
Spatiotemporal Variability in Start and End of Growing Season in China Related to Climate Variability



Abstract
:
1. Introduction
2. Materials and Methods
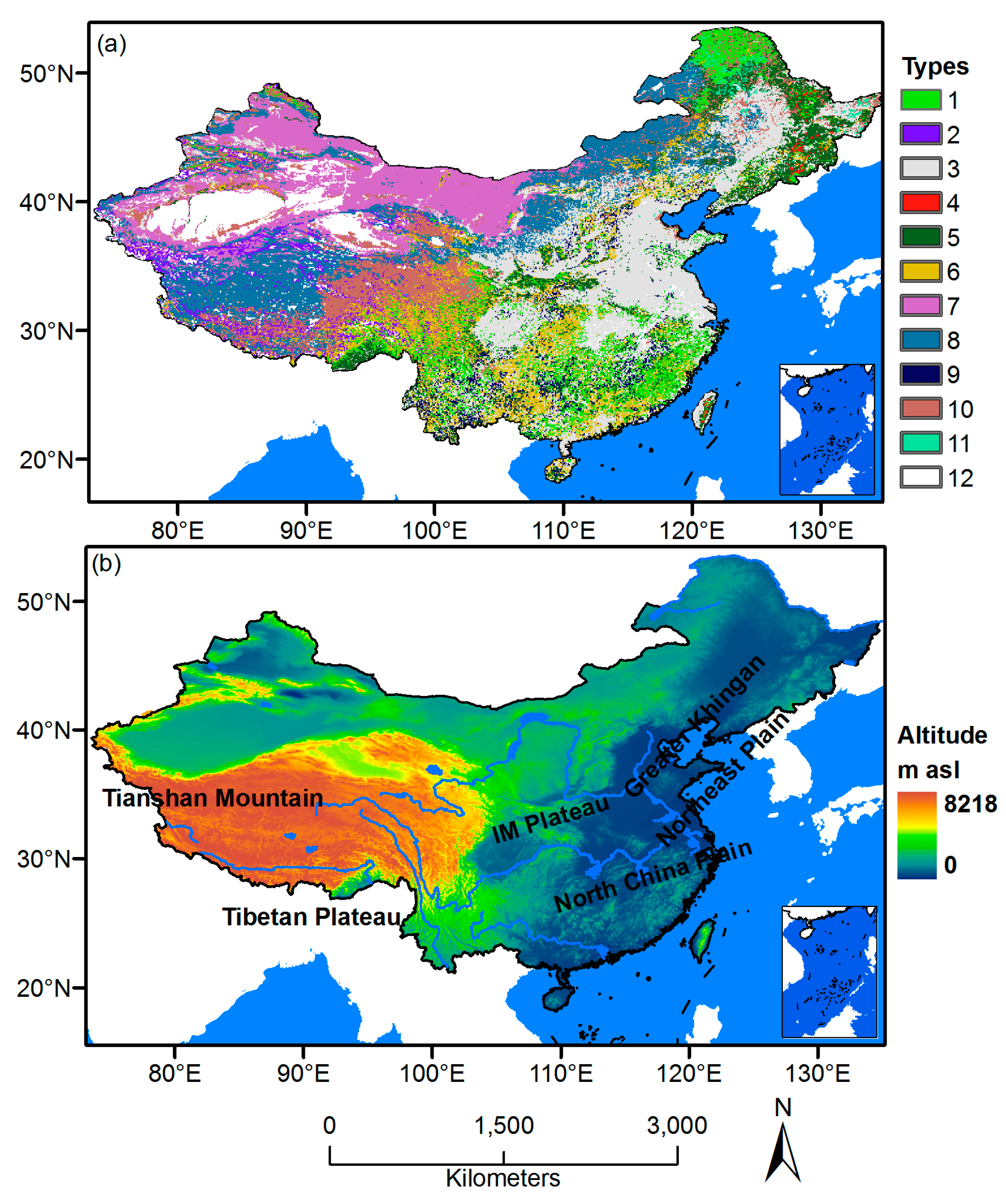
2.1. Data Sources
2.2. Methods
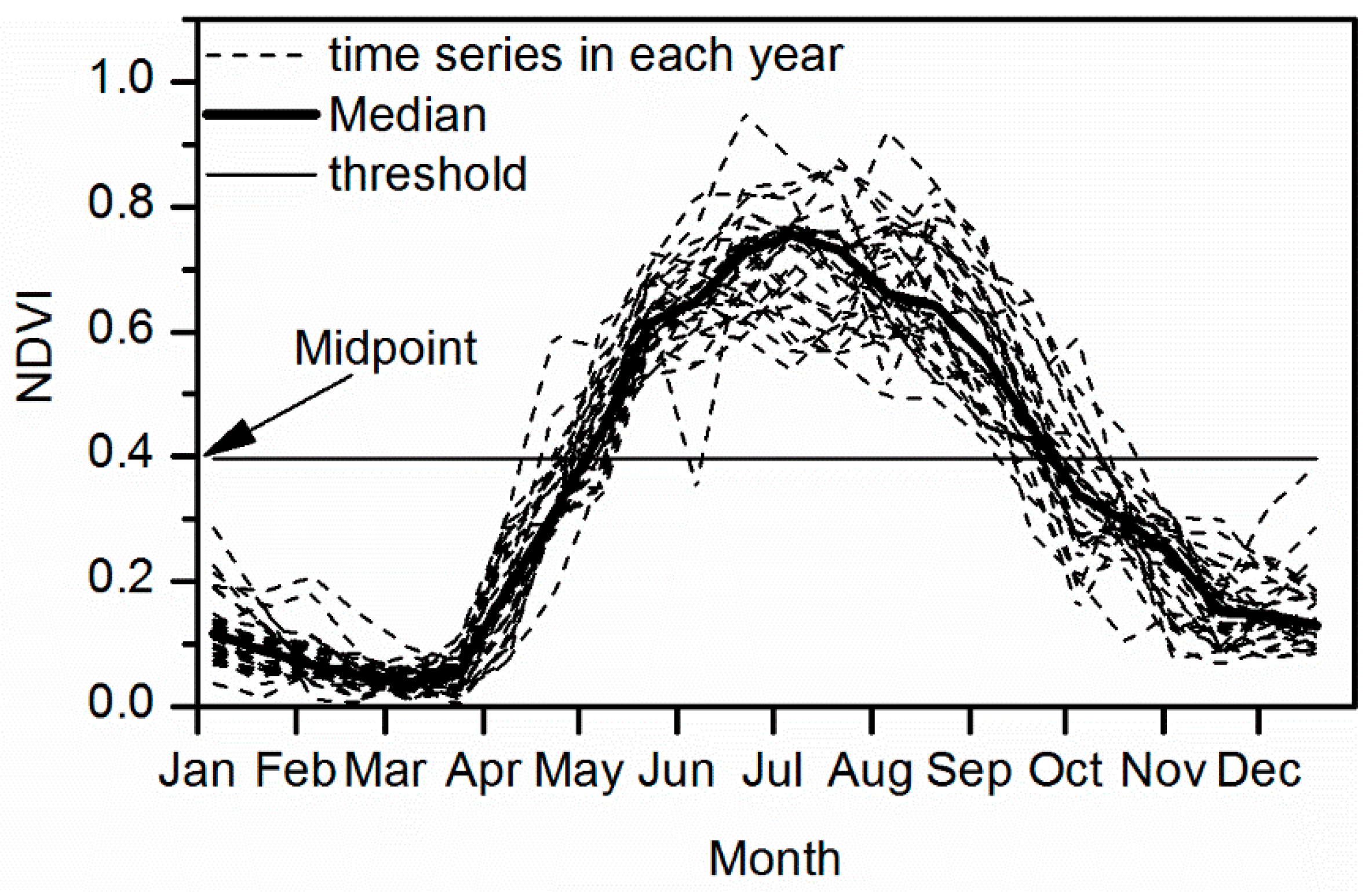
2.2.1. Estimation of the Start and End of Growing Season
- (1)
- Annual maximum NDVI > 0.15 and mean annual NDVI > 0.10 [36]. Such thresholds excluded deserts and sparsely vegetated areas, where the soil background would noticeably impact the spectral signals of vegetation.
- (2)
- The vegetation type should be natural vegetation, since the phenology of cultivated vegetation is strongly impacted by human activities [37].
- (3)
- The annual maximum NDVI occurred between June and September; mean NDVI of July to August >1.35 × NDVI of November to December; mean NDVI of July to August >1.35 × NDVI of January to February [36]. These criteria excluded the vegetation lack of seasonality, e.g., the evergreen vegetation over the humid tropics and subtropics, where aberrant NDVI fluctuation related to weather impact often occurred [37].
2.2.2. Empirical Orthogonal Function (EOF) Analysis
2.2.3. Canonical Correlation Analysis (CCA)
3. Results
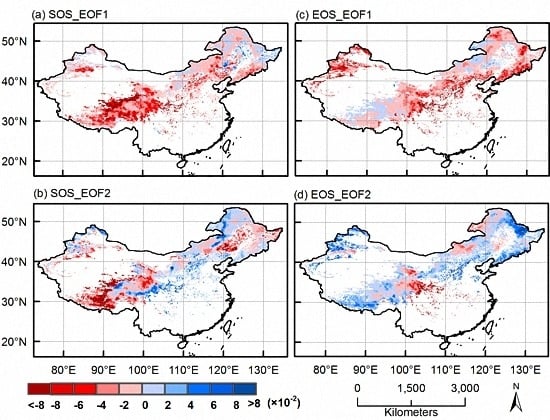
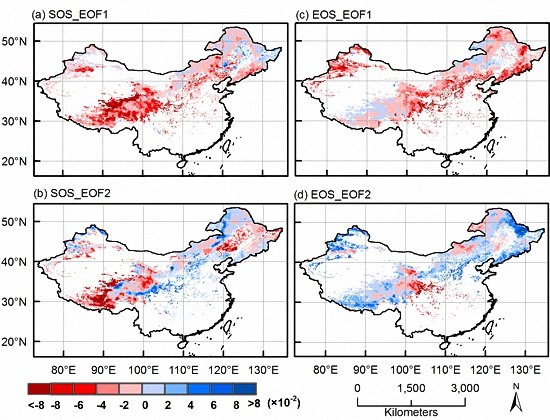
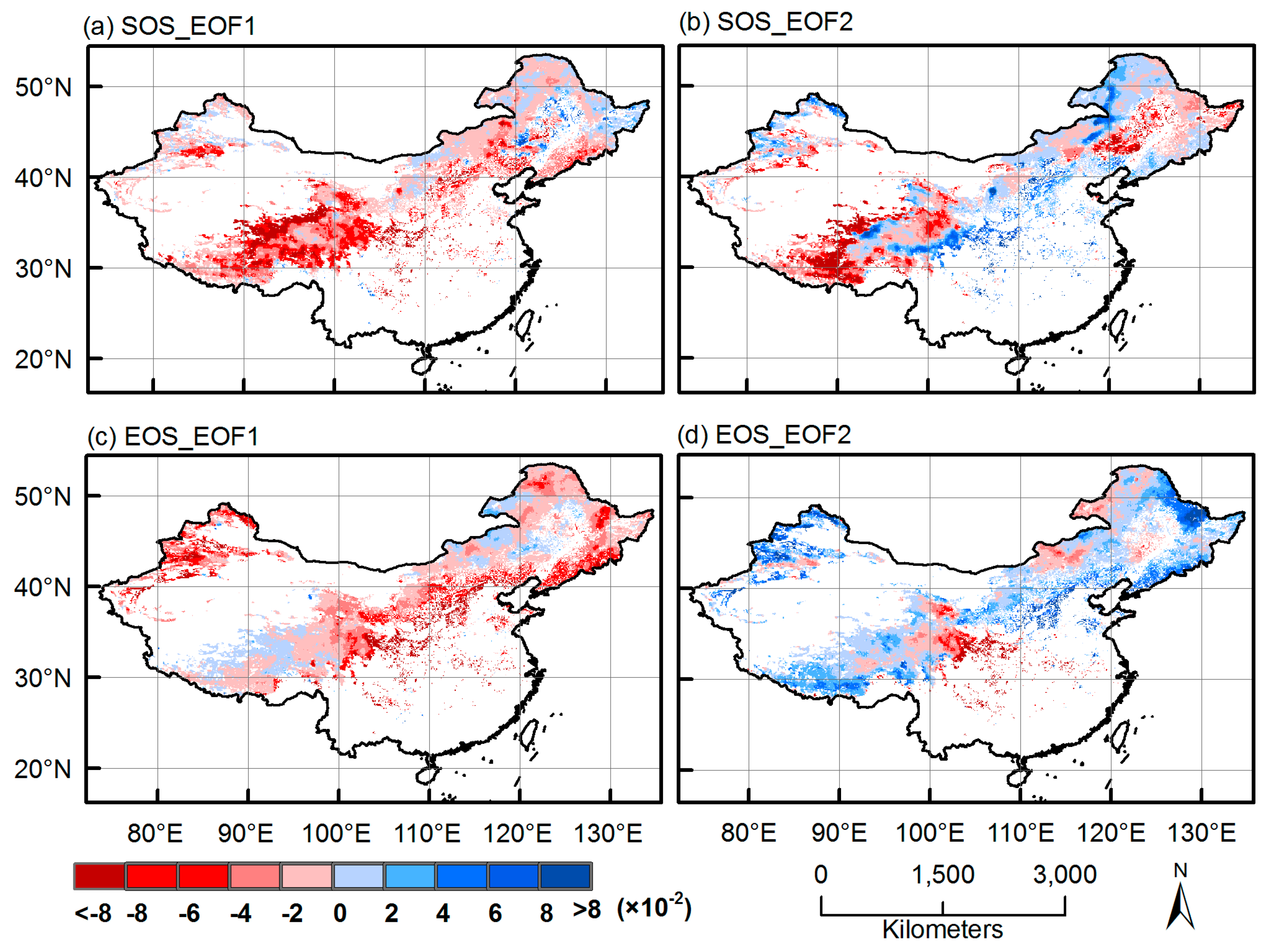
3.1. EOF Analyses
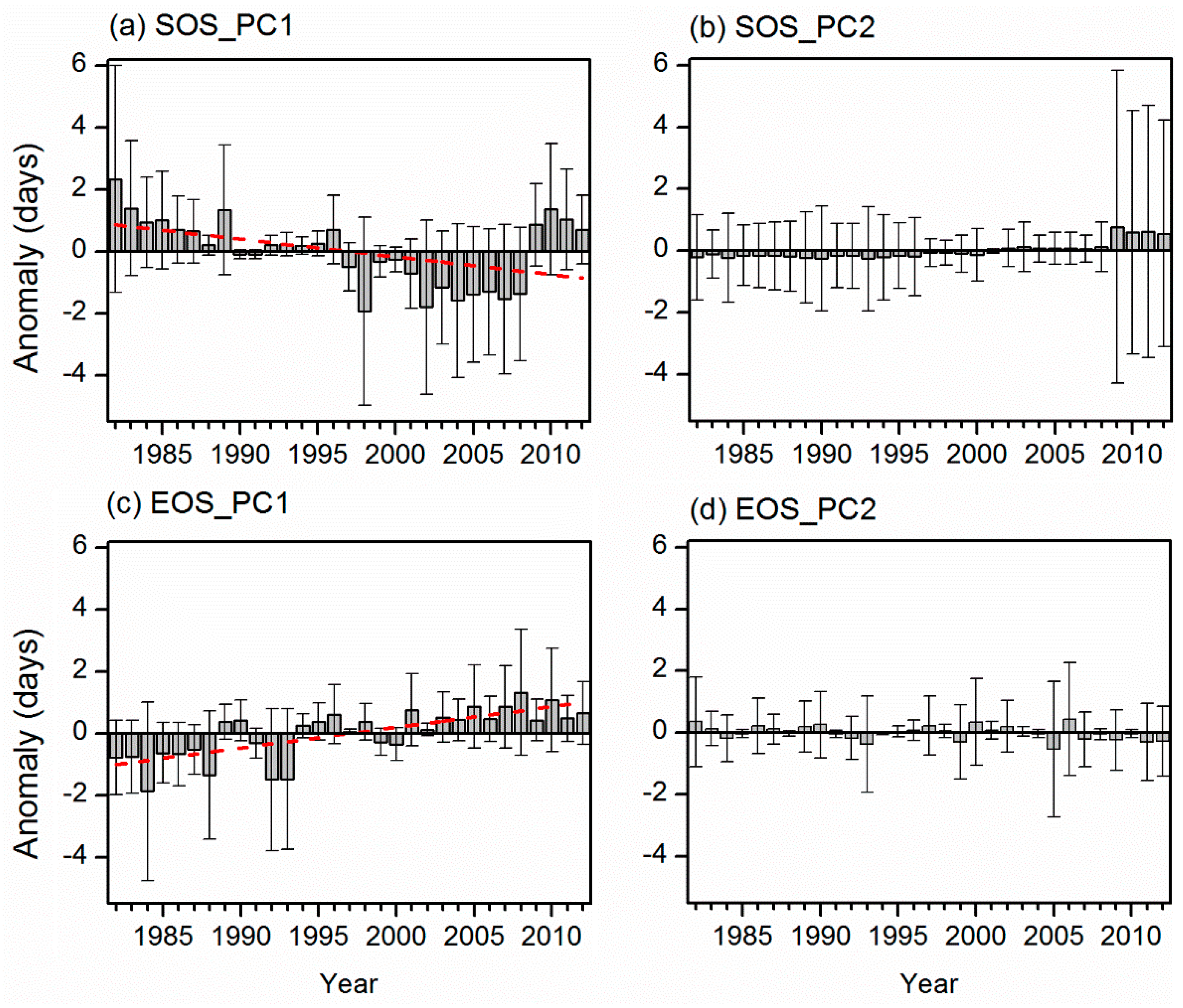
3.2. Dominant Phenological Patterns and Their Time Evolutions
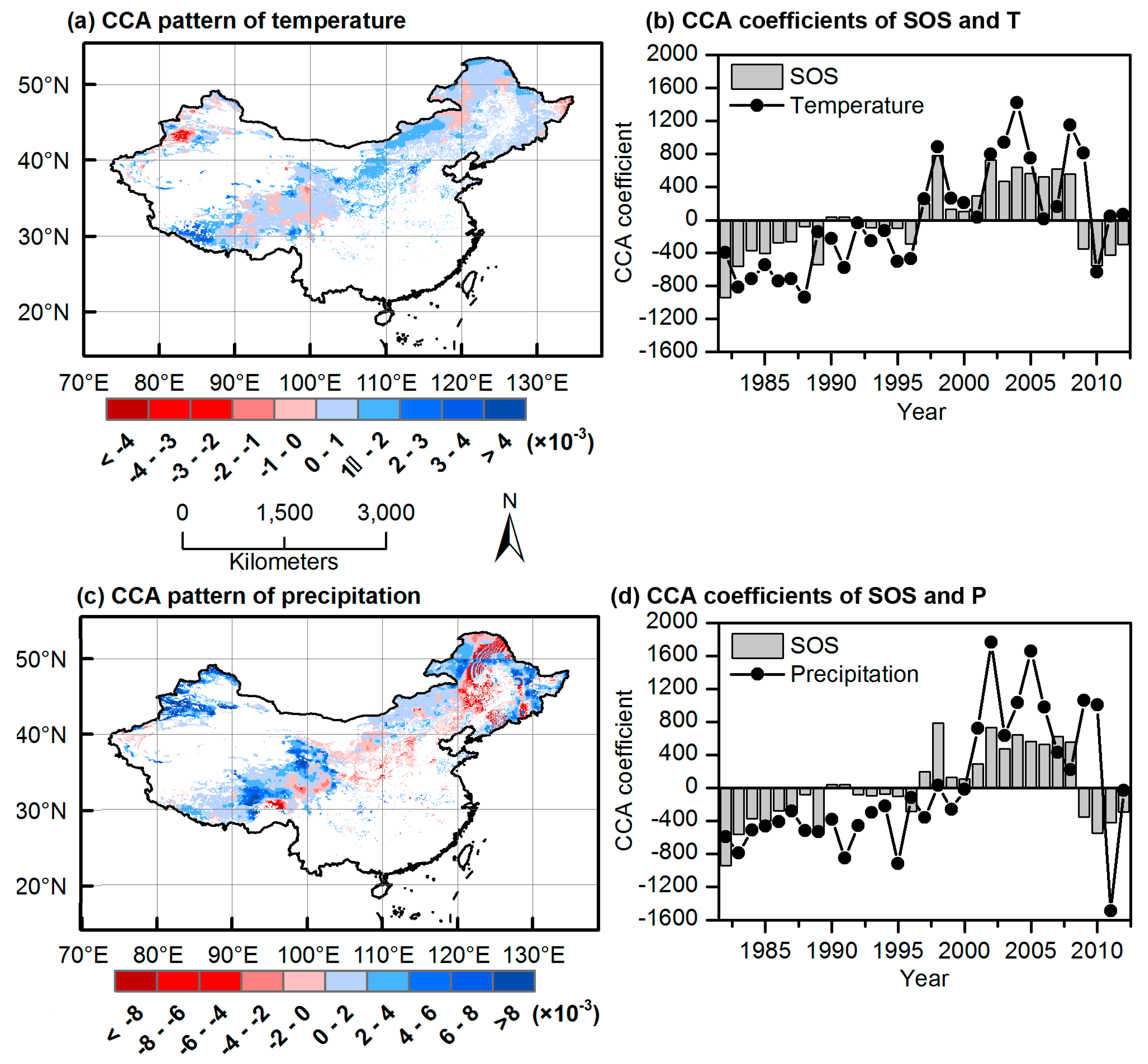
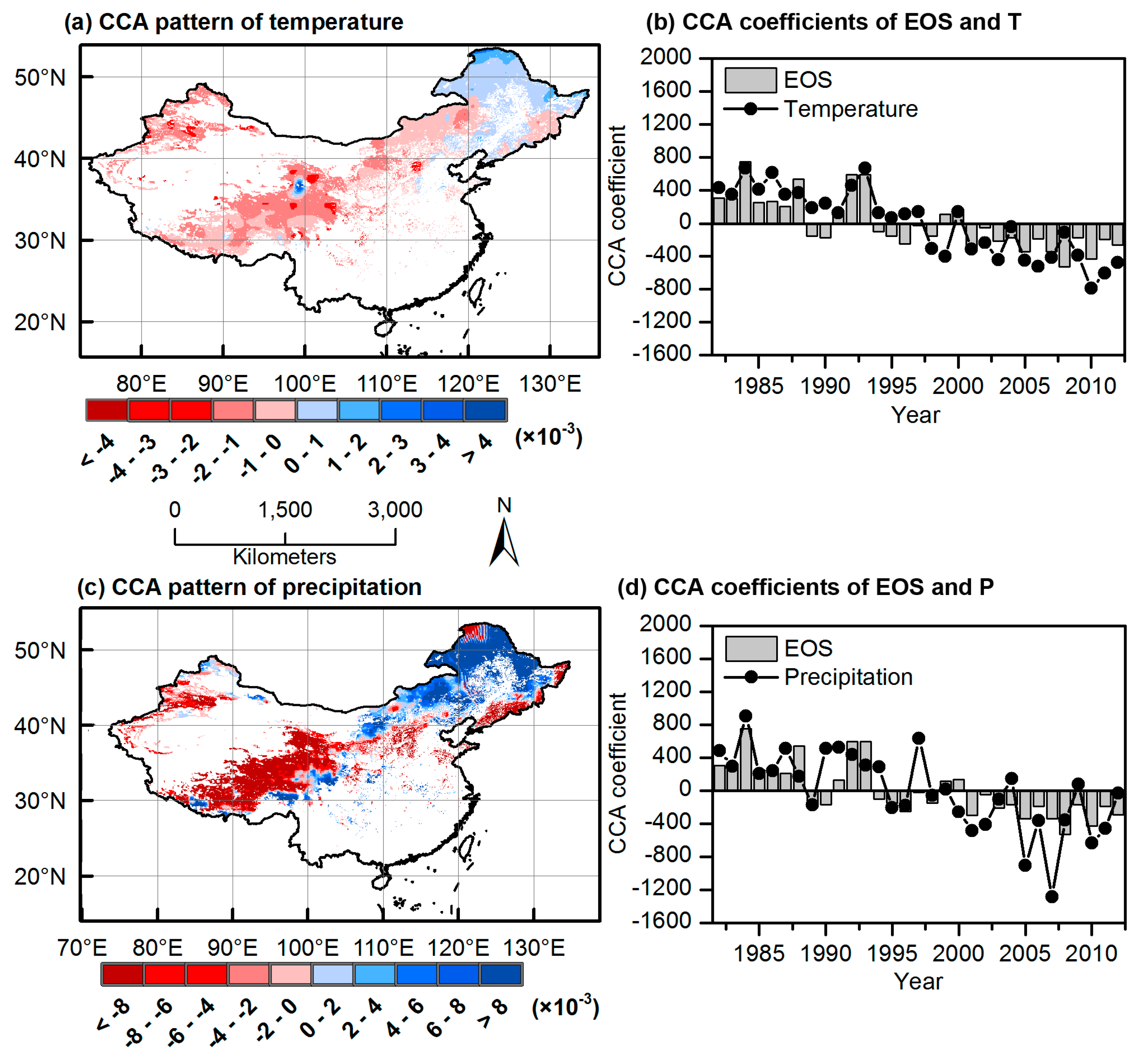
3.3. CCA Patterns and Time Coefficients of Climate and Phenology
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sultan, S.E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Schwartz, M.D. Phenology: An Integrative Environmental Science, 2nd ed.; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 1–2. [Google Scholar]
- Ge, Q.; Wang, H.; Rutishauser, T.; Dai, J. Phenological response to climate change in China: A meta-analysis. Glob. Change Biol. 2015, 21, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Change Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Chambers, L.E.; Altwegg, R.; Barbraud, C.; Barnard, P.; Beaumont, L.J.; Crawford, R.J.M.; Durant, J.M.; Hughes, L.; Keatley, M.R.; Low, M.; et al. Phenological Changes in the Southern Hemisphere. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, A.L.; Gallinat, A.S.; Sanders-DeMott, R.; Rigden, A.J.; Gianotti, D.J.S.; Mantooth, J.A.; Templer, P.H. Changes in autumn senescence in northern hemisphere deciduous trees: A meta-analysis of autumn phenology studies. Ann. Bot. Lond. 2015, 116, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Rutishauser, T.; Filella, I. Phenology feedbacks on climate change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef] [PubMed]
- Morisette, J.T.; Richardson, A.D.; Knapp, A.K.; Fisher, J.I.; Graham, E.A.; Abatzoglou, J.; Wilson, B.E.; Breshears, D.D.; Henebry, G.M.; Hanes, J.M.; et al. Tracking the rhythm of the seasons in the face of global change: Phenological research in the 21st century. Front. Ecol. Environ. 2008, 7, 253–260. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Hanes, J.M.; Liang, L. Comparing carbon flux and high-resolution spring phenological measurements in a northern mixed forest. Agric. Forest Meteorol. 2013, 169, 136–147. [Google Scholar] [CrossRef]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Change 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Richardson, A.D.; Andy Black, T.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. Royal Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Studer, S.; Stöckli, R.; Appenzeller, C.; Vidale, P.L. A comparative study of satellite and ground-based phenology. Int. J. Biometeorol. 2007, 51, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Hanes, J.M. Intercomparing multiple measures of the onset of spring in eastern North America. Int. J. Climatol. 2010, 30, 1614–1626. [Google Scholar] [CrossRef]
- Reed, B.C.; Schwartz, M.D.; Xiao, X. Remote sensing phenology. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer New York: New York, NY, USA, 2009; pp. 231–246. [Google Scholar]
- Hanes, J.; Liang, L.; Morisette, J. Land Surface Phenology. In Biophysical Applications of Satellite Remote Sensing; Hanes, J.M., Ed.; Springer: Berlin, Germany, 2014; pp. 99–125. [Google Scholar]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Global Change Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guan, H.; Shen, M.; Liang, W.; Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Global Change Biol. 2015, 21, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Garonna, I.; de Jong, R.; de Wit, A.J.W.; Mücher, C.A.; Schmid, B.; Schaepman, M.E. Strong contribution of autumn phenology to changes in satellite-derived growing season length estimates across Europe (1982–2011). Glob. Change Biol. 2014, 20, 3457–3470. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Tian, H.; Xu, X.; Pan, Y.; Chen, G.; Lin, W. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- Hoscilo, A.; Balzter, H.; Bartholome, E.; Boschetti, M.; Brivio, P.A.; Brink, A.; Clerici, M.; Pekel, J.F. A conceptual model for assessing rainfall and vegetation trends in sub-Saharan Africa from satellite data. Int. J. Climatol. 2015, 35, 3582–3592. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Xu, X.; Ciais, P.; MacBean, N.; Myneni, R.B.; Li, L. Has the advancing onset of spring vegetation green-up slowed down or changed abruptly over the last three decades? Glob. Ecol. Biogeogr. 2015, 24, 621–631. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, B.; Yu, Y. Interannual variations and trends in global land surface phenology derived from enhanced vegetation index during 1982–2010. Int. J. Biometeorol. 2014, 58, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Ho, C.; Gim, H.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Chen, J.; Yang, X.; Wang, C.; Cui, X.; Yang, Y.; Han, L.; Li, L.; Du, J.; et al. Earlier-Season Vegetation Has Greater Temperature Sensitivity of Spring Phenology in Northern Hemisphere. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Cayan, D.R.; Kammerdiener, S.A.; Dettinger, M.D.; Caprio, J.M.; Peterson, D.H. Changes in the onset of spring in the western United States. Bull. Am. Meteorol. Soc. 2001, 82, 399–416. [Google Scholar] [CrossRef]
- Studer, S.; Appenzeller, C.; Defila, C. Inter-annual variability and decadal trends in alpine spring phenology: A multivariate analysis approach. Clim. Change 2005, 73, 395–414. [Google Scholar] [CrossRef]
- Zhao, S. Physical Geography of China; Science Press: Beijing, China, 1986; pp. 1–209. [Google Scholar]
- Asrar, G.; Fuchs, M.; Kanemasu, E.T.; Hatfield, J.L. Estimating absorbed photosynthetic radiation and leaf area index from spectral reflectance in wheat. Agron. J. 1984, 76, 300–306. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Reed, B.C.; White, M.A. Assessing satellite-derived start-of-season measures in the conterminous USA. Int. J. Climatol. 2002, 22, 1793–1805. [Google Scholar] [CrossRef]
- Pinzon, J.E.; Tucker, C.J. A Non-Stationary 1981–2012 AVHRR NDVI3g Time Series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Editorial Board of the Vegetation Map of China. Vegetation Map of the People's Republic of China (1: 10,000,000); Geological Publishing House: Beijing, China, 2007. [Google Scholar]
- He, J.; Yang, K. China Meteorological Forcing Dataset. Cold Arid Reg. Sci. Data Cent. Lanzhou 2011. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Chen, J.; Zhu, X.; Zheng, Y. Influences of temperature and precipitation before the growing season on spring phenology in grasslands of the central and eastern Qinghai-Tibetan Plateau. Agric. Forest Meteorol. 2011, 151, 1711–1722. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; Zhou, L.M.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Change Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- White, M.A.; De Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Change Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Wang, H.; Dai, J.; Ge, Q. Comparison of satellite and ground-based phenology in China’s temperate monsoon area. Adv. Meteorol. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Schaber, J.; Badeck, F.W. Evaluation of methods for the combination of phenological time series and outlier detection. Tree Physiol. 2002, 22, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ottlé, C.; Peng, S.; Janssens, I.A.; Lin, X.; Poulter, B.; Yue, C.; Ciais, P. The influence of local spring temperature variance on temperature sensitivity of spring phenology. Glob. Change Biol. 2014, 20, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Clim. Change 2003, 57, 243–263. [Google Scholar] [CrossRef]
- Bolmgren, K.; Vanhoenacker, D.; Miller-Rushing, A.J. One man, 73 years, and 25 species. Evaluating phenological responses using a lifelong study of first flowering dates. Int. J. Biometeorol. 2013, 57, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Bretherton, C.S.; Smith, C.; Wallace, J.M. An intercomparison of methods for finding coupled patterns in climate data. J. Climate 1992, 5, 541–560. [Google Scholar] [CrossRef]
- Hotelling, H. Relations between two sets of variates. Biometrika 1936, 28, 321–377. [Google Scholar] [CrossRef]
- Von Storch, H. Spatial patterns: EOFs and CCA. In Analysis of Climate Variability; von Storch, H., Navarra, A., Eds.; Springer: Berlin, Germany; Heidelberg, Germany, 1999; pp. 231–263. [Google Scholar]
- Franklin, D.C.; Bach, C.S. Assessing intraspecific phenological synchrony in zoochorous trees from the monsoon forests of northern Australia. J. Trop. Ecol. 2006, 22, 419–429. [Google Scholar] [CrossRef]
- Sparks, T.H.; Mizera, T.; Wojtowicz, W.; Tryjanowski, P. Synchrony in the phenology of a culturally iconic spring flower. Int. J. Biometeorol. 2012, 56, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, T.; Luterbacher, J.; Jeanneret, F.; Pfister, C.; Wanner, H. A phenology-based reconstruction of interannual changes in past spring seasons. J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhang, Y.; Dong, J.; Xiao, X. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Nat. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, H. Consistent shifts in spring vegetation green-up date across temperate biomes in China, 1982–2006. Glob. Change Biol. 2013, 19, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biol. 2007, 13, 1860–1872. [Google Scholar] [CrossRef]
- Chmielewski, F.; Rötzer, T. Annual and spatial variability of the beginning of growing season in Europe in relation to air temperature changes. Clim. Res. 2002, 19, 257–264. [Google Scholar] [CrossRef]
- Liu, G.; Tang, Q.; Liu, X.; Dai, J.; Zhang, X.; Ge, Q.; Tang, Y. Spatiotemporal analysis of ground-based woody plant leafing in response to temperature in temperate eastern China. Int. J. Biometeorol. 2014, 58, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Badeck, F.W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Ge, Q.; Hao, Z.; Wang, W.C. Spring phenophases in recent decades over eastern China and its possible link to climate changes. Clim. Change 2006, 77, 449–462. [Google Scholar] [CrossRef]
- Schmidt, H.; Karnieli, A. Remote sensing of the seasonal variability of vegetation in a semi-arid environment. J. Arid Environ. 2000, 45, 43–59. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in Africa by coupling MODIS and TRMM instruments. J. Geophys. Res. Atmos. 2005, 110. [Google Scholar] [CrossRef]
- Dalmolin, A.N.C.; de Almeida, L.F.; Vourlitis, G.; Silva, P.R.; Dalmagro, H.J.E.; Antunes, M.Z.E.A., Jr.; Ortiz, C.E.R. Is the dry season an important driver of phenology and growth for two Brazilian savanna tree species with contrasting leaf habits? Plant Ecol. 2015, 216, 407–417. [Google Scholar] [CrossRef]
- Yu, H.; Luedeling, E.; Xu, J. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proc. Nat. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Global Change Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; An, S.; Inouye, D.; Schwartz, M. Temperature and snowfall trigger alpine vegetation green-up on the world’s roof. Global Change Biol. 2015, 21, 3635–3646. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, D.; Lin, H.; Montenegro, A.; Zhu, X. NDVI and vegetation phenology dynamics under the influence of sunshine duration on the Tibetan plateau. Int. J. Climatol. 2015, 35, 687–698. [Google Scholar] [CrossRef]
- Dai, J.; Wang, H.; Ge, Q. The spatial pattern of leaf phenology and its response to climate change in China. Int. J. Biometeorol. 2014, 58, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; François, C.; Delpierre, N.; Dufrêne, E.; Kremer, A.; Chuine, I.; Delzon, S. Assessing the effects of climate change on the phenology of European temperate trees. Agric. Forest Meteorol. 2011, 151, 969–980. [Google Scholar] [CrossRef]
- Estrella, N.; Menzel, A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany. Clim. Res. 2006, 32, 253–267. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Vegetation Types | Number of Pixels | Number of Pixels Analyzed | Percentage |
|---|---|---|---|---|
| 1 | Needleleaf forest | 11,349 | 3882 | 34.2% |
| 2 | Alpine vegetation | 4581 | 1892 | 41.3% |
| 3 | Cultivated vegetation | 30,158 | 0 | 0.0% |
| 4 | Mixed forest | 362 | 299 | 82.6% |
| 5 | Broadleaf forests | 10,309 | 6405 | 62.1% |
| 6 | Scrub | 12,361 | 4664 | 37.7% |
| 7 | Desert vegetation | 18,379 | 2433 | 13.2% |
| 8 | Steppe | 20,806 | 13,125 | 63.1% |
| 9 | Grass-forb community | 3936 | 737 | 18.7% |
| 10 | Meadow | 14,888 | 12,351 | 83.0% |
| 11 | Swamp | 1131 | 1069 | 94.5% |
| 12 | Nonvegetated area | 9466 | 0 | 0.0% |
| Sum | 137,726 | 46,857 | 34.0% |
| EOF Mode | Phenology | Temperature | Precipitation |
|---|---|---|---|
| Start of growing season | |||
| 1 | 0.17 | 0.46 | 0.20 |
| 2 | 0.14 | 0.20 | 0.13 |
| 3 | 0.10 | 0.11 | 0.09 |
| 4 | 0.07 | 0.04 | 0.07 |
| 5 | 0.05 | 0.03 | 0.05 |
| Sum | 0.53 | 0.84 | 0.54 |
| End of growing season | |||
| 1 | 0.19 | 0.50 | 0.18 |
| 2 | 0.08 | 0.12 | 0.14 |
| 3 | 0.06 | 0.08 | 0.10 |
| 4 | 0.06 | 0.05 | 0.08 |
| 5 | 0.05 | 0.05 | 0.06 |
| Sum | 0.44 | 0.80 | 0.56 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, Q.; Dai, J.; Cui, H.; Wang, H. Spatiotemporal Variability in Start and End of Growing Season in China Related to Climate Variability. Remote Sens. 2016, 8, 433. https://doi.org/10.3390/rs8050433
Ge Q, Dai J, Cui H, Wang H. Spatiotemporal Variability in Start and End of Growing Season in China Related to Climate Variability. Remote Sensing. 2016; 8(5):433. https://doi.org/10.3390/rs8050433
Chicago/Turabian StyleGe, Quansheng, Junhu Dai, Huijuan Cui, and Huanjiong Wang. 2016. "Spatiotemporal Variability in Start and End of Growing Season in China Related to Climate Variability" Remote Sensing 8, no. 5: 433. https://doi.org/10.3390/rs8050433
APA StyleGe, Q., Dai, J., Cui, H., & Wang, H. (2016). Spatiotemporal Variability in Start and End of Growing Season in China Related to Climate Variability. Remote Sensing, 8(5), 433. https://doi.org/10.3390/rs8050433