Analyzing the Long-Term Phenological Trends of Salt Marsh Ecosystem across Coastal LOUISIANA



Abstract
:
1. Introduction
2. Materials and Methods
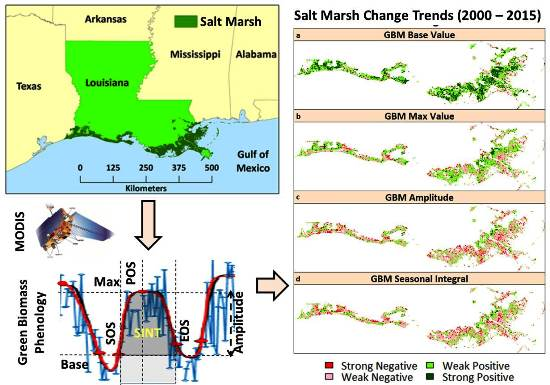
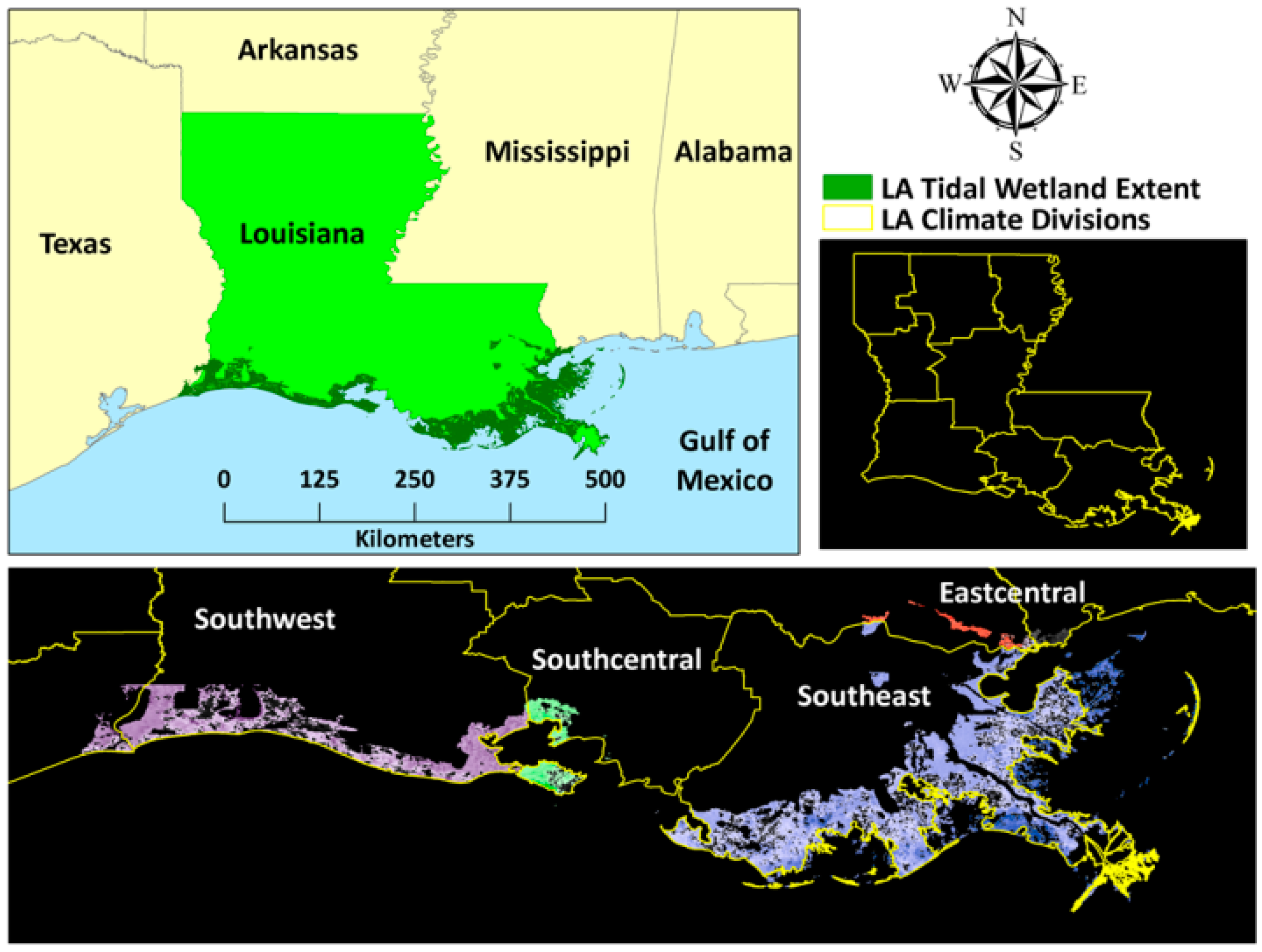
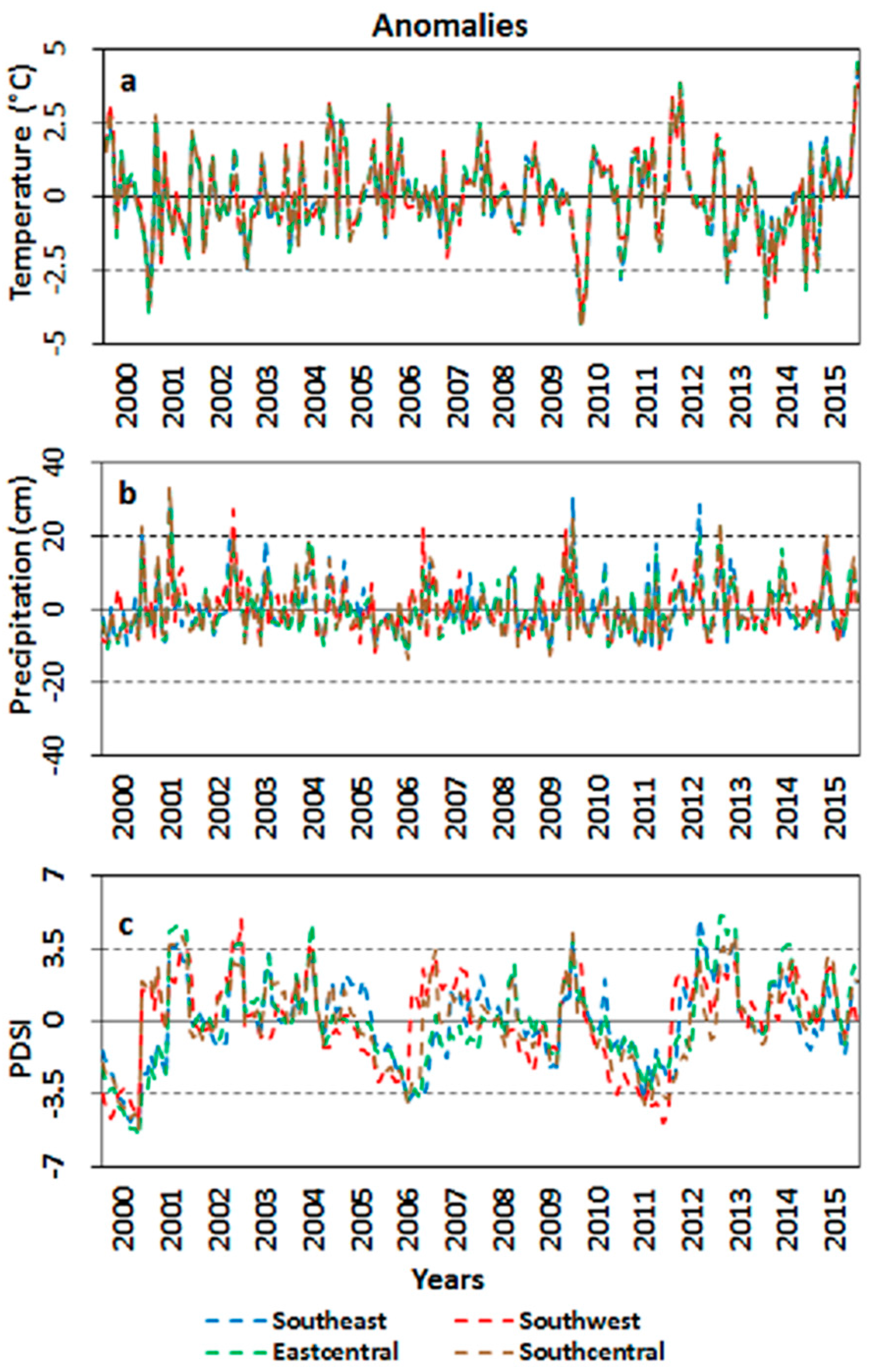
2.1. Study Area
2.2. Time-Series Composites
2.3. TIMESAT
2.4. Determination of Start and End of Season Determination Using TIMESAT and Derivative Analysis
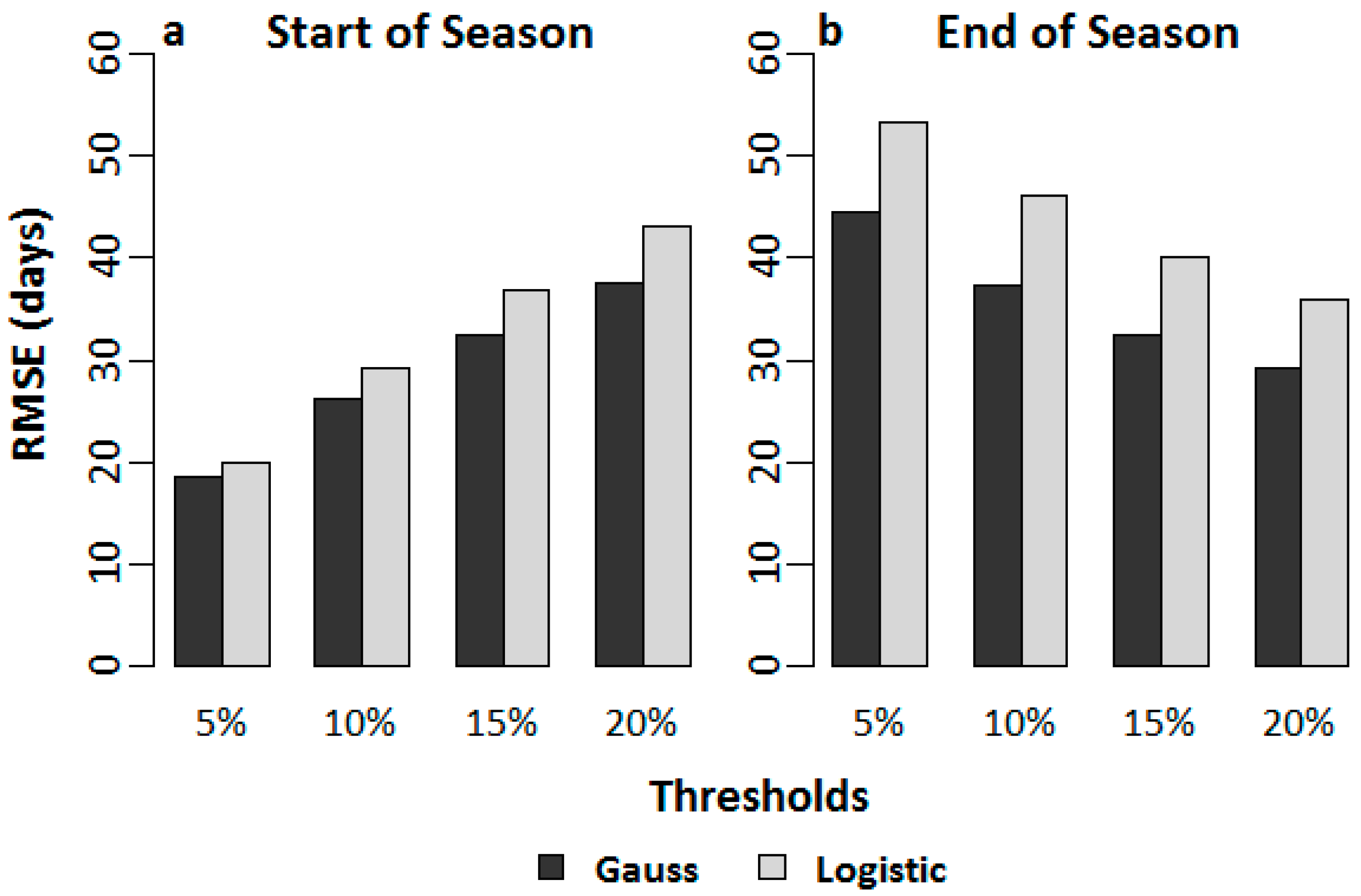
2.5. Analysis of SOS and EOS Fluctuations for the Validation of Thresholds
2.6. Seasonality Analysis and Simple Linear Trend Analysis
2.7. Trend Analysis
3. Results
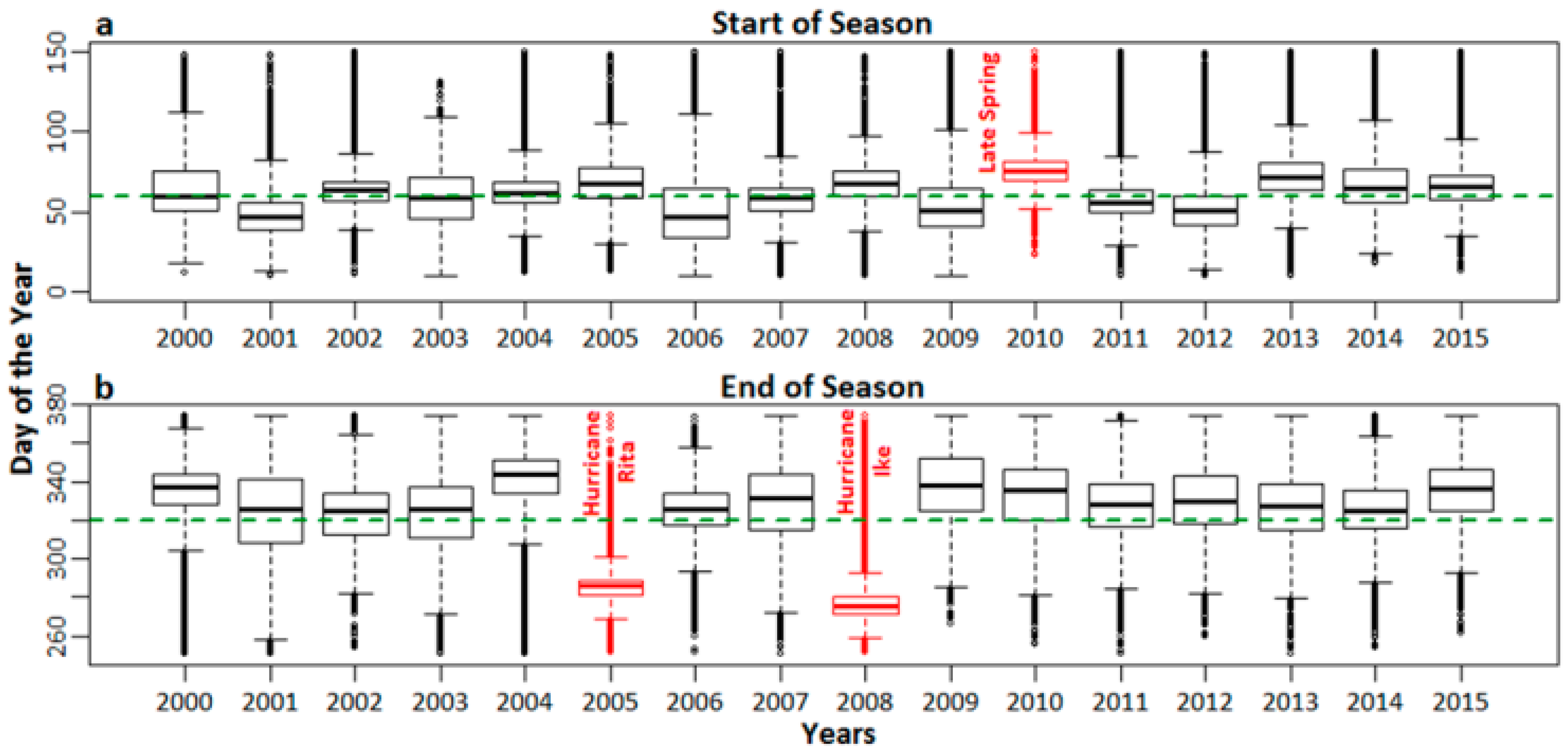
3.1. SOS and EOS Determination
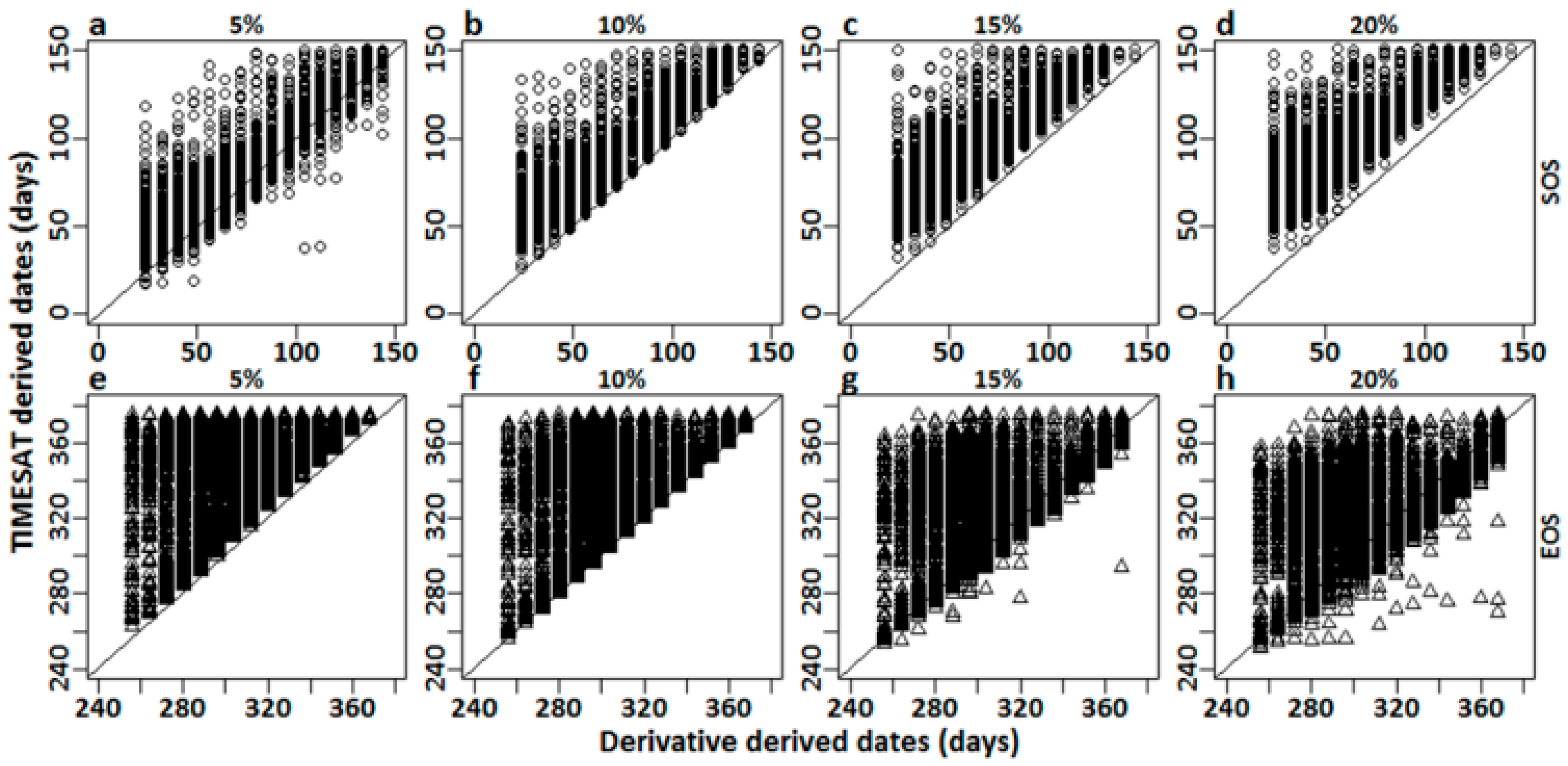
3.1.1. Derivative Analysis versus TIMESAT
3.1.2. SOS and EOS Validation through Analysis of Fluctuations
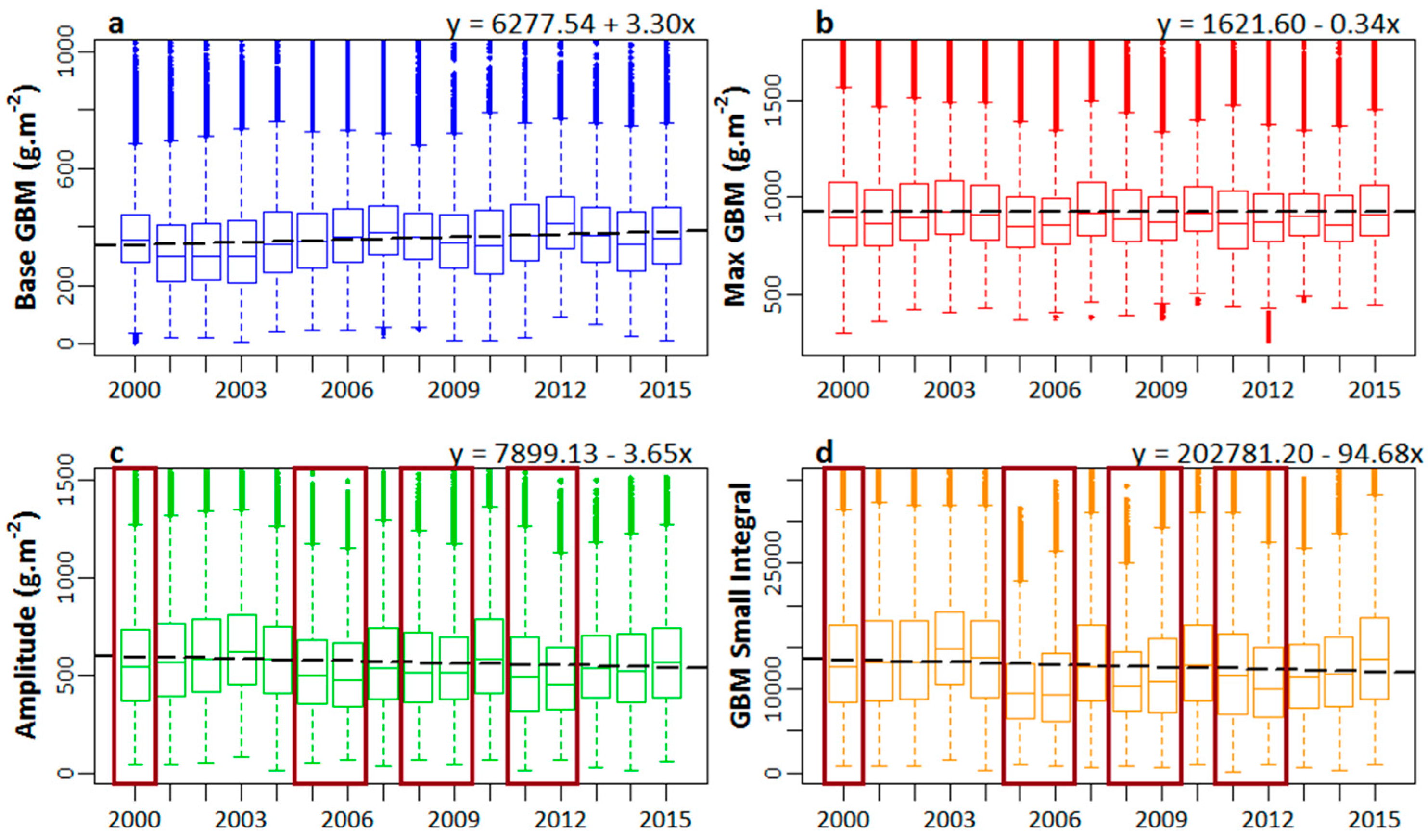
3.2. Seasonality Analysis and Simple Linear Trends
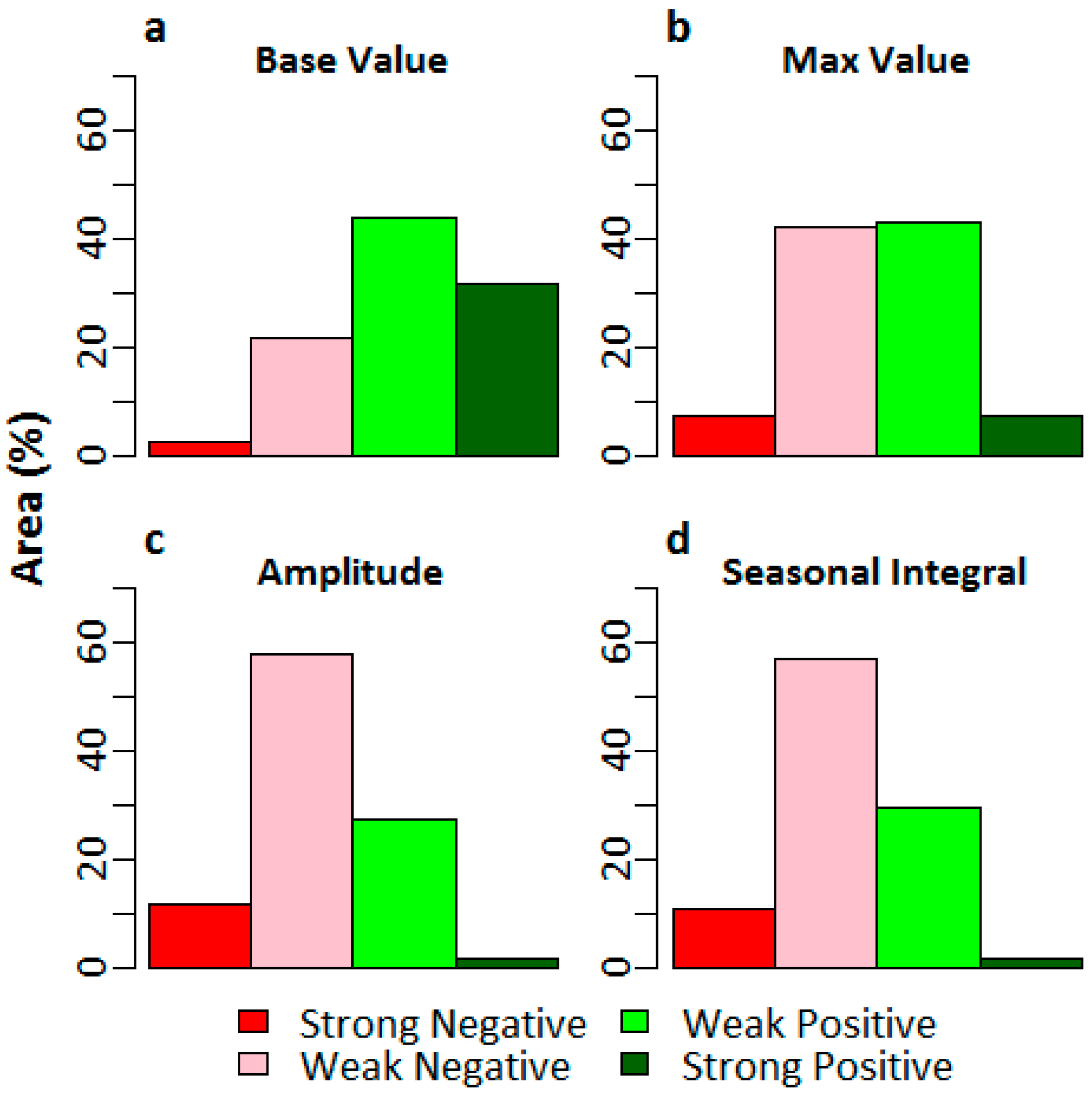
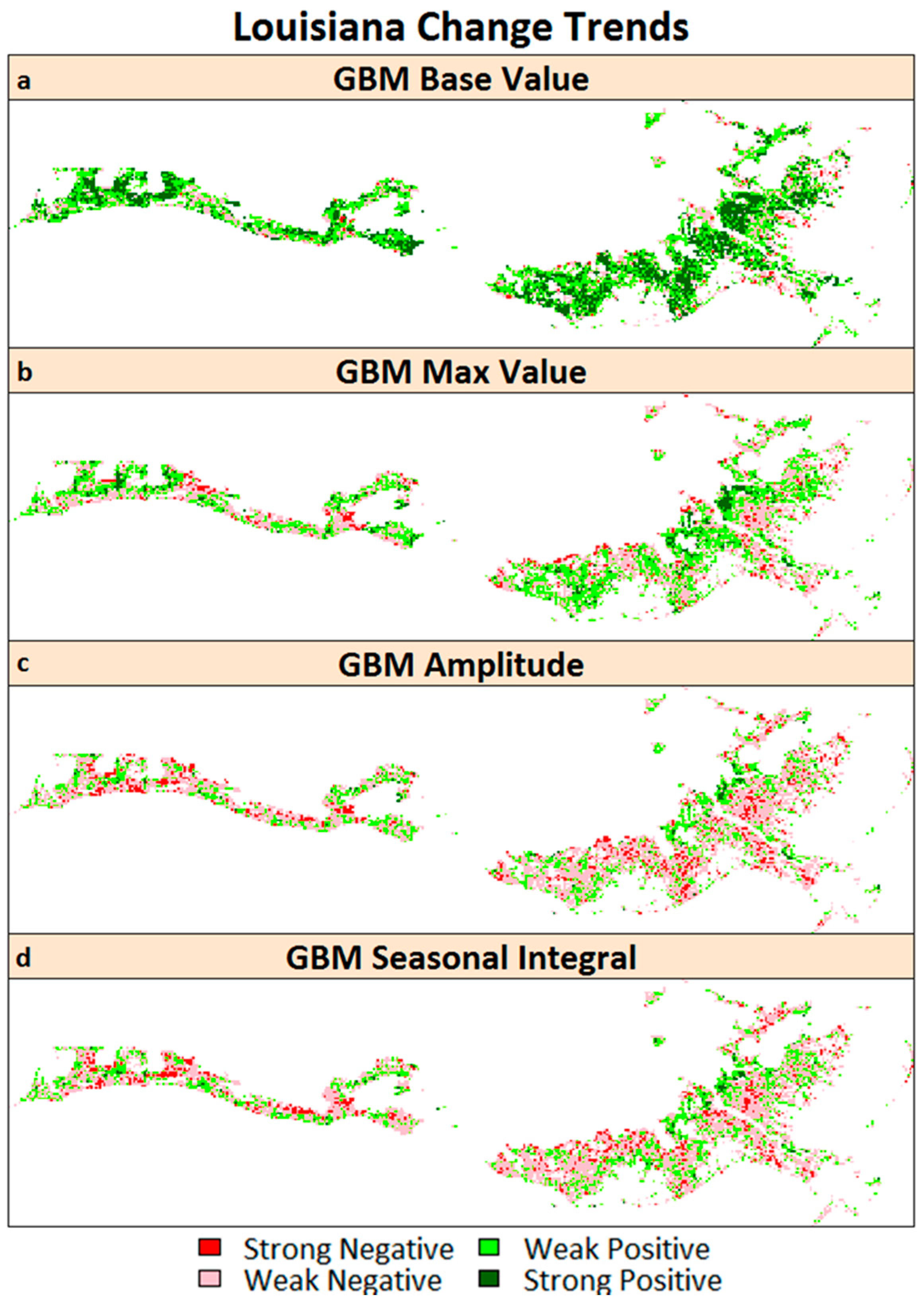
3.3. Mann-Kendall Trend Analysis
4. Discussion
4.1. SOS and EOS Determination
4.1.1. Derivative Analysis vs. TIMESAT
4.1.2. SOS and EOS Validation through Analysis of Fluctuations
4.2. Seasonality Analysis and Simple Linear Trends
4.3. Mann-Kendall Trend Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- NOAA Earth System Research Laboratory, Global Monitoring Division. Trends in Atmospheric Carbon Dioxide. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/graph (accessed on 25 December 2016).
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm–Kubler, K.; Bissolli, P.; Braslavska, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Post, E.; Forchhammer, M.C.; Bret-Harte, M.S.; Callaghan, T.V.; Christensen, T.R.; Elberling, B.; Fox, A.D.; Gilg, O.; Hik, D.S.; Høye, T.T.; et al. Ecological dynamics across the Arctic associated with recent climate change. Science 2009, 325, 1355–1358. [Google Scholar] [CrossRef] [PubMed]
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef] [PubMed]
- Connor, R.F.; Chmura, G.L.; Beecher, C.B. Carbon accumulation in Bay of Fundy salt marshes, Implications for restoration of reclaimed marshes. Glob. Biogeochem. Cycles 2001, 15, 943–954. [Google Scholar] [CrossRef]
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global carbon sequestration in tidal, saline wetland soils. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Mayor, J.R.; Hicks, C.E. Potential impacts of elevated CO2 on plant interactions, sustained growth, and carbon cycling in salt marsh ecosystems. In Human Impacts on Salt Marshes, a Global Perspective; Silliman, B.R., Grosholz, E.D., Bertness, M.D., Eds.; University of California Press: Berkeley, CA, USA, 2009; pp. 207–230. [Google Scholar]
- Erickson, J.E.; Megonigal, J.P.; Peresta, G.; Drake, B.G. Salinity and sea level mediate elevated CO2 effects on C3–C4 plant interactions and tissue nitrogen in a Chesapeake Bay tidal wetland. Glob. Chang. Biol. 2007, 13, 202–215. [Google Scholar] [CrossRef]
- White, K.P.; Langley, J.A.; Cahoon, D.R.; Megonigal, J.P. C3 and C4 biomass allocation responses to elevated CO2 and nitrogen, contrasting resource capture strategies. Estuaries Coasts 2012, 35, 1028–1035. [Google Scholar] [CrossRef]
- Polley, H.W.; Dugas, W.A.; Mielnick, P.C.; Johnson, H.B. C3–C4 composition and prior carbon dioxide treatment regulate the response of grassland carbon and water fluxes to carbon dioxide. Funct. Ecol. 2007, 21, 11–18. [Google Scholar] [CrossRef]
- Ghosh, S.; Mishra, D.R.; Gitelson, A.A. Long-term monitoring of biophysical characteristics of tidal wetlands in the northern Gulf of Mexico—A methodological approach using MODIS. Remote Sens. Environ. 2016, 173, 39–58. [Google Scholar] [CrossRef]
- Kramer, K.; Leinonen, I.; Loustau, D. The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems, an overview. Int. J. Biometeorol. 2000, 44, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Beaubien, E.G.; Freeland, H.J. Spring phenology trends in Alberta, Canada, links to ocean temperature. Int. J. Biometeorol. 2000, 44, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Primack, D.; Imbres, C.; Primack, R.B.; Miller-Rushing, A.J.; Del Tredici, P. Herbarium specimens demonstrate earlier flowering times in response to warming in Boston. Am. J. Bot. 2004, 91, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Keatinge, J.D.H.; Qi, A.; Wheeler, T.R.; Ellis, R.H.; Summerfield, R.J. Effects of temperature and photoperiod on phenology as a guide to the selection of annual legume cover and green manure crops for hillside farming systems. Field Crop. Res. 1998, 57, 139–152. [Google Scholar] [CrossRef]
- Hartkamp, A.D.; Hoogenboom, G.; White, J.W. Adaptation of the CROPGRO growth model to velvet bean (Mucuna pruriens), II. Cultivar evaluation and model development. Field Crop. Res. 2002, 78, 9–25. [Google Scholar] [CrossRef]
- Roetzer, T.; Wittenzeller, M.; Haeckel, H.; Nekovar, J. Phenology in central Europe–differences and trends of spring phenophases in urban and rural areas. Int. J. Biometeorol. 2000, 44, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H. Climate controls on vegetation phenological patterns in northern mid-and high latitudes inferred from MODIS data. Glob. Chang. Biol. 2004, 10, 1133–1145. [Google Scholar] [CrossRef]
- Keeling, C.D.; Chin, F.J.S.; Whorf, T.P. Increased activity of northern vegetation inferred from atmospheric CO2 measurements. Nature 1996, 382, 146–149. [Google Scholar] [CrossRef]
- Asner, G.P.; Townsend, A.R.; Braswell, B.H. Satellite observation of El Nino effects on Amazon forest phenology and productivity. Geophys. Res. Lett. 2000, 27, 981–984. [Google Scholar] [CrossRef]
- Vicente–Serrano, S.M. Spatial and temporal analysis of droughts in the Iberian Peninsula 1910–2000. Hydrol. Sci. J. 2006, 51, 83–97. [Google Scholar] [CrossRef]
- Van Vliet, A.J.; Overeem, A.; De Groot, R.S.; Jacobs, A.F.; Spieksma, F. The influence of temperature and climate change on the timing of pollen release in the Netherlands. Int. J. Climatol. 2002, 22, 1757–1767. [Google Scholar] [CrossRef]
- Arora, V.K. The use of the aridity index to assess climate change effect on annual runoff. J. Hydrol. 2002, 265, 164–177. [Google Scholar] [CrossRef]
- Running, S.W.; Nemani, R.R. Regional hydrologic and carbon balance responses of forests resulting from potential climate change. Clim. Chang. 1991, 19, 349–368. [Google Scholar] [CrossRef]
- Wilson, K.B.; Baldocchi, D.D. Seasonal and inter–annual variability of energy fluxes over a broadleaved temperate deciduous forest in North America. Agric. For. Meteorol. 2000, 100, 1–18. [Google Scholar] [CrossRef]
- Kunkel, K.E.; Easterling, D.R.; Hubbard, K.; Redmond, K. Temporal variations in frost-free season in the United States, 1895–2000. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Scheifinger, H.; Menzel, A.; Koch, E.; Peter, C.; Ahas, R. Atmospheric mechanisms governing the spatial and temporal variability of phenological phases in central Europe. Int. J. Climatol. 2002, 22, 1739–1755. [Google Scholar] [CrossRef]
- Soudani, K.; Le Maire, G.; Dufrêne, E.; François, C.; Delpierre, N.; Ulrich, E.; Cecchini, S. Evaluation of the onset of green–up in temperate deciduous broadleaf forests derived from Moderate Resolution Imaging Spectroradiometer (MODIS) data. Remote Sens. Environ. 2008, 112, 2643–2655. [Google Scholar] [CrossRef]
- Zhang, X.; Goldberg, M.D. Monitoring fall foliage coloration dynamics using time–series satellite data. Remote Sens. Environ. 2011, 115, 382–391. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.; Gao, F.; Reed, B.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Wolfe, R.E.; Nishihama, M.; Fleig, A.J.; Kuyper, J.A.; Roy, D.P.; Storey, J.C.; Patt, F.S. Achieving sub–pixel geolocation accuracy in support of MODIS land science. Remote Sens. Environ. 2002, 83, 31–49. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Ahl, D.E.; Gower, S.T.; Burrows, S.N.; Shabanov, N.V.; Myneni, R.B.; Knyazikhin, Y. Monitoring spring canopy phenology of a deciduous broadleaf forest using MODIS. Remote Sens. Environ. 2006, 104, 88–95. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F. Cross–scalar satellite phenology from ground, Landsat, and MODIS data. Remote Sens. Environ. 2007, 109, 261–273. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B. Global vegetation phenology from Moderate Resolution Imaging Spectroradiometer (MODIS), Evaluation of global patterns and comparison with in situ measurements. J. Geophys. Res. Biogeosci. 2006, 111. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. A statistical framework for the analysis of long image time series. Int. J. Remote Sens. 2005, 26, 1551–1573. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Spatio–temporal statistical methods for modelling land surface phenology. In Phenological Research: Methods for Environmental and Climate Change Analysis; Hudson, I.L., Keatley, M.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 177–208. ISBN 9789048133352. [Google Scholar]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’keefe, J.; Zhang, G.; Nemani, R.R.; Leeuwen, V. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Walker, J.J.; De Beurs, K.M.; Wynne, R.H.; Gao, F. Evaluation of Landsat and MODIS data fusion products for analysis of dryland forest phenology. Remote Sens. Environ. 2012, 117, 381–393. [Google Scholar] [CrossRef]
- Verbesselt, J.; Hyndman, R.; Zeileis, A.; Culvenor, D. Phenological change detection while accounting for abrupt and gradual trends in satellite image time series. Remote Sens. Environ. 2010, 114, 2970–2980. [Google Scholar] [CrossRef]
- Gutman, G.G. Vegetation indices from AVHRR: An update and future prospects. Remote Sens. Environ. 1991, 35, 121–136. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q. An error and sensitivity analysis of the atmospheric and soil–correcting variants of the NDVI for the MODIS–EOS. IEEE Trans. Geosci. Remote Sens. 1994, 32, 897–905. [Google Scholar] [CrossRef]
- Xiao, X.; Braswell, B.; Zhang, Q.; Boles, S.; Frolking, S.; Moore, B. Sensitivity of vegetation indices to atmospheric aerosols, continental–scale observations in Northern Asia. Remote Sens. Environ. 2003, 84, 385–392. [Google Scholar] [CrossRef]
- Atkinson, P.M.; Jeganathan, C.; Dash, J.; Atzberger, C. Inter-comparison of four models for smoothing satellite sensor time–series data to estimate vegetation phenology. Remote Sens. Environ. 2012, 123, 400–417. [Google Scholar] [CrossRef]
- Viovy, N.; Arino, O.; Belward, A.S. The Best Index Slope Extraction (BISE): A method for reducing noise in NDVI time–series. Int. J. Remote Sens. 1992, 13, 1585–1590. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time–series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Chen, X.; Hu, B.; Yu, R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Glob. Chang. Biol. 2005, 11, 1118–1130. [Google Scholar] [CrossRef]
- Moody, E.G.; King, M.D.; Platnick, S.; Schaaf, C.B.; Gao, F. Spatially complete global spectral surface albedos, Value–added datasets derived from Terra MODIS land products. IEEE Trans. Geosci. Remote Sens. 2005, 43, 144–158. [Google Scholar] [CrossRef]
- Sellers, P.J.; Tucker, C.J.; Collatz, G.J.; Los, S.O.; Justice, C.O.; Dazlich, D.A.; Randall, D.A. A global 1° by 1° NDVI data set for climate studies, Part II. The generation of global fields of terrestrial biophysical parameters from the NDVI. Int. J. Remote Sens. 1994, 15, 3519–3545. [Google Scholar] [CrossRef]
- Roerink, G.J.; Menenti, M.; Verhoef, W. Reconstructing cloudfree NDVI composites using Fourier analysis of time series. Int. J. Remote Sens. 2000, 21, 1911–1917. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. TIMESAT [Software], Lund University, Lund, Sweden. 2002. Available online: http://web.nateko.lu.se/TIMESAT/TIMESAT.asp (accessed on 1 May 2016).
- Jonsson, P.; Eklundh, L. TIMESAT—A program for analyzing time–series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jonsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Asner, G.P.; Alencar, A. Drought impacts on the Amazon forest, the Rem. Sens perspective. New Phytol. 2010, 187, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite Remote Sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Chen, J.; Zhu, X.; Zheng, Y. Influences of temperature and precipitation before the growing season on spring phenology in grasslands of the central and eastern Qinghai–Tibetan Plateau. Agric. For. Meteorol. 2011, 151, 1711–1722. [Google Scholar] [CrossRef]
- Palmer, S.C.; Odermatt, D.; Hunter, P.D.; Brockmann, C.; Presing, M.; Balzter, H.; Tóth, V.R. Satellite Remote Sensing of phytoplankton phenology in Lake Balaton using 10 years of MERIS observations. Remote Sens. Environ. 2015, 158, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Mishra, D.R.; Ghosh, S. Using moderate resolution satellite sensors for monitoring the biophysical parameters and phenology of tidal wetlands. In Remote Sensing of Wetlands, Applications and Advances; Tiner, R., Land, M., Klemas, V., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 283–314. ISBN 9781482237351. [Google Scholar]
- Mishra, D.R.; Ghosh, S.; Hladik, C.; O’Connell, J.L.; Cho, H.J. Wetland mapping methods and techniques using multi-sensor, multi-resolution Remote Sensing, Successes and Challenges. In Remote Sensing Handbook; Thenkabail, P.S., Ed.; CRC Press: Boca Raton, FL, USA, 2015; Volume III, pp. 191–227. ISBN 9781482218015. [Google Scholar]
- Mo, Y.; Kearney, M.; Momen, B. Drought-associated phenological changes of coastal marshes in Louisiana. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- United States Fish and Wildlife Service, National Wetlands Inventory. Available online: https://www.fws.gov/wetlands/Data/Data–Download (accessed on 1 December 2016).
- Weis, J.S. Salt marsh. In Encyclopedia of Earth; Cleveland, C.J., Ed.; Environmental Information Coalition, National Council for Science and the Environment: Washington, DC, USA, 2010. [Google Scholar]
- National Oceanic and Atmospheric Administration National Centers for Environmental Information, Climate at a Glance, U.S. Time Series. Available online: http://www.ncdc.noaa.gov/cag/ (accessed on 31 December 2016).
- National Oceanic and Atmospheric Administration National Weather Service. Available online: http://www.weather.gov/ (accessed on 21 September 2016).
- National Oceanic and Atmospheric Administration National Hurricane Centre, NHC Data Archive. Available online: http://www.nhc.noaa.gov/data/ (accessed on 30 June 2016).
- Lyles, L.D.; Namwamba, F.; Campus, B.R. Louisiana coastal zone erosion, 100+ years of land use and land loss using GIS and Remote Sensing. In Proceedings of the 5th Annual ESRI Education User Conference, San Diego, CA, USA, 23–26 July 2005; pp. 23–26. [Google Scholar]
- Tiner, R. Tidal Wetlands Primer, an Introduction to Their Ecology, Natural History, Status, and Conservation; Tiner, R., Ed.; University of Massachusetts Press: Amherst, MA, USA, 2013; ISBN 9781625340290. [Google Scholar]
- Bertness, M.D.; Silliman, B.R.; Holdredge, C. Shoreline development and the future of New England salt marsh landscapes. In Human Impacts on Salt Marshes, A Global Perspective; Silliman, B.R., Grosholz, E.D., Bertness, M.D., Eds.; University of California Press: Berkeley, CA, USA, 2009; pp. 137–148. ISBN 9780520258921. [Google Scholar]
- Lindstedt, D.M.; Swenson, E.M. The Case of the Dying Marsh Grass; Report submitted to Louisiana Department of natural Resources: Baton Rouge, LA, USA, 2006; pp. 1–19. [Google Scholar]
- Biber, P.D.; Wu, W.; Peterson, M.S.; Liu, Z.; Pham, L. Oil contamination in Mississippi salt marsh habitats and the impacts to Spartina alterniflora photosynthesis. In Impacts of Oil Spill Disasters on Marine Habitats and Fisheries in North America; Alford, J.B., Peterson, M.S., Green, C.G., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 133–172. ISBN 9781466557208. [Google Scholar]
- Mishra, D.R.; Cho, H.J.; Ghosh, S.; Fox, A.; Downs, C.; Merani, P.B.T.; Kirui, P.; Jackson, N.; Mishra, S. Post–spill state of the marsh, Remote estimation of the ecological impact of the Gulf of Mexico oil spill on Louisiana Salt Marshes. Remote Sens. Environ. 2012, 118, 176–185. [Google Scholar] [CrossRef]
- Vermote, E.F.; Kotchenova, S.Y.; Ray, J.P. MODIS Surface Reflectance User’s Guide; Version 1; MODIS Land Surface Reflectance Science Computing Facility: Greenbelt, MD, USA, 2011; pp. 1–35. [Google Scholar]
- Gitelson, A.A.; Stark, R.; Grits, U.; Rundquist, D.; Kaufman, Y.; Derry, D. Vegetation and soil lines in visible spectral space, a concept and technique for remote estimation of vegetation fraction. Int. J. Remote Sens. 2002, 23, 2537–2562. [Google Scholar] [CrossRef]
- Mo, Y.; Momen, B.; Kearney, M.S. Quantifying moderate resolution remote sensing phenology of Louisiana coastal marshes. Ecol. Model. 2015, 312, 191–199. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological drought. In Weather Bureau Research Paper No. 45; U.S. Department of Commerce: Washington, DC, USA, 1965; pp. 1–58. [Google Scholar]
- Andres, L.; Salas, W.A.; Skole, D. Fourier analysis of multi–temporal AVHRR data applied to a land cover classification. Int. J. Remote Sens. 1994, 15, 1115–1121. [Google Scholar] [CrossRef]
- Olsson, L.; Eklundh, L. Fourier series for analysis of temporal sequences of satellite sensor imagery. Int. J. Remote Sens. 1994, 15, 3735–3741. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Lauenroth, W.K. Interannual variability of NDVI and its relationship to climate for North American shrublands and grasslands. J. Biogeogr. 1998, 25, 721–733. [Google Scholar] [CrossRef]
- Eklundh, L.; Jonsson, P. TIMESAT 3.2 with Parallel Processing—Software Manual; Lund University: Lund, Sweden, 2015; pp. 1–88. [Google Scholar]
- Gao, F.; Morisette, J.T.; Wolfe, R.E.; Ederer, G.; Pedelty, J.; Masuoka, E.; Myneni, R.; Tan, B.; Nightingale, J. An algorithm to produce temporally and spatially continuous MODIS–LAI time series. IEEE Geosci. Remote Sens. Lett. 2008, 5, 60–64. [Google Scholar] [CrossRef]
- Tan, B.; Morisette, J.T.; Wolfe, R.E.; Gao, F.; Ederer, G.A.; Nightingale, J.; Pedelty, J.A. An enhanced TIMESAT algorithm for estimating vegetation phenology metrics from MODIS data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2011, 4, 361–371. [Google Scholar] [CrossRef]
- Cleveland, R.B.; Cleveland, W.S.; Terpenning, I. STL, a seasonal–trend decomposition procedure based on loess. J. Off. Stat. 1990, 6, 3–73. [Google Scholar]
- Suepa, T.; Qi, J.; Lawawirojwong, S.; Messina, J.P. Understanding spatio–temporal variation of vegetation phenology and rainfall seasonality in the monsoon Southeast Asia. Environ. Res. 2016, 147, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Jia, G.; Epstein, H. Recent changes in phenology over the northern high latitudes detected from multi–satellite data. Environ. Res. Lett. 2011, 6. [Google Scholar] [CrossRef]
- Delbart, N.; Kergoat, L.; Le Toan, T.; Lhermitte, J.; Picard, G. Determination of phenological dates in boreal regions using normalized difference water index. Remote Sens. Environ. 2005, 97, 26–38. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Griffin: London, UK, 1970; pp. 1–202. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R–project.org/ (accessed on 15 August 2016).
- Alber, M.; Swenson, E.M.; Adamowicz, S.C.; Mendelssohn, I.A. Salt marsh dieback: An overview of recent events in the US. Estuar. Coast. Shelf Sci. 2008, 80, 1–11. [Google Scholar] [CrossRef]
- Khanna, S.; Santos, M.J.; Ustin, S.L.; Koltunov, A.; Kokaly, R.F.; Roberts, D.A. Detection of salt marsh vegetation stress after the Deepwater Horizon BP oil spill along the shoreline of Gulf of Mexico using AVIRIS data. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Santos, M.J.; Koltunov, A.; Shapiro, K.D.; Lay, M.; Ustin, S.L. Marsh Loss due to cumulative impacts of Hurricane Isaac and the Deepwater Horizon Oil Spill in Louisiana. Remote Sens. 2017, 9, 169. [Google Scholar] [CrossRef]
- Costanza, R.; Mitsch, W.J.; Day, J.W. A new vision for New Orleans and the Mississippi delta, applying ecological economics and ecological engineering. Front. Ecol. Environ. 2006, 4, 465–472. [Google Scholar] [CrossRef]
- McKee, K.L.; Mendelssohn, I.A.; Materne, M.D. Salt marsh dieback in coastal Louisiana: Survey of plant and soil conditions in Barataria and Terrebonne Basins, June 2000–September 2001. U.S. Geological Survey Open-File Report 2006-1167; Submitted to United States of Geological Survey; 2006. Available online: https://pubs.usgs.gov/of/2006/1167/ (accessed on 25 June 2016).
- Ramsey, E.; Rangoonwala, A.; Chi, Z.; Jones, C.E.; Bannister, T. Marsh dieback, loss, and recovery mapped with satellite optical, airborne polarimetric radar, and field data. Remote Sens. Environ. 2014, 152, 364–374. [Google Scholar] [CrossRef]
- Day, J.W.; Boesch, D.F.; Clairain, E.J.; Kemp, G.P.; Laska, S.B.; Mitsch, W.J.; Orth, K.; Mashriqui, H.; Reed, D.J.; Shabman, L.; et al. Restoration of the Mississippi Delta: Lessons from hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar] [CrossRef] [PubMed]
- Barras, J.A. Satellite Images and Aerial Photographs of the Effects of Hurricanes Katrina and Rita on Coastal Louisiana: U.S. Geological Survey Data Series 281. 2007. Available online: https://pubs.usgs.gov/ds/2007/281/ (accessed on 10 January 2016).
- Barras, J.A.; Brock, J.C.; Morton, R.A.; Travers, L.J. Remotely sensed imagery revealing the effects of hurricanes Gustav and Ike on coastal Louisiana: U.S. Geological Survey Data Series 566. 2010. Available online: https://pubs.usgs.gov/ds/566/ (accessed on 21 January 2016).
- Guy, K.K.; Stockdon, H.F.; Plant, N.G.; Doran, K.S.; Morgan, K.L.M. Hurricane Isaac: Observations and Analysis of Coastal Change. U.S. Geological Survey Open-File Report 2013-1270; Submitted to United States of Geological Survey; 2013. Available online: https://pubs.usgs.gov/of/2013/1270/ (accessed on 18 February 2016).
- Song, Q.; Zhang, G.; Zhu, X.G. Optimal crop canopy architecture to maximize canopy photosynthetic CO2 uptake under elevated CO2: A theoretical study using a mechanistic model of canopy photosynthesis. Funct. Plant Biol. 2013, 40, 108–124. [Google Scholar] [CrossRef]
- Riddin, T.; Adams, J.B. The effect of a storm surge event on the macrophytes of a temporarily open/closed estuary, South Africa. Estuar. Coast. Shelf Sci. 2010, 89, 119–123. [Google Scholar] [CrossRef]
- NOAA Center for Operational Oceanographic Products and Services, NOAA Tides and Currents. Sea Level Trends. Available online: https://tidesandcurrents.noaa.gov/sltrends/sltrends (accessed on 21 January 2017).
- Taiz, L.; Zeiger, E. Plant Physiology; Sinauer Associates Inc.: Sunderland, MA, USA, 2015; pp. 1–559. [Google Scholar]
- Leakey, A.D. Rising atmospheric carbon dioxide concentration and the future of C4 crops for food and fuel. Proc. R. Soc. B 2009, 276, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- DeLaune, R.D.; White, J.R. Will coastal wetlands continue to sequester carbon in response to an increase in global sea level? A case study of the rapidly subsiding Mississippi river deltaic plain. Clim. Chang. 2012, 110, 297–314. [Google Scholar] [CrossRef]
- Howard, R.J.; Travis, S.E.; Sikes, B.A. Rapid growth of a Eurasian haplotype of Phragmites australis in a restored brackish marsh in Louisiana, USA. Biol. Invasions 2008, 10, 369–379. [Google Scholar] [CrossRef]
- Antonielli, M.; Pasqualini, S.; Batini, P.; Ederli, L.; Massacci, A.; Loreto, F. Physiological and anatomical characterization of Phragmites australis leaves. Aquat. Bot. 2002, 72, 55–66. [Google Scholar] [CrossRef]
- Bertness, M.D.; Ewanchuk, P.J.; Silliman, B.R. Anthropogenic modification of New England salt marsh landscapes. Proc. Natl. Acad. Sci. USA 2002, 99, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Chambers, R.M.; Osgood, D.T.; Bart, D.J.; Montalto, F. Phragmites australis invasion and expansion in tidal wetlands, interactions among salinity, sulfide, and hydrology. Estuaries Coasts 2003, 26, 398–406. [Google Scholar] [CrossRef]
- Minchinton, T.E.; Bertness, M.D. Disturbance-mediated competition and the spread of Phragmites australis in a coastal marsh. Ecol. Appl. 2003, 13, 1400–1416. [Google Scholar] [CrossRef]
- Silliman, B.R.; Bertness, M.D. Shoreline development drives invasion of Phragmites australis and the loss of plant diversity on New England salt marshes. Conserv. Biol. 2004, 18, 1424–1434. [Google Scholar] [CrossRef]
- Pennings, S.C.; Silliman, B.R. Linking biogeography and community ecology, latitudinal variation in plant–herbivore interaction strength. Ecology 2005, 86, 2310–2319. [Google Scholar] [CrossRef]
- Windham, L.; Ehrenfeld, J.G. Net impact of a plant invasion on nitrogen-cycling processes within a brackish tidal marsh. Ecol. Appl. 2003, 13, 883–896. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seasonality Parameters | Description/Phenological Interpretation | Unit |
|---|---|---|
| Start of Season (SOS) | Time at the beginning of the growing season, when GBM begins to increase and photosynthesis starts | Julian days starting from January 1 |
| End of Season (EOS) | Time at the end of growing season, when GBM begins to decrease and photosynthesis stops completely | Julian days starting from January 1 |
| Peak of Season (POS) | Computed as the mean of the time period for which the green-up process stops and brown-down starts; time when the GBM and photosynthesis reach their maximum level | Julian days starting from January 1 |
| Length of Season (LOS) | Time from start to end of growing season | Days |
| Base Value | Mean of the minimum GBM values at the start (initial GBM) and end (final GBM) of the growing season | GBM unit |
| Max Value | Maximum GBM value for the fitted function during the growing season/ GBM value during the peak of the growing season | GBM unit |
| Amplitude | Difference between the base and max value; Maximum increase in canopy photosynthetic activity above the baseline | GBM unit |
| Left Derivative | Rate of green-up; rate of increase of GBM values from the beginning till the peak of the growing season | GBM unit/8 days |
| Right Derivative | Rate of brown-down; rate of decrease of GBM values from the peak to the end of the growing season | GBM unit/8 days |
| Small Seasonal Integral | Integral of the function describing the season from the start to end of the season, above the base level; indicator of net canopy photosynthetic rate across the entire growing season | GBM unit |
| Large Seasonal Integral | Integral of the function describing the season from start to end of the season; indicator of gross canopy photosynthetic rate across the entire growing season, along with the base GBM | GBM unit |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, S.; Mishra, D.R. Analyzing the Long-Term Phenological Trends of Salt Marsh Ecosystem across Coastal LOUISIANA. Remote Sens. 2017, 9, 1340. https://doi.org/10.3390/rs9121340
Ghosh S, Mishra DR. Analyzing the Long-Term Phenological Trends of Salt Marsh Ecosystem across Coastal LOUISIANA. Remote Sensing. 2017; 9(12):1340. https://doi.org/10.3390/rs9121340
Chicago/Turabian StyleGhosh, Shuvankar, and Deepak R. Mishra. 2017. "Analyzing the Long-Term Phenological Trends of Salt Marsh Ecosystem across Coastal LOUISIANA" Remote Sensing 9, no. 12: 1340. https://doi.org/10.3390/rs9121340
APA StyleGhosh, S., & Mishra, D. R. (2017). Analyzing the Long-Term Phenological Trends of Salt Marsh Ecosystem across Coastal LOUISIANA. Remote Sensing, 9(12), 1340. https://doi.org/10.3390/rs9121340