
Sensitivity of Sentinel-1 Backscatter to Management-Related Disturbances in Temperate Forests

 , , , ,
, , , ,
Abstract
:1. Introduction
- The sensitivity of C-band backscatter to species-specific seasonality and canopy cover;
- The effect of radar shadow and radar layover on canopy cover loss-related C-band backscatter change;
- The sensitivity of C-band backscatter to disturbance severity for different monitoring scenarios (ascending/descending orbits and VV/VH polarization).
2. Study Area and Data
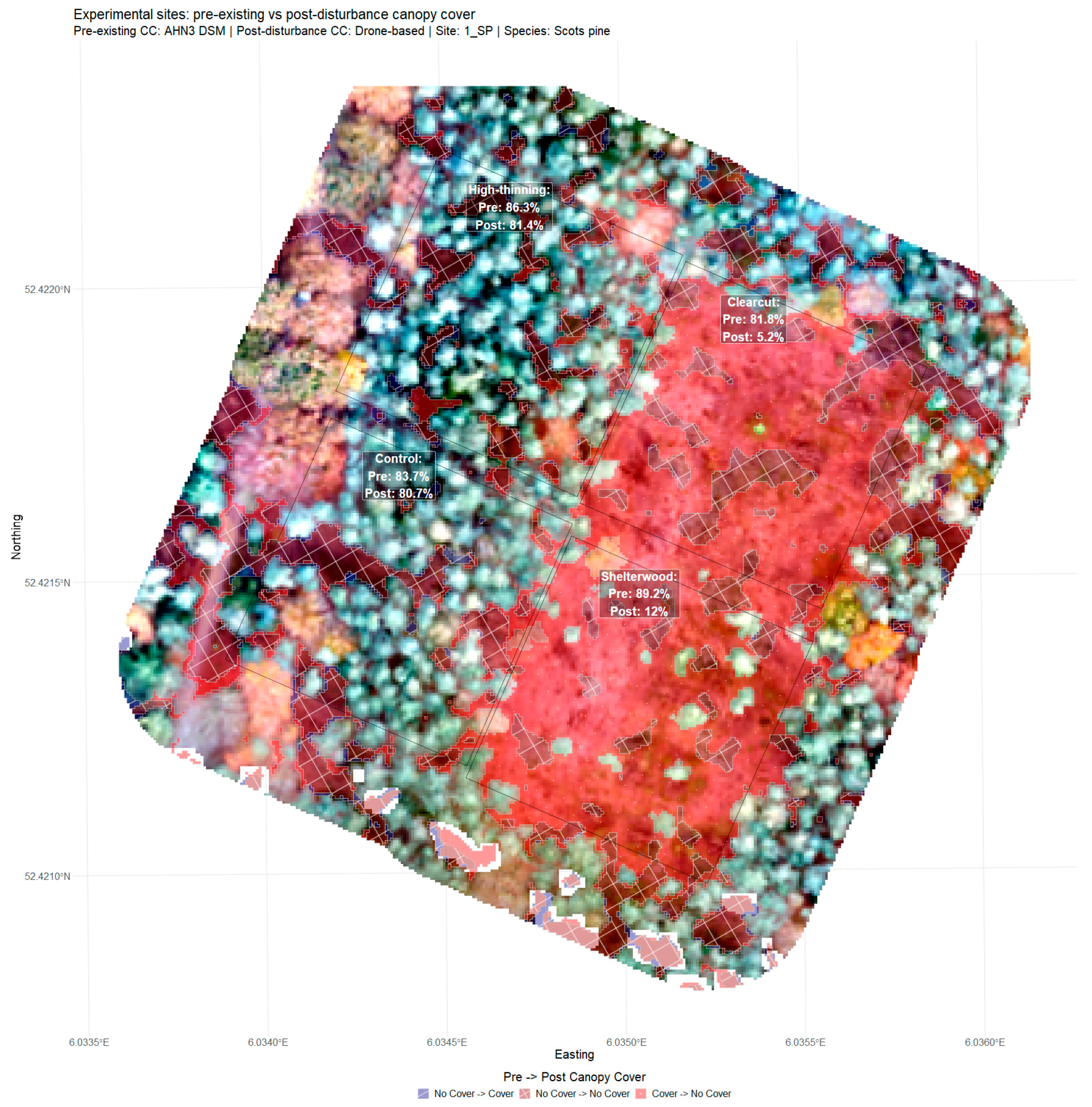
2.1. Study Area and Experimental Sites
2.2. Data
2.2.1. Digital Surface Model
2.2.2. Sentinel-1 Radar
3. Methods
3.1. Preparation of Reference Data
3.2. Assessing Species-Specific Backscatter Seasonality
3.3. Assessing Backscatter Sensitivity to Canopy Cover
3.4. Assessing Backscatter Sensitivity to Canopy Cover Loss and the Effects of Shadow and Layover
3.4.1. Backscatter Sensitivity to Canopy Cover Loss
3.4.2. Effects of Shadow and Layover
3.5. Monitoring Scenarios
4. Results
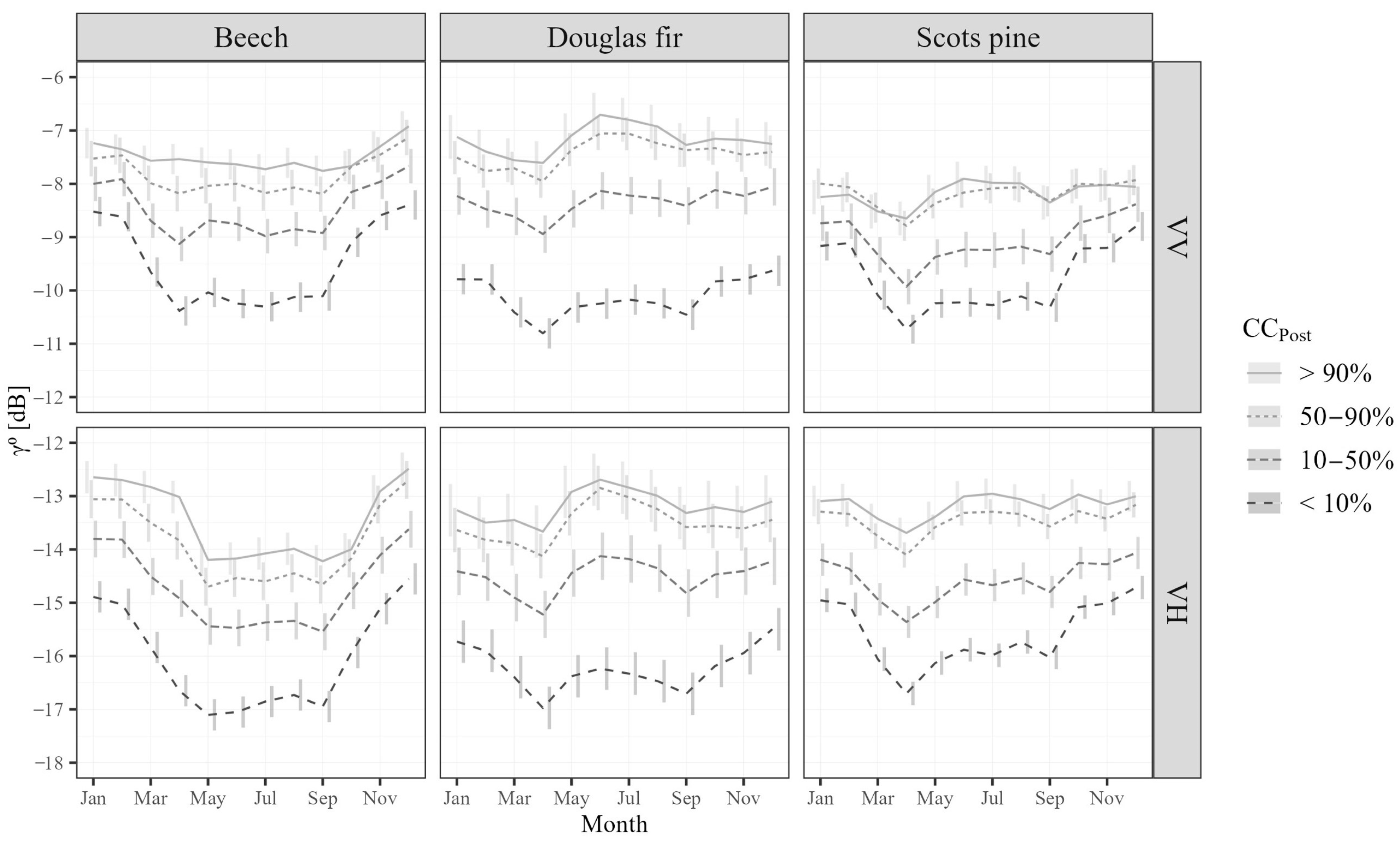
4.1. Backscatter Sensitivity to Species-Specific Seasonality
4.2. Backscatter Sensitivity to Canopy Cover
4.3. Backscatter Sensitivity to Canopy Cover Loss and the Effects of Shadow and Layover
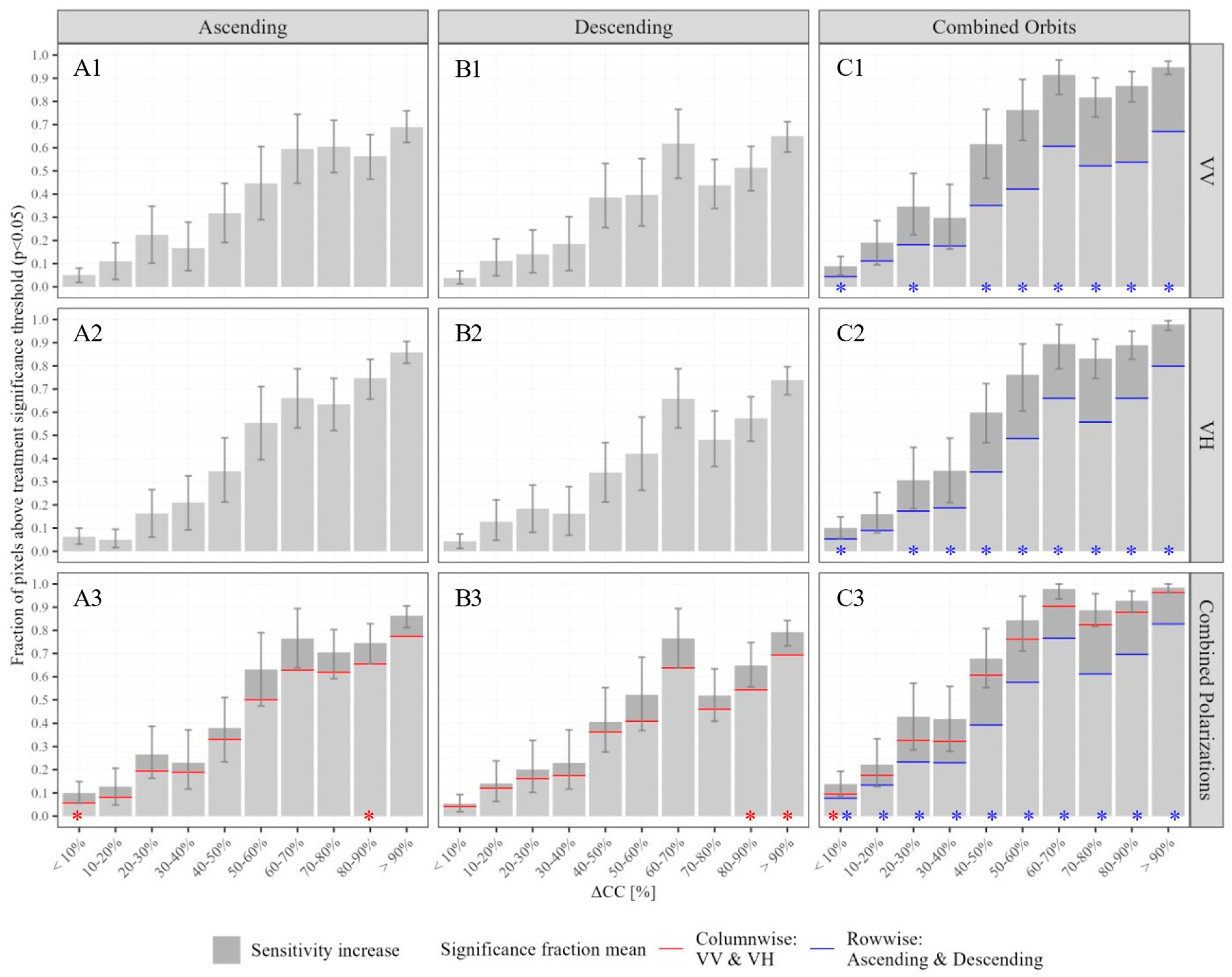
4.4. Monitoring Scenarios
5. Discussion
5.1. Backscatter Seasonality Is Species-Dependent
5.2. Canopy Cover Affects Backscatter Mean and Seasonality
5.3. Backscatter Sensitivity to Canopy Cover Loss Is Affected by Shadow and Layover
5.4. Combining Orbit Directions and Polarizations Increases Detection Sensitivity across Disturbances Severities
5.5. Limitations
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Radar Data Pre-Processing
Appendix A.2. Canopy Cover Layer Generation
Appendix A.3. Shadow and Layover Layers Generation
Appendix B














Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contrast–VH Polarization | ||||
|---|---|---|---|---|
| Beech—Douglas | Beech—Scots Pine | Douglas—Scots Pine | ||
| Month | 1 | <0.001 | <0.001 | <0.001 |
| 2 | <0.001 | <0.001 | 0.010 | |
| 3 | <0.001 | <0.001 | 0.026 | |
| 4 | <0.001 | <0.001 | 0.286 | |
| 5 | 0.130 | 0.293 | 0.886 | |
| 6 | <0.001 | 0.037 | 0.269 | |
| 7 | 0.017 | 0.324 | 0.345 | |
| 8 | 0.152 | 0.080 | 0.951 | |
| 9 | 0.833 | 0.783 | 0.995 | |
| 10 | 0.999 | 0.288 | 0.267 | |
| 11 | <0.001 | 0.003 | 0.054 | |
| 12 | <0.001 | <0.001 | 0.002 | |
| Contrast–VV Polarization | ||||
|---|---|---|---|---|
| Beech—Douglas | Beech—Scots Pine | Douglas—Scots Pine | ||
| Month | 1 | <0.001 | <0.001 | 0.006 |
| 2 | <0.001 | <0.001 | <0.001 | |
| 3 | 0.002 | <0.001 | <0.001 | |
| 4 | 0.095 | <0.001 | <0.001 | |
| 5 | 0.945 | <0.001 | <0.001 | |
| 6 | 0.831 | <0.001 | <0.001 | |
| 7 | 0.193 | <0.001 | <0.001 | |
| 8 | 0.852 | <0.001 | <0.001 | |
| 9 | 0.995 | <0.001 | <0.001 | |
| 10 | 0.936 | <0.001 | <0.001 | |
| 11 | 0.011 | <0.001 | 0.003 | |
| 12 | 0.003 | <0.001 | 0.001 | |
Appendix D
| VV Polarization | VH Polarization | ||||
|---|---|---|---|---|---|
| Partial eta Squared | Confidence Interval | Partial eta Squared | Confidence Interval | ||
| Fixed Effects | species | 0.81 | (0.49, 0.91) | 0.43 | (0.00, 0.70) |
| year | 0.00 | (0.00, 0.00) | 0.01 | (0.00, 0.01) | |
| month | 0.10 | (0.09, 0.11) | 0.10 | (0.09, 0.10) | |
| species:year | 0.00 | (0.00, 0.01) | 0.00 | (0.00, 0,00) | |
| species:month | 0.05 | (0.05, 0.06) | 0.28 | (0.27, 0.29) | |
| year:month | 0.10 | (0.09, 0.11) | 0.14 | (0.13, 0.15) | |
| species:year:month | 0.02 | (0.01, 0.02) | 0.02 | (0.02, 0.02) | |
References
- Nabuurs, G.-J.; Hatab, A.A.; Bustamante, M.; Clark, H.; Havlík, P.; Ninan, K.N.; Popp, A.; Roe, S.; Aoki, L.; Angers, D.; et al. 2022: Agriculture, Forestry and Other Land Uses (AFOLU). In IPCC, 2022: Climate Change 2022: Mitigation of Climate Change. Contribution of Working Group III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; ISBN 978-1-00-915792-6. [Google Scholar]
- European Commission. New EU Forest Strategy for 2030; European Commission: Brussels, Belgium, 2021. [Google Scholar]
- Canadian Council of Forest Ministers. Renewed Forest Bioeconomy Framework. 2021. Available online: https://www.ccfm.org/releases/renewed-forest-bioeconomy-framework/ (accessed on 15 February 2023).
- Bastos, A.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Pongratz, J.; Fan, L.; Wigneron, J.P.; Weber, U.; Reichstein, M.; Fu, Z.; et al. Direct and Seasonal Legacy Effects of the 2018 Heat Wave and Drought on European Ecosystem Productivity. Sci. Adv. 2020, 6, eaba2724. [Google Scholar] [CrossRef] [PubMed]
- Cohen, W.B.; Yang, Z.; Stehman, S.V.; Schroeder, T.A.; Bell, D.M.; Masek, J.G.; Huang, C.; Meigs, G.W. Forest Disturbance across the Conterminous United States from 1985–2012: The Emerging Dominance of Forest Decline. For. Ecol. Manag. 2016, 360, 242–252. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive Shifts in Forest Dynamics in a Changing World. Science 2020, 368, eaaz9463. [Google Scholar] [CrossRef]
- Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant Increase in Natural Disturbance Impacts on European Forests since 1950. Glob. Chang. Biol. 2023, 29, 1359–1376. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances under Climate Change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Ceccherini, G.; Duveiller, G.; Grassi, G.; Lemoine, G.; Avitabile, V.; Pilli, R.; Cescatti, A. Abrupt Increase in Harvested Forest Area over Europe after 2015. Nature 2020, 583, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Palahí, M.; Valbuena, R.; Senf, C.; Acil, N.; Pugh, T.A.M.; Sadler, J.; Seidl, R.; Potapov, P.; Gardiner, B.; Hetemäki, L.; et al. Concerns about Reported Harvests in European Forests. Nature 2021, 592, E15–E17. [Google Scholar] [CrossRef] [PubMed]
- Sebald, J.; Senf, C.; Seidl, R. Human or Natural? Landscape Context Improves the Attribution of Forest Disturbances Mapped from Landsat in Central Europe. Remote Sens. Environ. 2021, 262, 112502. [Google Scholar] [CrossRef]
- Hirschmugl, M.; Gallaun, H.; Dees, M.; Datta, P.; Deutscher, J.; Koutsias, N.; Schardt, M. Methods for Mapping Forest Disturbance and Degradation from Optical Earth Observation Data: A Review. Curr. For. Rep. 2017, 3, 32–45. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R. Mapping the Forest Disturbance Regimes of Europe. Nat. Sustain. 2021, 4, 63–70. [Google Scholar] [CrossRef]
- Francini, S.; McRoberts, R.E.; D’Amico, G.; Coops, N.C.; Hermosilla, T.; White, J.C.; Wulder, M.A.; Marchetti, M.; Mugnozza, G.S.; Chirici, G. An Open Science and Open Data Approach for the Statistically Robust Estimation of Forest Disturbance Areas. Int. J. Appl. Earth Obs. Geoinf. 2022, 106, 102663. [Google Scholar] [CrossRef]
- Giannetti, F.; Pecchi, M.; Travaglini, D.; Francini, S.; D’Amico, G.; Vangi, E.; Cocozza, C.; Chirici, G. Estimating VAIA Windstorm Damaged Forest Area in Italy Using Time Series Sentinel-2 Imagery and Continuous Change Detection Algorithms. Forests 2021, 12, 680. [Google Scholar] [CrossRef]
- Lastovicka, J.; Svec, P.; Paluba, D.; Kobliuk, N.; Svoboda, J.; Hladky, R.; Stych, P. Sentinel-2 Data in an Evaluation of the Impact of the Disturbances on Forest Vegetation. Remote Sens. 2020, 12, 1914. [Google Scholar] [CrossRef]
- Senf, C.; Pflugmacher, D.; Hostert, P.; Seidl, R. Using Landsat Time Series for Characterizing Forest Disturbance Dynamics in the Coupled Human and Natural Systems of Central Europe. ISPRS J. Photogramm. Remote Sens. 2017, 130, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Thonfeld, F.; Gessner, U.; Holzwarth, S.; Kriese, J.; da Ponte, E.; Huth, J.; Kuenzer, C. A First Assessment of Canopy Cover Loss in Germany’s Forests after the 2018–2020 Drought Years. Remote Sens. 2022, 14, 562. [Google Scholar] [CrossRef]
- Gao, Y.; Skutsch, M.; Paneque-Gálvez, J.; Ghilardi, A. Remote Sensing of Forest Degradation: A Review. Environ. Res. Lett. 2020, 15, 103001. [Google Scholar] [CrossRef]
- Pantze, A.; Santoro, M.; Fransson, J.E.S. Change Detection of Boreal Forest Using Bi-Temporal ALOS PALSAR Backscatter Data. Remote Sens. Environ. 2014, 155, 120–128. [Google Scholar] [CrossRef]
- Reiche, J.; Hamunyela, E.; Verbesselt, J.; Hoekman, D.; Herold, M. Improving Near-Real Time Deforestation Monitoring in Tropical Dry Forests by Combining Dense Sentinel-1 Time Series with Landsat and ALOS-2 PALSAR-2. Remote Sens. Environ. 2018, 204, 147–161. [Google Scholar] [CrossRef]
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New Global Forest/Non-Forest Maps from ALOS PALSAR Data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Watanabe, M.; Koyama, C.N.; Hayashi, M.; Nagatani, I.; Tadono, T.; Shimada, M. Refined Algorithm for Forest Early Warning System with ALOS-2/PALSAR-2 ScanSAR Data in Tropical Forest Regions. Remote Sens. Environ. 2021, 265, 112643. [Google Scholar] [CrossRef]
- Tanase, M.A.; Villard, L.; Pitar, D.; Apostol, B.; Petrila, M.; Chivulescu, S.; Leca, S.; Borlaf-Mena, I.; Pascu, I.S.; Dobre, A.C.; et al. Synthetic Aperture Radar Sensitivity to Forest Changes: A Simulations-Based Study for the Romanian Forests. Sci. Total Environ. 2019, 689, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Hethcoat, M.G.; Carreiras, J.M.B.; Edwards, D.P.; Bryant, R.G.; Quegan, S. Detecting Tropical Selective Logging with C-Band SAR Data May Require a Time Series Approach. Remote Sens. Environ. 2021, 259, 112411. [Google Scholar] [CrossRef]
- Reiche, J.; Mullissa, A.; Slagter, B.; Gou, Y.; Tsendbazar, N.E.; Odongo-Braun, C.; Vollrath, A.; Weisse, M.J.; Stolle, F.; Pickens, A.; et al. Forest Disturbance Alerts for the Congo Basin Using Sentinel-1. Environ. Res. Lett. 2021, 16, 024005. [Google Scholar] [CrossRef]
- Doblas Prieto, J.; Lima, L.; Mermoz, S.; Bouvet, A.; Reiche, J.; Watanabe, M.; Sant Anna, S.; Shimabukuro, Y. Inter-Comparison of Optical and SAR-Based Forest Disturbance Warning Systems in the Amazon Shows the Potential of Combined SAR-Optical Monitoring. Int. J. Remote Sens. 2023, 44, 59–77. [Google Scholar] [CrossRef]
- Hoekman, D.; Kooij, B.; Quiñones, M.; Vellekoop, S.; Carolita, I.; Budhiman, S.; Arief, R.; Roswintiarti, O. Wide-Area Near-Real-Time Monitoring of Tropical Forest Degradation and Deforestation Using Sentinel-1. Remote Sens. 2020, 12, 3263. [Google Scholar] [CrossRef]
- Rüetschi, M.; Small, D.; Waser, L.T. Rapid Detection of Windthrows Using Sentinel-1 C-Band SAR Data. Remote Sens. 2019, 11, 115. [Google Scholar] [CrossRef]
- Dascălu, A.; Catalão, J.; Navarro, A. Detecting Deforestation Using Logistic Analysis and Sentinel-1 Multitemporal Backscatter Data. Remote Sens. 2023, 15, 290. [Google Scholar] [CrossRef]
- Olesk, A.; Voormansik, K.; Pohjala, M.; Noorma, M. Forest Change Detection from Sentinel-1 and ALOS-2 Satellite Images. In Proceedings of the 2015 IEEE 5th Asia-Pacific Conference on Synthetic Aperture Radar (APSAR), Singapore, 1–4 September 2015; IEEE: Singapore, 2015; pp. 522–527. [Google Scholar]
- Belenguer-Plomer, M.A.; Tanase, M.A.; Fernandez-Carrillo, A.; Chuvieco, E. Burned Area Detection and Mapping Using Sentinel-1 Backscatter Coefficient and Thermal Anomalies. Remote Sens. Environ. 2019, 233, 111345. [Google Scholar] [CrossRef]
- De Luca, G.; Silva, J.M.N.; Modica, G. A Workflow Based on Sentinel-1 SAR Data and Open-Source Algorithms for Unsupervised Burned Area Detection in Mediterranean Ecosystems. GISci. Remote Sens. 2021, 58, 516–541. [Google Scholar] [CrossRef]
- JRC. Forest Landscape in Europe: Pattern, Fragmentation and Connectivity; Publications Office: Luxembourg, 2013. [Google Scholar]
- Ballère, M.; Bouvet, A.; Mermoz, S.; Le Toan, T.; Koleck, T.; Bedeau, C.; André, M.; Forestier, E.; Frison, P.-L.; Lardeux, C. SAR Data for Tropical Forest Disturbance Alerts in French Guiana: Benefit over Optical Imagery. Remote Sens. Environ. 2021, 252, 112159. [Google Scholar] [CrossRef]
- Bouvet, A.; Mermoz, S.; Ballère, M.; Koleck, T.; Le Toan, T. Use of the SAR Shadowing Effect for Deforestation Detection with Sentinel-1 Time Series. Remote Sens. 2018, 10, 1250. [Google Scholar] [CrossRef]
- Carstairs, H.; Mitchard, E.T.A.; McNicol, I.; Aquino, C.; Chezeaux, E.; Ebanega, M.O.; Dikongo, A.M.; Disney, M. Sentinel-1 Shadows Used to Quantify Canopy Loss from Selective Logging in Gabon. Remote Sens. 2022, 14, 4233. [Google Scholar] [CrossRef]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The Development of Pine Plantation Silviculture in the Southern United States. J. For. 2007, 105, 337–347. [Google Scholar] [CrossRef]
- Gilliam, F.S. Forest Ecosystems of Temperate Climatic Regions: From Ancient Use to Climate Change. New Phytol. 2016, 212, 871–887. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Buddenbaum, H.; Nink, S.; Hill, J. Potential of Sentinel-1 Data for Spatially and Temporally High-Resolution Detection of Drought Affected Forest Stands. Forests 2022, 13, 2148. [Google Scholar] [CrossRef]
- Pulliainen, J.; Hari, P.; Hallikainen, M.; Patrikainen, N.; Peramaki, M.; Kolari, P. Monitoring of Soil Moisture and Vegetation Water Content Variations in Boreal Forest from C-Band SAR Data. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, IGARSS ’04, Anchorage, AK, USA, 20–24 September 2004; IEEE: Anchorage, AK, USA, 2004; Volume 2, pp. 1013–1016. [Google Scholar]
- Dostálová, A.; Wagner, W.; Milenković, M.; Hollaus, M. Annual Seasonality in Sentinel-1 Signal for Forest Mapping and Forest Type Classification. Int. J. Remote Sens. 2018, 39, 7738–7760. [Google Scholar] [CrossRef]
- Dostálová, A.; Lang, M.; Ivanovs, J.; Waser, L.T.; Wagner, W. European Wide Forest Classification Based on Sentinel-1 Data. Remote Sens. 2021, 13, 337. [Google Scholar] [CrossRef]
- Hamunyela, E.; Verbesselt, J.; Herold, M. Using Spatial Context to Improve Early Detection of Deforestation from Landsat Time Series. Remote Sens. Environ. 2016, 172, 126–138. [Google Scholar] [CrossRef]
- Ahern, F.J.; Leckie, D.J.; Drieman, J.A. Seasonal Changes in Relative C-Band Backscatter of Northern Forest Cover Types. IEEE Trans. Geosci. Remote Sens. 1993, 31, 668–680. [Google Scholar] [CrossRef]
- Rüetschi, M.; Schaepman, M.; Small, D. Using Multitemporal Sentinel-1 C-Band Backscatter to Monitor Phenology and Classify Deciduous and Coniferous Forests in Northern Switzerland. Remote Sens. 2017, 10, 55. [Google Scholar] [CrossRef]
- Udali, A.; Lingua, E.; Persson, H.J. Assessing Forest Type and Tree Species Classification Using Sentinel-1 C-Band SAR Data in Southern Sweden. Remote Sens. 2021, 13, 3237. [Google Scholar] [CrossRef]
- Borlaf-Mena, I.; García-Duro, J.; Santoro, M.; Villard, L.; Badea, O.; Tanase, M.A. Seasonality and Directionality Effects on Radar Backscatter Are Key to Identify Mountain Forest Types with Sentinel-1 Data. Remote Sens. Environ. 2023, 296, 113728. [Google Scholar] [CrossRef]
- Dubois, C.; Mueller, M.M.; Pathe, C.; Jagdhuber, T.; Cremer, F.; Thiel, C.; Schmullius, C. Characterization of land cover seasonality in sentinel-1 time series data. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2020; V-3-2020, 97–104. [Google Scholar] [CrossRef]
- Woodhouse, I.H. Introduction to Microwave Remote Sensing; Taylor&Francis: Boca Raton, FL, USA, 2006; ISBN 978-0-415-27123-3. [Google Scholar]
- Sun, G.; Ranson, K.J. A Three-Dimensional Radar Backscatter Model of Forest Canopies. IEEE Trans. Geosci. Remote Sens. 1995, 33, 372–382. [Google Scholar] [CrossRef]
- Vos, M.A.E.; de Boer, D.; de Vries, W.; den Ouden, J.; Sterck, F.J. Aboveground Carbon and Nutrient Distributions Are Hardly Associated with Canopy Position for Trees in Temperate Forests on Poor and Acidified Sandy Soils. For. Ecol. Manag. 2023, 529, 120731. [Google Scholar] [CrossRef]
- Vos, M.A.E.; den Ouden, J.; Hoosbeek, M.; Valtera, M.; de Vries, W.; Sterck, F. The Sustainability of Timber and Biomass Harvest in Perspective of Forest Nutrient Uptake and Nutrient Stocks. For. Ecol. Manag. 2023, 530, 120791. [Google Scholar] [CrossRef]
- Iwasaki, A.; Noda, T. A Framework for Quantifying the Relationship between Intensity and Severity of Impact of Disturbance across Types of Events and Species. Sci. Rep. 2018, 8, 795. [Google Scholar] [CrossRef]
- Schelhaas, M.-J.; Clerkx, S.; Lerink, B. Zevende Nederlandse Bosinventarisatie: 2017–2021; Wettelijke Onderzoekstaken Natuur & Milieu: Wageningen, The Netherlands, 2022. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Fletcher, K. (Ed.) Sentinel-1: ESA’s Radar Observatory Mission for GMES Operational Services; ESA SP-1322/1; ESA Communications: Noordwijk, The Netherlands, 2012; ISBN 978-92-9221-418-0. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Soft. 2017, 82, 26. [Google Scholar] [CrossRef]
- Cnaan, A.; Laird, N.M.; Slasor, P. Using the General Linear Mixed Model to Analyse Unbalanced Repeated Measures and Longitudinal Data. Statist. Med. 1997, 16, 2349–2380. [Google Scholar] [CrossRef]
- Millard, K.; Thompson, D.; Parisien, M.-A.; Richardson, M. Soil Moisture Monitoring in a Temperate Peatland Using Multi-Sensor Remote Sensing and Linear Mixed Effects. Remote Sens. 2018, 10, 903. [Google Scholar] [CrossRef]
- Ben-Shachar, M.; Lüdecke, D.; Makowski, D. Effectsize: Estimation of Effect Size Indices and Standardized Parameters. JOSS 2020, 5, 2815. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 February 2023).
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population Marginal Means in the Linear Model: An Alternative to Least Squares Means. Am. Stat. 1980, 34, 216–221. [Google Scholar] [CrossRef]
- Brodersen, K.H.; Gallusser, F.; Koehler, J.; Remy, N.; Scott, S.L. Inferring Causal Impact Using Bayesian Structural Time-Series Models. Ann. Appl. Stat. 2015, 9, 247–274. [Google Scholar] [CrossRef]
- Muñoz-Sabater, J.; Dutra, E.; Agustí-Panareda, A.; Albergel, C.; Arduini, G.; Balsamo, G.; Boussetta, S.; Choulga, M.; Harrigan, S.; Hersbach, H.; et al. ERA5-Land: A State-of-the-Art Global Reanalysis Dataset for Land Applications. Earth Syst. Sci. Data 2021, 13, 4349–4383. [Google Scholar] [CrossRef]
- Benninga, H.-J.; van der Velde, R.; Su, Z. Impacts of Radiometric Uncertainty and Weather-Related Surface Conditions on Soil Moisture Retrievals with Sentinel-1. Remote Sens. 2019, 11, 2025. [Google Scholar] [CrossRef]
- Cohen, J.; Rautiainen, K.; Lemmetyinen, J.; Smolander, T.; Vehviläinen, J.; Pulliainen, J. Sentinel-1 Based Soil Freeze/Thaw Estimation in Boreal Forest Environments. Remote Sens. Environ. 2021, 254, 112267. [Google Scholar] [CrossRef]
- Zhang, D. A Coefficient of Determination for Generalized Linear Models. Am. Stat. 2017, 71, 310–316. [Google Scholar] [CrossRef]
- Zhang, D. Package ‘Rsq’ 2022. Available online: https://cran.r-project.org/web/packages/rsq/rsq.pdf (accessed on 15 February 2023).
- Kassambara, A. Practical Statistics in R II—Comparing Groups: Numerical Variables, 1st ed.; Datanovia: Montpellier, France, 2019. [Google Scholar]
- Dostálová, A.; Milenković, M.; Hollaus, M.; Wagner, W. Influence of Forest Structure on the Sentinel-1 Backscatter Variation—Analysis with Full-Waveform LIDAR Data. In Proceedings of the Living Planet Symposium, Prague, Czech Republic, 9–13 May 2016; Volume 740, p. 202. [Google Scholar]
- Soudani, K.; Delpierre, N.; Berveiller, D.; Hmimina, G.; Vincent, G.; Morfin, A.; Dufrêne, É. Potential of C-Band Synthetic Aperture Radar Sentinel-1 Time-Series for the Monitoring of Phenological Cycles in a Deciduous Forest. Int. J. Appl. Earth Obs. Geoinf. 2021, 104, 102505. [Google Scholar] [CrossRef]
- Lemmetyinen, J.; Ruiz, J.J.; Cohen, J.; Haapamaa, J.; Kontu, A.; Pulliainen, J.; Praks, J. Attenuation of Radar Signal by a Boreal Forest Canopy in Winter. IEEE Geosci. Remote Sensing Lett. 2022, 19, 1–5. [Google Scholar] [CrossRef]
- Philpot, C.; Mutch, R. The Seasonal Trends in Moisture Content, Ether Extractives, and Energy of Ponderosa Pine and Douglas-Fir Needles; Intermountain Forest & Range Experiment Station, Forest Service, US Department of Agriculture: Ogden, UT, USA, 1971.
- Puhm, M.; Deutscher, J.; Hirschmugl, M.; Wimmer, A.; Schmitt, U.; Schardt, M. A Near Real-Time Method for Forest Change Detection Based on a Structural Time Series Model and the Kalman Filter. Remote Sens. 2020, 12, 3135. [Google Scholar] [CrossRef]
- Ruiz-Ramos, J.; Marino, A.; Boardman, C.; Suarez, J. Continuous Forest Monitoring Using Cumulative Sums of Sentinel-1 Timeseries. Remote Sens. 2020, 12, 3061. [Google Scholar] [CrossRef]
- Magagi, R.; Bernier, M.; Ung, C.H. Quantitative Analysis of RADARSAT SAR data over a Sparse Forest Canopy. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1301–1313. [Google Scholar] [CrossRef]
- Pulliainen, J.T.; Mikhela, P.J.; Hallikainen, M.T.; Ikonen, J.-P. Seasonal Dynamics of C-Band Backscatter of Boreal Forests with Applications to Biomass and Soil Moisture Estimation. IEEE Trans. Geosci. Remote Sens. 1996, 34, 758–770. [Google Scholar] [CrossRef]
- Frison, P.-L.; Fruneau, B.; Kmiha, S.; Soudani, K.; Dufrêne, E.; Le Toan, T.; Koleck, T.; Villard, L.; Mougin, E.; Rudant, J.-P. Potential of Sentinel-1 Data for Monitoring Temperate Mixed Forest Phenology. Remote Sens. 2018, 10, 2049. [Google Scholar] [CrossRef]
- Ulaby, F.; Long, D. Microwave Radar and Radiometric Remote Sensing; University of Michigan Press: Ann Arbor, MI, USA, 2014; ISBN 978-0-472-11935-6. [Google Scholar]
- Schmidt, K.; Tous Ramon, N.; Schwerdt, M. Radiometric Accuracy and Stability of Sentinel-1A Determined Using Point Targets. Int. J. Microw. Wirel. Technol. 2018, 10, 538–546. [Google Scholar] [CrossRef]
- Eriksson, L.E.B.; Fransson, J.E.S.; Soja, M.J.; Santoro, M. Backscatter Signatures of Wind-Thrown Forest in Satellite SAR Images. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium, Munich, Germany, 22–27 July 2012; IEEE: Munich, Germany, 2012; pp. 6435–6438. [Google Scholar]
- Aquino, C.; Mitchard, E.T.A.; McNicol, I.M.; Carstairs, H.; Burt, A.; Puma Vilca, B.L.; Obiang Ebanéga, M.; Modinga Dikongo, A.; Dassi, C.; Mayta, S.; et al. Reliably Mapping Low-Intensity Forest Disturbance Using Satellite Radar Data. Front. For. Glob. Chang. 2022, 5, 1018762. [Google Scholar] [CrossRef]
- Google Developers Sentinel-1 Algorithms. Available online: https://developers.google.com/earth-engine/guides/sentinel1 (accessed on 15 February 2023).
- Mullissa, A.; Vollrath, A.; Odongo-Braun, C.; Slagter, B.; Balling, J.; Gou, Y.; Gorelick, N.; Reiche, J. Sentinel-1 SAR Backscatter Analysis Ready Data Preparation in Google Earth Engine. Remote Sens. 2021, 13, 1954. [Google Scholar] [CrossRef]
- Bauer-Marschallinger, B.; Cao, S.; Navacchi, C.; Freeman, V.; Reuß, F.; Geudtner, D.; Rommen, B.; Vega, F.C.; Snoeij, P.; Attema, E.; et al. The Normalised Sentinel-1 Global Backscatter Model, Mapping Earth’s Land Surface with C-Band Microwaves. Sci Data 2021, 8, 277. [Google Scholar] [CrossRef]
- D’Odorico, P.; Schönbeck, L.; Vitali, V.; Meusburger, K.; Schaub, M.; Ginzler, C.; Zweifel, R.; Velasco, V.M.E.; Gisler, J.; Gessler, A.; et al. Drone-based Physiological Index Reveals Long-term Acclimation and Drought Stress Responses in Trees. Plant Cell Environ. 2021, 44, 3552–3570. [Google Scholar] [CrossRef]
- Penner, J.; Long, D. Ground-Based 3D Radar Imaging of Trees Using a 2D Synthetic Aperture. Electronics 2017, 6, 11. [Google Scholar] [CrossRef]
- Dorman, M.; Erell, E.; Vulkan, A.; Kloog, I. Shadow: R Package for Geometric Shadow Calculations in an Urban Environment. R J. 2019, 11, 287. [Google Scholar] [CrossRef]







| Species | Sites | Median Stand Density | Mean Height | Mean Age | Tree Biomass Distribution [%] | Tree Characteristics | ||
|---|---|---|---|---|---|---|---|---|
| Stem | Branches | Foliage | ||||||
| n | n/ha | m | Years | % | % | % | ||
| Beech | 4 | 240 | 22.9 | 73 | 71.2 ± 3.9 | 29.1 ± 3.8 | - | Broadleaved deciduous, 6–10 cm long oblong leaves, 5–15 m crown diameter, round/oblong crown shape. |
| Douglas fir | 5 | 150 | 35.8 | 68 | 89.3 ± 4.3 | 10.5 ± 1.5 | 1.8 ± 0.2 | Coniferous, 1.5–3.5 cm long needles arranged in two rows along each twig, 5–10 m crown diameter, conical crown shape. |
| Scots pine | 5 | 450 | 18.5 | 57 | 78.4 ± 5.2 | 21.0 ± 2.4 | 3.6 ± 0.4 | Coniferous, 3–7 cm long needles in bundles of 2, 2–5 m crown diameter, conical/ovoid crown shape. |
| Parameter | Symbol | Description | Range | |
|---|---|---|---|---|
| Radar | VV Backscatter coefficient | γ0VV | Ground range detected radiometrically terrain-corrected backscatter coefficient in vertical–vertical (VV) and vertical–horizontal polarizations (VH), respectively. | ~−50 to 1 [dB] * |
| VH Backscatter coefficient | γ0VH | |||
| Canopy | Canopy cover pre-treatment | CCPre | Tree canopy cover as a percentage of total Sentinel-1 pixel area before and after date of treatment, respectively. | 0 to 100 [%] |
| Canopy cover post-treatment | CCPost | |||
| Canopy cover loss | ΔCC | Difference in pre- vs post-treatment canopy cover as a percentage of total Sentinel-1 pixel area: | 0 to 100 [%] | |
| Geometric effects | Layover coverage | L | Layover cover as a percentage of total Sentinel-1 pixel area. | 0 to 100 [%] |
| Shadow coverage | S | Shadow cover as a percentage of total Sentinel-1 pixel area. | 0 to 100 [%] | |
| Relative layover fraction | LRel | Layover normalized by post-treatment non-canopy cover: | 0 to 1 | |
| Relative shadow fraction | SRel | Shadow normalized by post-treatment non-canopy cover: | 0 to 1 | |
| Relative layover-shadow fraction | LS | Relative layover fraction minus relative shadow fraction: | −1 to 1 |
| Period | Canopy Parameters | Canopy Cover Classes | Orbit-Polarization Combinations |
| Pre-treatment: 1 January 2016–1 January 2019 | CCPre | >90% | VV-combined orbits VH-combined orbits |
| Post-treatment: 1 April 2019–1 April 2022 | CCPost | <10%, 10–50%, 50–90%, >90% | VV-ascending VV-descending VH-ascending VH-descending |
| Entire: 1 January 2016–1 April 2022 | ΔCC | <10%, 10–20%, 30–40%, 40–50%, 50–60%, 60–70%, 70–80%, 80–90%, >90% | VV-ascending VV-descending VH-ascending VH-descending |
| Model | Species | Polarization | Coefficient Significance | Adjusted Partial R2 | |||
|---|---|---|---|---|---|---|---|
| Srel | Srel × ΔCC | Lrel | Lrel × ΔCC | ||||
| Combined (Srel, Lrel) | Beech | VV | * | - | * | * | 0.33 |
| Douglas fir | * | - | * | * | 0.21 | ||
| Scots pine | * | - | * | * | 0.39 | ||
| Beech | VH | * | * | * | * | 0.24 | |
| Douglas fir | - | - | * | * | 0.15 | ||
| Scots pine | * | * | * | * | 0.37 | ||
| Shadow only (Srel) | Beech | VV | - | * | 0.05 | ||
| Douglas fir | - | * | 0.07 | ||||
| Scots pine | - | * | 0.06 | ||||
| Beech | VH | - | - | 0.00 | |||
| Douglas fir | - | * | 0.05 | ||||
| Scots pine | - | - | 0.01 | ||||
| Layover only (Lrel) | Beech | VV | - | * | 0.27 | ||
| Douglas fir | * | * | 0.17 | ||||
| Scots pine | * | * | 0.34 | ||||
| Beech | VH | - | * | 0.22 | |||
| Douglas fir | - | * | 0.13 | ||||
| Scots pine | * | * | 0.35 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Woude, S.; Reiche, J.; Sterck, F.; Nabuurs, G.-J.; Vos, M.; Herold, M. Sensitivity of Sentinel-1 Backscatter to Management-Related Disturbances in Temperate Forests. Remote Sens. 2024, 16, 1553. https://doi.org/10.3390/rs16091553
van der Woude S, Reiche J, Sterck F, Nabuurs G-J, Vos M, Herold M. Sensitivity of Sentinel-1 Backscatter to Management-Related Disturbances in Temperate Forests. Remote Sensing. 2024; 16(9):1553. https://doi.org/10.3390/rs16091553
Chicago/Turabian Stylevan der Woude, Sietse, Johannes Reiche, Frank Sterck, Gert-Jan Nabuurs, Marleen Vos, and Martin Herold. 2024. "Sensitivity of Sentinel-1 Backscatter to Management-Related Disturbances in Temperate Forests" Remote Sensing 16, no. 9: 1553. https://doi.org/10.3390/rs16091553
APA Stylevan der Woude, S., Reiche, J., Sterck, F., Nabuurs, G. -J., Vos, M., & Herold, M. (2024). Sensitivity of Sentinel-1 Backscatter to Management-Related Disturbances in Temperate Forests. Remote Sensing, 16(9), 1553. https://doi.org/10.3390/rs16091553