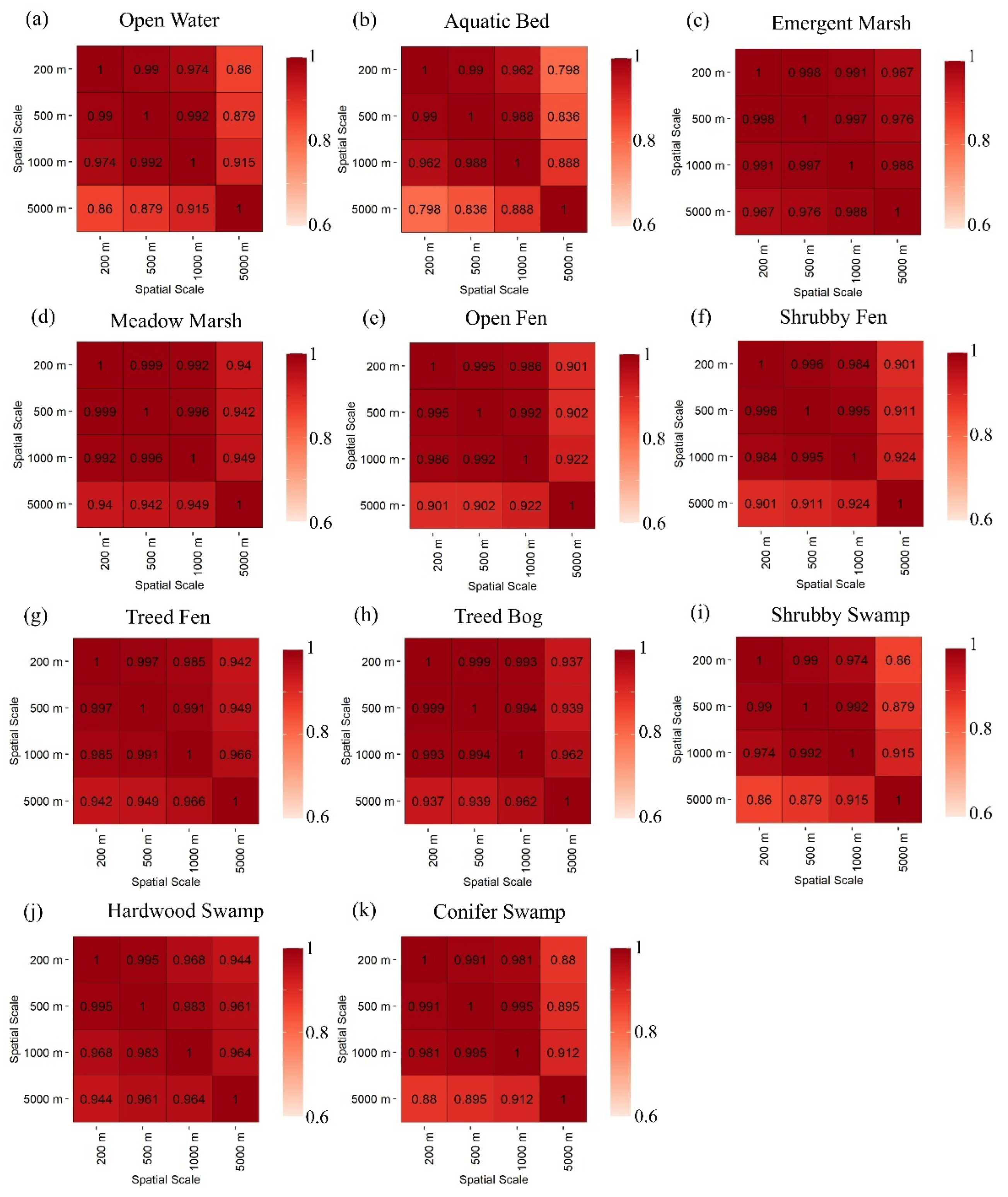
3.1. Pairwise Correlations of Wetland Classes and Types
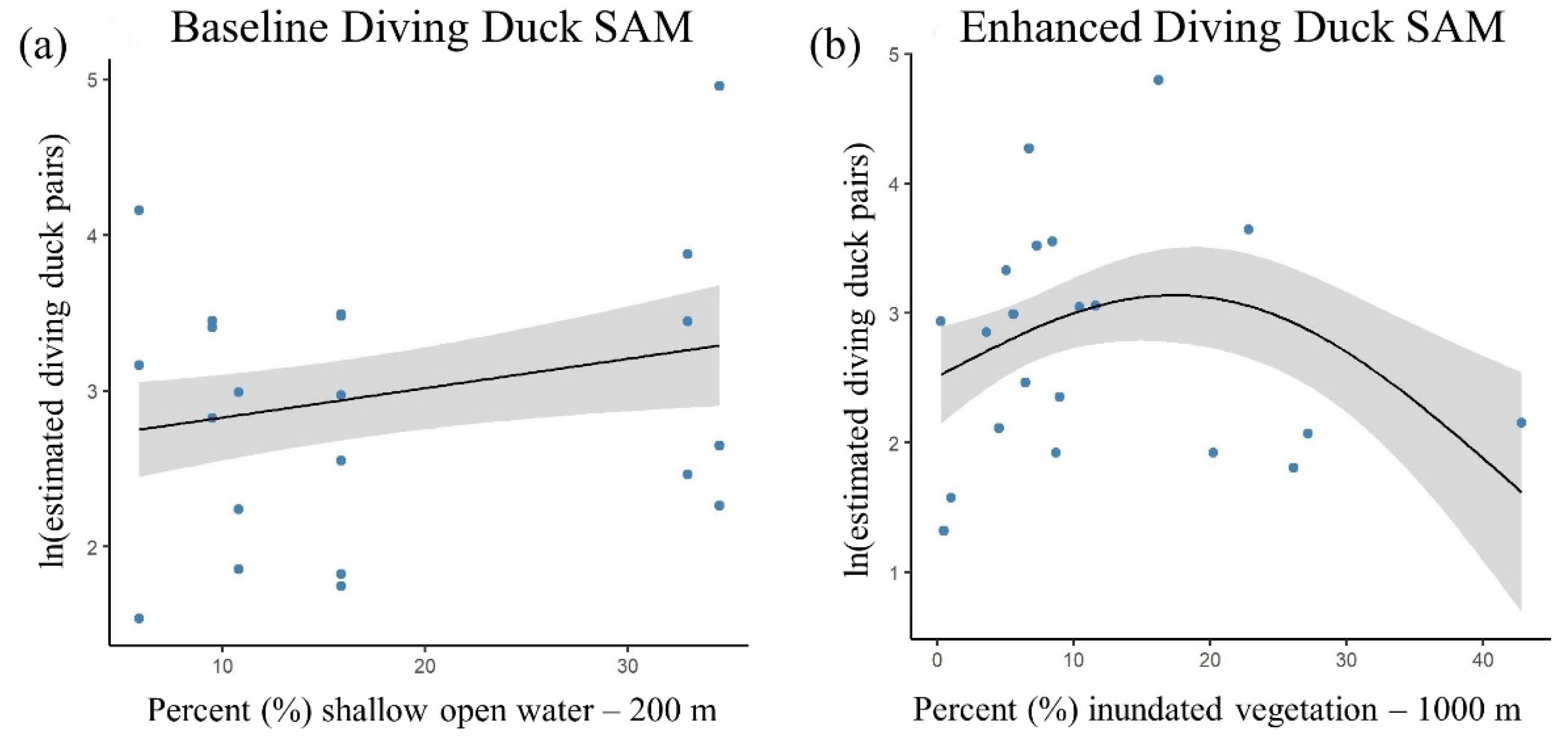
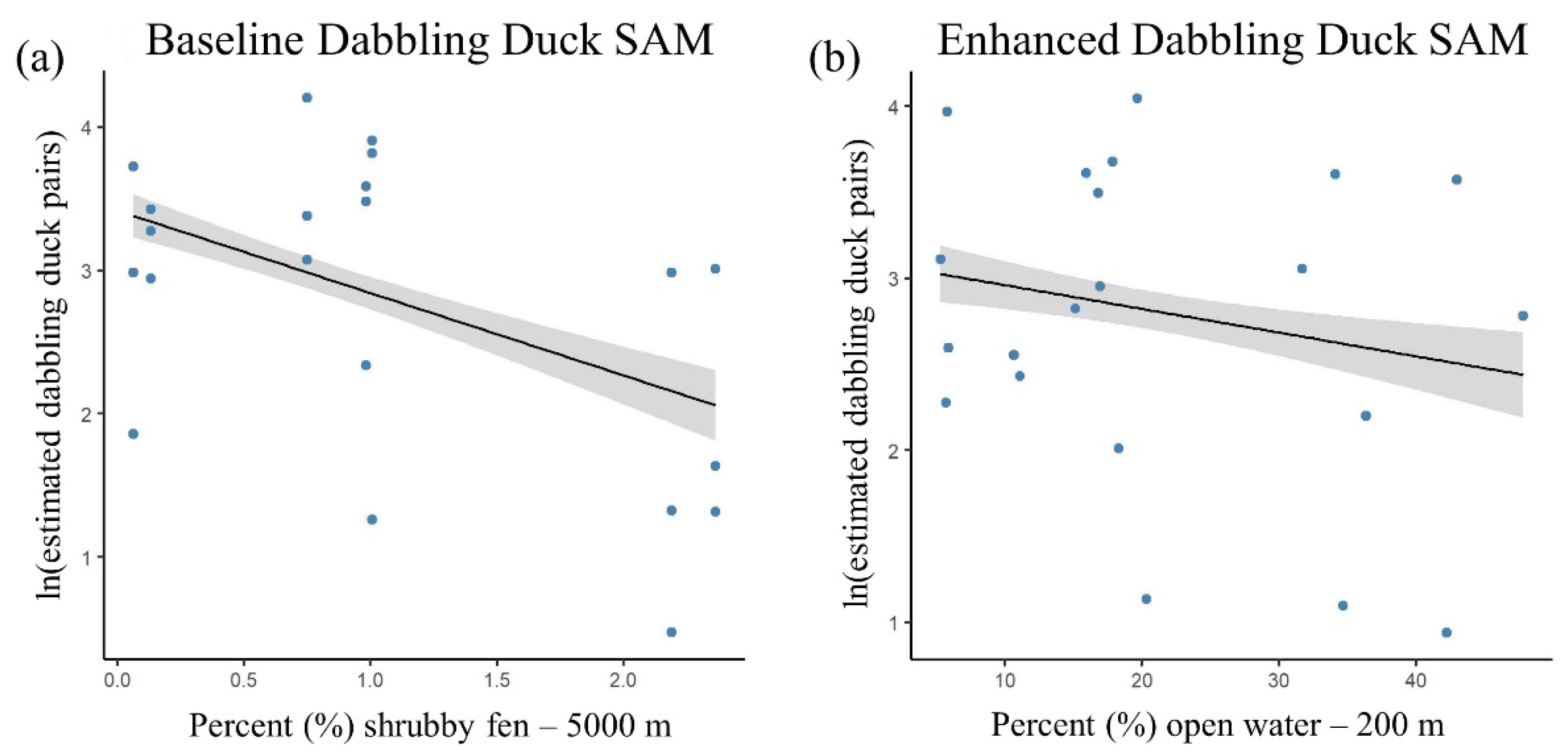
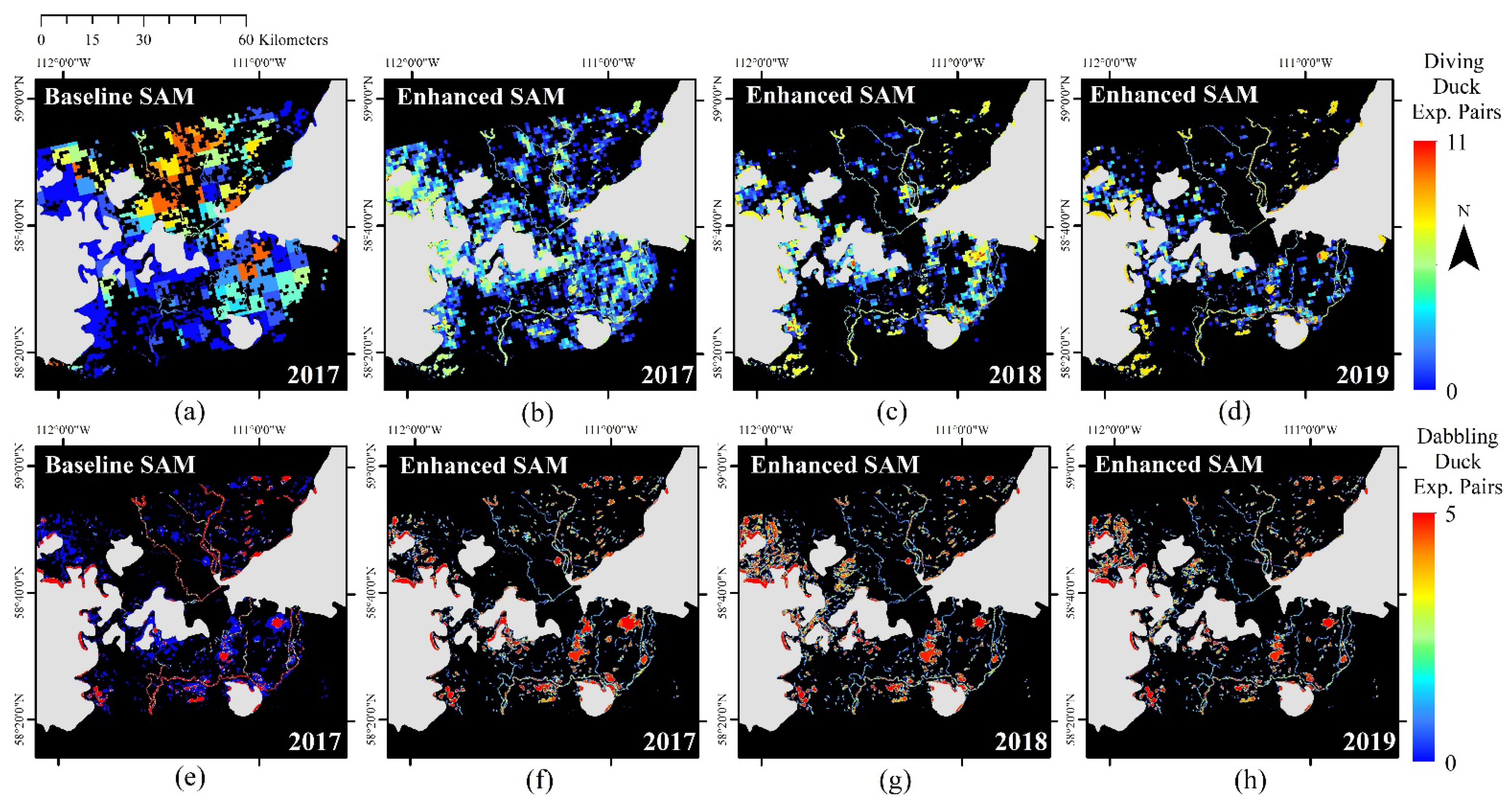
3.2. Baseline Duck SAMs
3.3. Enhanced Duck SAMs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AOI | PALSAR-2 Date 1 | PALSAR-2 Date 2 | Landsat 8 Date 1 | Landsat 8 Date 2 | Sentinel-1 Date Ranges |
| PAD East | 2 July 2018 | 3 July 2017 | 28 May 2019 | 26 August 2017 | 2 June 2017–20 August 2018 |
| PAD West | 17 July 2017 | 27 August 2018 | 28 May 2019 | 26 August 2017 | 2 June 2017–20 August 2018 |
| SRD East | 2 July 2018 | 21 July 2018 | 14 June 2017 | 17 August 2017 | 12 June 2017–20 August 2018 |
| SRD West | 28 June 2018 | 21 July 2018 | 30 May 2018 | 31 August 2017 | 12 June 2017–18 August 2019 |
References
- Olefeldt, D.; Hovemyr, M.; Kuhn, M.A.; Bastviken, D.; Bohn, T.J.; Connolly, J.; Crill, P.; Euskirchen, E.S.; Finkelstein, S.A.; Genet, H.; et al. The Boreal–Arctic Wetland and Lake Dataset (BAWLD). Earth Syst. Sci. Data 2021, 13, 5127–5149. [Google Scholar] [CrossRef]
- Wrona, F.; Reist, J.; Lehtonen, H.; Kahilainen, K.; Forsstrom, L.; Wrona, F.; Reist, J.; Amundsen, P.; Chambers, P.; Christoffersen, K.; et al. Freshwater Ecosystems. In Arctic Biodiversity Assessment: Status and Trends in Arctic Biodiversity; Narayana Press: Odder, Denmark, 2013; pp. 443–485. [Google Scholar]
- Hofgaard, A.; Harper, K.; Golubeva, E. The Role of the Circumarctic Forest–Tundra Ecotone for Arctic Biodiversity. Biodiversity 2012, 13, 174–181. [Google Scholar] [CrossRef]
- Berteaux, D.; Gauthier, G.; Domine, F.; Ims, R.A.; Lamoureux, S.F.; Lévesque, E.; Yoccoz, N. Effects of Changing Permafrost and Snow Conditions on Tundra Wildlife: Critical Places and Times. Arct. Sci. 2017, 3, 65–90. [Google Scholar] [CrossRef]
- Kreplin, H.N.; Santos Ferreira, C.S.; Destouni, G.; Keesstra, S.D.; Salvati, L.; Kalantari, Z. Arctic Wetland System Dynamics under Climate Warming. Wiley Interdiscip. Rev. Water 2021, 8, e1526. [Google Scholar] [CrossRef]
- Previdi, M.; Smith, K.; Polvani, L. Arctic Amplification of Climate Change: A Review of Underlying Mechanisms. Environ. Res. Lett. 2021, 16, 093003. [Google Scholar] [CrossRef]
- Rawlins, M.A.; Steele, M.; Holland, M.M.; Adam, J.C.; Cherry, J.E.; Francis, J.A.; Groisman, P.Y.; Hinzman, L.D.; Huntington, T.G.; Kane, D.L.; et al. Analysis of the Arctic System for Freshwater Cycle Intensification: Observations and Expectations. J. Clim. 2010, 23, 5715–5737. [Google Scholar] [CrossRef]
- Cohen, J.; Screen, J.A.; Furtado, J.C.; Barlow, M.; Whittleston, D.; Coumou, D.; Francis, J.; Dethloff, K.; Entekhabi, D.; Overland, J.; et al. Recent Arctic Amplification and Extreme Mid-Latitude Weather. Nat. Geosci. 2014, 7, 627–637. [Google Scholar] [CrossRef]
- Wrona, F.J.; Johansson, M.; Culp, J.M.; Jenkins, A.; Mård, J.; Myers-Smith, I.H.; Prowse, T.D.; Vincent, W.F.; Wookey, P.A. Transitions in Arctic Ecosystems: Ecological Implications of a Changing Hydrological Regime. J. Geophys. Res. Biogeosci. 2016, 121, 650–674. [Google Scholar] [CrossRef]
- Kåresdotter, E.; Destouni, G.; Ghajarnia, N.; Hugelius, G.; Kalantari, Z. Mapping the Vulnerability of Arctic Wetlands to Global Warming. Earths Future 2021, 9, e2020EF001858. [Google Scholar] [CrossRef]
- Chen, J.; Brissette, F.P.; Poulin, A.; Leconte, R. Overall Uncertainty Study of the Hydrological Impacts of Climate Change for a Canadian Watershed. Water Resour. Res. 2011, 47, W12509. [Google Scholar] [CrossRef]
- Taylor, J.J.; Lawler, J.P.; Aronsson, M.; Barry, T.; Bjorkman, A.D.; Christensen, T.; Coulson, S.J.; Cuyler, C.; Ehrich, D.; Falk, K.; et al. Arctic Terrestrial Biodiversity Status and Trends: A Synopsis of Science Supporting the CBMP State of Arctic Terrestrial Biodiversity Report. Ambio 2020, 49, 833–847. [Google Scholar] [CrossRef] [PubMed]
- Slattery, S.; Morissette, J.; Mack, G.; Butterworth, E. Waterfowl Conservation Planning. In Boreal Birds of North America: A Hemispheric View of Their Conservation Links and Significance; University of California Press: Berkeley, CA, USA, 2011; pp. 23–41. [Google Scholar]
- Holopainen, S.; Arzel, C.; Dessborn, L.; Elmberg, J.; Gunnarsson, G.; Nummi, P.; Pöysä, H.; Sjöberg, K. Habitat Use in Ducks Breeding in Boreal Freshwater Wetlands: A Review. Eur. J. Wildl. Res. 2015, 61, 339–363. [Google Scholar] [CrossRef]
- Kusack, J.; Tozer, D.; Schummer, M.; Hobson, K. Origins of Harvested American Black Ducks: Stable Isotopes Support the Flyover Hypothesis. J. Wildl. Manag. 2023, 87, e22324. [Google Scholar] [CrossRef]
- Runge, M.; Boomer, G. Population Dynamics and Harvest Management of the Continental Northern Pintail Population; Division of Migratory Bird Management, United States Fish and Wildlife Service: Washington, DC, USA, 2005.
- Prosser, D.J.; Ding, C.; Erwin, R.M.; Mundkur, T.; Sullivan, J.D.; Ellis, E.C. Species Distribution Modeling in Regions of High Need and Limited Data: Waterfowl of China. Avian Res. 2018, 9, 7. [Google Scholar] [CrossRef]
- Tsuji, L.; Wilton, M.; Spiegelaar, N.; Oelbermann, M.; Barbeau, C.; Solomon, A.; Tsuji, C.; Liberda, E.; Meldrum, R.; Karagatzides, J. Enhancing Food Security in Subarctic Canada in the Context of Climate Change: The Harmonization of Indigenous Harvesting Pursuits and Agroforestry Activities to Form a Sustainable Import-Substitution Strategy. In Sustainable Solutions for Food Security: Combating Climate Change by Adaptation; Springer: Cham, Switzerland, 2019; pp. 409–435. [Google Scholar]
- Steen, V.A.; Skagen, S.K.; Melcher, C.P. Implications of Climate Change for Wetland-Dependent Birds in the Prairie Pothole Region. Wetlands 2016, 36, 445–459. [Google Scholar] [CrossRef]
- Ballard, D.C.; Jones, O.E.; Janke, A.K. Factors Affecting Wetland Use by Spring Migrating Ducks in the Southern Prairie Pothole Region. J. Wildl. Manag. 2021, 85, 1490–1506. [Google Scholar] [CrossRef]
- Doherty, K.E.; Evans, J.S.; Walker, J.; Devries, J.H.; Howerter, D.W. Building the Foundation for International Conservation Planning for Breeding Ducks across the U.S. and Canadian Border. PLoS ONE 2015, 10, e0116735. [Google Scholar] [CrossRef]
- Smith, W. A Critical Review of the Aerial and Ground Surveys of Breeding Waterfowl in North America; National Biological Service: Laurel, MD, USA, 1995.
- Niemuth, N.D.; Solberg, J.W. Response of Waterbirds to Number of Wetlands in the Prairie Pothole Region of North Dakota, U.S.A. Waterbirds 2003, 26, 233–238. [Google Scholar] [CrossRef]
- Wong, L.; Cornelis Van Kooten, G.; Clarke, J.A. Search the Impact of Agriculture on Waterfowl Abundance: Evidence from Panel Data. J. Agric. Resour. Econ. 2012, 37, 321–334. [Google Scholar]
- Walker, J.; Rotella, J.J.; Stephens, S.E.; Lindberg, M.S.; Ringelman, J.K.; Hunter, C.; Smith, A.J. Time-Lagged Variation in Pond Density and Primary Productivity Affects Duck Nest Survival in the Prairie Pothole Region. Ecol. Appl. 2013, 23, 1061–1074. [Google Scholar] [CrossRef]
- Niemuth, N.D.; Fleming, K.K.; Reynolds, R.E. Waterfowl Conservation in the US Prairie Pothole Region: Confronting the Complexities of Climate Change. PLoS ONE 2014, 9, e100034. [Google Scholar] [CrossRef]
- McIntyre, N.E.; Liu, G.; Gorzo, J.; Wright, C.K.; Guntenspergen, G.R.; Schwartz, F. Simulating the Effects of Climate Variability on Waterbodies and Wetland-Dependent Birds in the Prairie Pothole Region. Ecosphere 2019, 10, e02711. [Google Scholar] [CrossRef]
- Beamish, A.; Raynolds, M.; Epstein, H.; Frost, G.; Macander, M.; Bergstedt, H.; Bartsch, A.; Kruse, S.; Miles, V.; Tanis, C.; et al. Recent Trends and Remaining Challenges for Optical Remote Sensing of Arctic Tundra Vegetation: A Review and Outlook. Remote Sens. Environ. 2020, 246, 111872. [Google Scholar] [CrossRef]
- Yang, D.; Kane, D. Arctic Hydrology, Permafrost and Ecosystems; Springer Nature: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Cooley, S.; Smith, L.; Ryan, J.; Pitcher, L.; Pavelsky, T. Arctic-Boreal Lake Dynamics Revealed Using CubeSat Imagery. Geophys. Res. Lett. 2019, 46, 2111–2120. [Google Scholar] [CrossRef]
- Carroll, M.; Wooten, M.; DiMiceli, C.; Sohlberg, R.; Kelly, M. Quantifying Surface Water Dynamics at 30 Meter Spatial Resolution in the North American High Northern Latitudes 1991–2011. Remote Sens. 2016, 8, 622. [Google Scholar] [CrossRef]
- Schumann, G.; Giustarini, L.; Tarpanelli, A.; Jarihani, B.; Martinis, S. Flood Modeling and Prediction Using Earth Observation Data. Surv. Geophys. 2023, 44, 1553–1578. [Google Scholar] [CrossRef]
- Brisco, B. Mapping and monitoring surface water and wetlands with synthetic aperture radar. In Remote Sensing of Wetlands: Applications and Advances; CRC Press: Boca Raton, FL, USA, 2015; pp. 119–136. [Google Scholar]
- Huang, C.; Smith, L.C.; Kyzivat, E.D.; Fayne, J.V.; Ming, Y.; Spence, C. Tracking Transient Boreal Wetland Inundation with Sentinel-1 SAR: Peace-Athabasca Delta, Alberta and Yukon Flats, Alaska. GIsci Remote Sens. 2022, 59, 1767–1792. [Google Scholar] [CrossRef]
- Barker, N.; Cumming, S.; Darveau, M. Models to Predict the Distribution and Abundance of Breeding Ducks in Canada. Avian Conserv. Ecol. 2014, 9, 7. [Google Scholar] [CrossRef]
- Smith, P.A.; McKinnon, L.; Meltofte, H.; Lanctot, R.B.; Fox, A.D.; Leafloor, J.O.; Soloviev, M.; Franke, A.; Falk, K.; Golovatin, M.; et al. Status and Trends of Tundra Birds across the Circumpolar Arctic. Ambio 2020, 49, 732–748. [Google Scholar] [CrossRef]
- Adde, A.; Darveau, M.; Barker, N.; Cumming, S. Predicting Spatiotemporal Abundance of Breeding Waterfowl across Canada: A Bayesian Hierarchical Modelling Approach. Divers. Distrib. 2020, 26, 1248–1263. [Google Scholar] [CrossRef]
- Singer, H.V.; Slattery, S.M.; Armstrong, L.; Witherly, S. Assessing Breeding Duck Population Trends Relative to Anthropogenic Disturbances across the Boreal Plains of Canada, 1960–2007. Avian Conserv. Ecol. 2020, 15, 1. [Google Scholar] [CrossRef]
- Adde, A.; Casabona i Amat, C.; Mazerolle, M.J.; Darveau, M.; Cumming, S.G.; O’Hara, R.B. Integrated Modeling of Waterfowl Distribution in Western Canada Using Aerial Survey and Citizen Science (EBird) Data. Ecosphere 2021, 12, e03790. [Google Scholar] [CrossRef]
- Smith, K.; Smith, C.; Forest, S.; Richard, A. A Field Guide to the Wetlands of the Boreal Plains Ecozone of Canada; Ducks Unlimited Canada, Western Boreal Office: Edmonton, AB, Canada, 2007.
- Merchant, M.A.; Warren, R.K.; Edwards, R.; Kenyon, J.K. An Object-Based Assessment of Multi-Wavelength SAR, Optical Imagery and Topographical Datasets for Operational Wetland Mapping in Boreal Yukon, Canada. Can. J. Remote Sens. 2019, 45, 308–332. [Google Scholar] [CrossRef]
- Merchant, M.; Haas, C.; Schroder, J.; Warren, R.; Edwards, R. High-Latitude Wetland Mapping Using Multidate and Multisensor Earth Observation Data: A Case Study in the Northwest Territories. J. Appl. Remote Sens. 2020, 14, 034511. [Google Scholar] [CrossRef]
- Wright, J.R.; Johnson, J.A.; Bayne, E.; Powell, L.L.; Foss, C.R.; Kennedy, J.C.; Marra, P.P. Migratory Connectivity and Annual Cycle Phenology of Rusty Blackbirds (Euphagus Carolinus) Revealed through Archival Gps Tags. Avian Conserv. Ecol. 2021, 16, 20. [Google Scholar] [CrossRef]
- Dyson, M.E.; Slattery, S.M.; Fedy, B.C. Multiscale Nest-Site Selection of Ducks in the Western Boreal Forest of Alberta. Ecol. Evol. 2022, 12, e9139. [Google Scholar] [CrossRef]
- Kahara, S.N.; Skalos, D.; Madurapperuma, B.; Hernandez, K. Habitat Quality and Drought Effects on Breeding Mallard and Other Waterfowl Populations in California, USA. J. Wildl. Manag. 2022, 86, e22133. [Google Scholar] [CrossRef]
- Miller, C.E.; Griffith, P.C.; Goetz, S.J.; Hoy, E.E.; Pinto, N.; McCubbin, I.B.; Thorpe, A.K.; Hofton, M.; Hodkinson, D.; Hansen, C.; et al. An Overview of above Airborne Campaign Data Acquisitions and Science Opportunities. Environ. Res. Lett. 2019, 14, 080201. [Google Scholar] [CrossRef]
- Bourgeau-Chavez, L.; Endres, S.; Powell, R.; Battaglia, M.; Benscoter, B.; Turetsky, M.; Kasischke, E.; Banda, E. Mapping Boreal Peatland Ecosystem Types from Multitemporal Radar and Optical Satellite Imagery. Can. J. For. Res. 2017, 47, 545–559. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Martins, V.S.; Kaleita, A.L.; Gelder, B.K.; Nagel, G.W.; Maciel, D.A. Deep Neural Network for Complex Open-Water Wetland Mapping Using High-Resolution WorldView-3 and Airborne LiDAR Data. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102215. [Google Scholar] [CrossRef]
- Behnamian, A.; Banks, S.; White, L.; Brisco, B.; Milard, K.; Pasher, J.; Chen, Z.; Duffe, J.; Bourgeau-Chavez, L.; Battaglia, M. Semi-Automated Surfacewater Detection with Synthetic Aperture Radar Data: A Wetland Case Study. Remote Sens. 2017, 9, 1209. [Google Scholar] [CrossRef]
- Battaglia, M.J.; Banks, S.; Behnamian, A.; Bourgeau-Chavez, L.; Brisco, B.; Corcoran, J.; Chen, Z.; Huberty, B.; Klassen, J.; Knight, J.; et al. Multi-Source Eo for Dynamic Wetland Mapping and Monitoring in the Great Lakes Basin. Remote Sens. 2021, 13, 599. [Google Scholar] [CrossRef]
- Achanta, R.; Susstrunk, S. Superpixels and Polygons Using Simple Non-Iterative Clustering. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Honolulu, HI, USA, 21–26 July 2017; pp. 4651–4660. [Google Scholar]
- Kemink, K.M.; Adams, V.M.; Pressey, R.L. Integrating Dynamic Processes into Waterfowl Conservation Prioritization Tools. Divers. Distrib. 2021, 27, 585–601. [Google Scholar] [CrossRef]
- Tozer, D.C.; Stewart, R.L.M.; Steele, O.; Gloutney, M. Species-Habitat Relationships and Priority Areas for Marsh-Breeding Birds in Ontario. J. Wildl. Manag. 2020, 84, 786–801. [Google Scholar] [CrossRef]
- Jones, N. Hybrid Wetland Layer (Version 2.1) User Guide; Ducks Unlimited Canada: Edmonton, AB, Canada, 2010.
- Wood, S. Mixed GAM Computation Vehicle with Automatic Smoothness Estimation; Version 1.8-40; RStudio: Vienna, Austria, 2022. [Google Scholar]
- Wen, L.; Rogers, K.; Saintilan, N.; Ling, J. The Influences of Climate and Hydrology on Population Dynamics of Waterbirds in the Lower Murrumbidgee River Floodplains in Southeast Australia: Implications for Environmental Water Management. Ecol. Model. 2011, 222, 154–163. [Google Scholar] [CrossRef]
- Nykänen, M.; Pöysä, H.; Hakkarainen, S.; Rajala, T.; Matala, J.; Kunnasranta, M. Seasonal and Diel Activity Patterns of the Endangered Taiga Bean Goose (Anser fabalis fabalis) during the Breeding Season, Monitored with Camera Traps. PLoS ONE 2021, 16, e0254254. [Google Scholar] [CrossRef]
- Cavanaugh, J.E.; Neath, A.A. The Akaike Information Criterion: Background, Derivation, Properties, Application, Interpretation, and Refinements. Wiley Interdiscip. Rev. Comput. Stat. 2019, 11, e1460. [Google Scholar] [CrossRef]
- Roberts, N.J.; Krilov, L.R. The Continued Threat of Influenza A Viruses. Viruses 2022, 14, 883. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive Effects of Habitat Modification and Species Invasion on Native Species Decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef]
- Venier, L.A.; Thompson, I.D.; Fleming, R.; Malcolm, J.; Aubin, I.; Trofymow, J.A.; Langor, D.; Sturrock, R.; Patry, C.; Outerbridge, R.O.; et al. Effects of Natural Resource Development on the Terrestrial Biodiversity of Canadian Boreal Forests1. Environ. Rev. 2014, 22, 457–490. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural Disturbance Impacts on Ecosystem Services and Biodiversity in Temperate and Boreal Forests. Biol. Rev. Camb. Philos. Soc. 2016, 91, 760–781. [Google Scholar] [CrossRef]
- Webster, K.L.; Beall, F.D.; Creed, I.F.; Kreutzweiser, D.P. Impacts and Prognosis of Natural Resource Development on Water and Wetlands in Canada’s Boreal Zone. Environ. Rev. 2015, 23, 78–131. [Google Scholar] [CrossRef]
- Volik, O.; Elmes, M.; Petrone, R.; Kessel, E.; Green, A.; Cobbaert, D.; Price, J. Wetlands in the Athabasca Oil Sands Region: The Nexus between Wetland Hydrological Function and Resource Extraction. Environ. Rev. 2020, 28, 246–261. [Google Scholar] [CrossRef]
- Petrescu, A.M.R.; Van Beek, L.P.H.; Van Huissteden, J.; Prigent, C.; Sachs, T.; Corradi, C.A.R.; Parmentier, F.J.W.; Dolman, A.J. Modeling Regional to Global CH4 Emissions of Boreal and Arctic Wetlands. Glob. Biogeochem. Cycles 2010, 24, GB4009. [Google Scholar] [CrossRef]
- Grosse, G.; Harden, J.; Turetsky, M.; McGuire, A.D.; Camill, P.; Tarnocai, C.; Frolking, S.; Schuur, E.A.G.; Jorgenson, T.; Marchenko, S.; et al. Vulnerability of High-Latitude Soil Organic Carbon in North America to Disturbance. J. Geophys. Res. Biogeosci 2011, 116, G4. [Google Scholar] [CrossRef]
- Zhang, Z.; Poulter, B.; Feldman, A.F.; Ying, Q.; Ciais, P.; Peng, S.; Li, X. Recent Intensification of Wetland Methane Feedback. Nat. Clim. Chang. 2023, 13, 430–433. [Google Scholar] [CrossRef]
- Watts, J.D.; Kimball, J.S.; Bartsch, A.; McDonald, K.C. Surface Water Inundation in the Boreal-Arctic: Potential Impacts on Regional Methane Emissions. Environ. Res. Lett. 2014, 9, 075001. [Google Scholar] [CrossRef]
- Chasmer, L.; Mahoney, C.; Millard, K.; Nelson, K.; Peters, D.; Merchant, M.; Hopkinson, C.; Brisco, B.; Niemann, O.; Montgomery, J.; et al. Remote Sensing of Boreal Wetlands 2: Methods for Evaluating Boreal Wetland Ecosystem State and Drivers of Change. Remote Sens. 2020, 12, 1321. [Google Scholar] [CrossRef]
- Adeli, S.; Salehi, B.; Mahdianpari, M.; Quackenbush, L.J.; Brisco, B.; Tamiminia, H.; Shaw, S. Wetland Monitoring Using SAR Data: A Meta-Analysis and Comprehensive Review. Remote Sens. 2020, 12, 2190. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Watts, J.D.; Kimball, J.S.; Jones, L.A.; Schroeder, R.; McDonald, K.C. Satellite Microwave Remote Sensing of Contrasting Surface Water Inundation Changes within the Arctic-Boreal Region. Remote Sens. Environ. 2012, 127, 223–236. [Google Scholar] [CrossRef]
- Mitsch, W.; Gosselink, J. Wetlands; John Wiley & Sons: Hoboken, NY, USA, 2015. [Google Scholar]
- Mitsch, W.J.; Hernandez, M.E. Landscape and Climate Change Threats to Wetlands of North and Central America. Aquat. Sci. 2013, 75, 133–149. [Google Scholar] [CrossRef]
- Canisius, F.; Brisco, B.; Murnaghan, K.; Van Der Kooij, M.; Keizer, E. SAR Backscatter and InSAR Coherence for Monitoring Wetland Extent, Flood Pulse and Vegetation: A Study of the Amazon Lowland. Remote Sens. 2019, 11, 720. [Google Scholar] [CrossRef]
- White, L.; Brisco, B.; Dabboor, M.; Schmitt, A.; Pratt, A. A Collection of SAR Methodologies for Monitoring Wetlands. Remote Sens. 2015, 7, 7615–7645. [Google Scholar] [CrossRef]
- Austin, J.; Slattery, S.; Clark, R. Waterfowl Populations of Conservation Concern: Learning from Diverse Challenges, Models and Conservation Strategies. Wildfowl 2014, 470–497. [Google Scholar]
- Hagy, H.; Yaich, S.; Simpson, J.; Carrera, E.; Haukos, D.; Johnson, W.; Loesch, C.; Reid, F.; Stephens, S.; Tiner, R.; et al. Wetland Issues Affecting Waterfowl Conservation in North America. Wildfowl 2014, SI4, 343–367. [Google Scholar]
- Prairie Habitat Joint Venture. Prairie Habitat Joint Venture Implementation Plan 2021–2025: The Prairie Parklands. Report of the Prairie Habitat Joint Venture; Environment Canada: Edmonton, AB, Canada, 2021.












| Total Pair Counts | ||
|---|---|---|
| Survey Year | Diving Ducks | Dabbling Ducks |
| 2017 | 102 | 107 |
| 2018 | 155 | 117 |
| 2019 | 124 | 151 |
| Predictor Variables and Description | Source | Layer Type | Scales | Spatiotemporal Characterization | Application |
|---|---|---|---|---|---|
| open water, aquatic bed, emergent marsh, meadow marsh, open fen, shrubby fen, treed fen, tree bog, shrub swamp, hardwood swamp, conifer swamp | This study | Wetland type | 200 m, 500 m, 1000 m, 5000 m | Spatially varying and time invariant | Baseline and enhanced modelling |
| shallow open water, marsh, fen, bog, swamp | This study | Wetland class | 200 m, 500 m, 1000 m, 5000 m | ||
| Spatially varying and time invariant | Baseline and enhanced modelling | ||||
| open water, wetland | HWL | Wetland extent | 200 m, 1000 m | Spatially varying and time invariant | Baseline and enhanced modelling |
| open water, inundated vegetation, not inundated | This study | Inundation | 200 m, 500 m, 1000 m, 5000 m | Spatially and time varying | Enhanced modelling |
| Model Rank | Covariate | Total EDF 1 | ΔAIC 2 |
|---|---|---|---|
| 1 | s (wetland extent—open water—200 m) | 4.00 | 0.00 |
| 2 | * s (wetland type—open water—200 m) | 4.00 | 1.04 |
| 3 | s (wetland extent—open water—1000 m) | 4.00 | 1.06 |
| 4 | s (wetland type—open water—500 m) | 4.00 | 1.75 |
| 5 | * s (wetland class—shallow open water—200 m) | 4.57 | 2.73 |
| 6 | s (wetland type—hardwood swamp—500 m) | 4.52 | 2.75 |
| 7 | s (wetland type—open water—1000 m) | 4.53 | 3.06 |
| 8 | s (wetland class—shallow open water—500 m) | 4.92 | 3.32 |
| 9 | s (wetland class—swamp—200 m) | 4.39 | 3.66 |
| 10 | s (wetland class—shallow open water—1000 m) | 5.25 | 3.89 |
| - | Null 3 | 7.69 | 7.47 |
| Model Rank | Covariate | Total EDF 1 | ΔAIC 2 |
|---|---|---|---|
| 1 | * s (wetland type—shrubby fen—5000 m) | 7.86 | 0.00 |
| 2 | s (wetland type—shrub swamp—500 m) | 8.73 | 0.45 |
| 3 | s (wetland class—swamp—200 m) | 8.75 | 0.46 |
| 4 | s (wetland class—swamp—1000 m) | 8.75 | 0.46 |
| 5 | s (wetland class—swamp—500 m) | 8.76 | 0.47 |
| 6 | s (wetland type—shrub swamp—200 m) | 8.75 | 0.49 |
| 7 | s (wetland type—shrub swamp—1000 m) | 8.76 | 0.51 |
| 8 | s (wetland type—emergent marsh—200 m) | 8.77 | 0.51 |
| 9 | s (wetland type—emergent marsh—500 m) | 8.78 | 0.52 |
| 10 | * s (wetland type—conifer swamp—5000 m) | 8.59 | 0.52 |
| - | Null 3 | 8.71 | 0.61 |
| Model Rank | Covariate | Total EDF 1 | AIC 2 | ΔAIC 3 | Deviance Explained |
|---|---|---|---|---|---|
| Enhanced | wetland class—shallow open water—200 m + s (inundation—inundated vegetation—1000 m) | 8.16 | 140.57 | 0.00 | 68.9% |
| - | s (inundation—inundated vegetation—1000 m) | 9.86 | 140.97 | 0.40 | 74.0% |
| Enhanced | wetland type—open water—200 m + s (inundation—inundated vegetation—1000 m) | 8.59 | 141.72 | 1.15 | 68.7% |
| Baseline | wetland type—open water—200 m | 4.00 | 146.15 | 5.58 | 35.7% |
| Baseline | wetland class—shallow open water—200 m | 4.67 | 148.03 | 7.46 | 34.2% |
| Model Rank | Covariate | Total EDF 1 | AIC 2 | ΔAIC 3 | Deviance Explained |
|---|---|---|---|---|---|
| Enhanced | wetland type—shrubby fen—5000 m + | 5.00 | 122.43 | 0.00 | 82.7 |
| Enhanced | s (inundation—open water—200 m) | 5.00 | 122.95 | 0.52 | 82.1 |
| Baseline | wetland type—conifer swamp—5000 m + | 7.86 | 127.04 | 4.61 | 83.9 |
| Baseline | s (inundation—open water—500 m) | 8.59 | 127.56 | 5.13 | 84.9 |
| - | s (wetland type—shrubby fen—5000 m) | 9.43 | 128.59 | 6.16 | 85.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merchant, M.A.; Battaglia, M.J.; French, N.; Smith, K.; Singer, H.V.; Armstrong, L.; Harriman, V.B.; Slattery, S. Species Abundance Modelling of Arctic-Boreal Zone Ducks Informed by Satellite Remote Sensing. Remote Sens. 2024, 16, 1175. https://doi.org/10.3390/rs16071175
Merchant MA, Battaglia MJ, French N, Smith K, Singer HV, Armstrong L, Harriman VB, Slattery S. Species Abundance Modelling of Arctic-Boreal Zone Ducks Informed by Satellite Remote Sensing. Remote Sensing. 2024; 16(7):1175. https://doi.org/10.3390/rs16071175
Chicago/Turabian StyleMerchant, Michael Allan, Michael J. Battaglia, Nancy French, Kevin Smith, Howard V. Singer, Llwellyn Armstrong, Vanessa B. Harriman, and Stuart Slattery. 2024. "Species Abundance Modelling of Arctic-Boreal Zone Ducks Informed by Satellite Remote Sensing" Remote Sensing 16, no. 7: 1175. https://doi.org/10.3390/rs16071175
APA StyleMerchant, M. A., Battaglia, M. J., French, N., Smith, K., Singer, H. V., Armstrong, L., Harriman, V. B., & Slattery, S. (2024). Species Abundance Modelling of Arctic-Boreal Zone Ducks Informed by Satellite Remote Sensing. Remote Sensing, 16(7), 1175. https://doi.org/10.3390/rs16071175