
Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA

Abstract
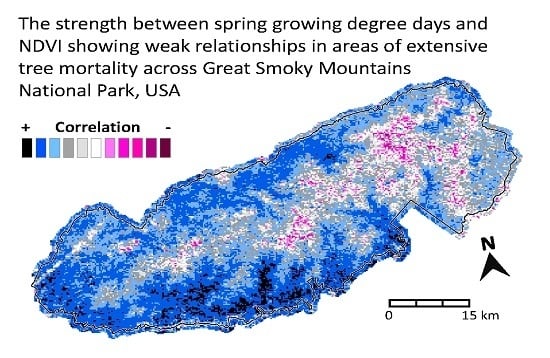
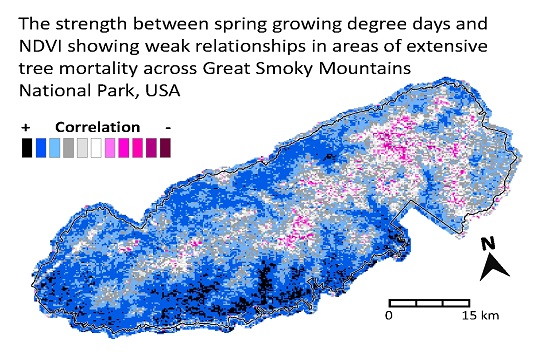
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Land Surface Phenology
2.3. Comparison with Independent Data
2.4. Pheno-Topographic Relationships
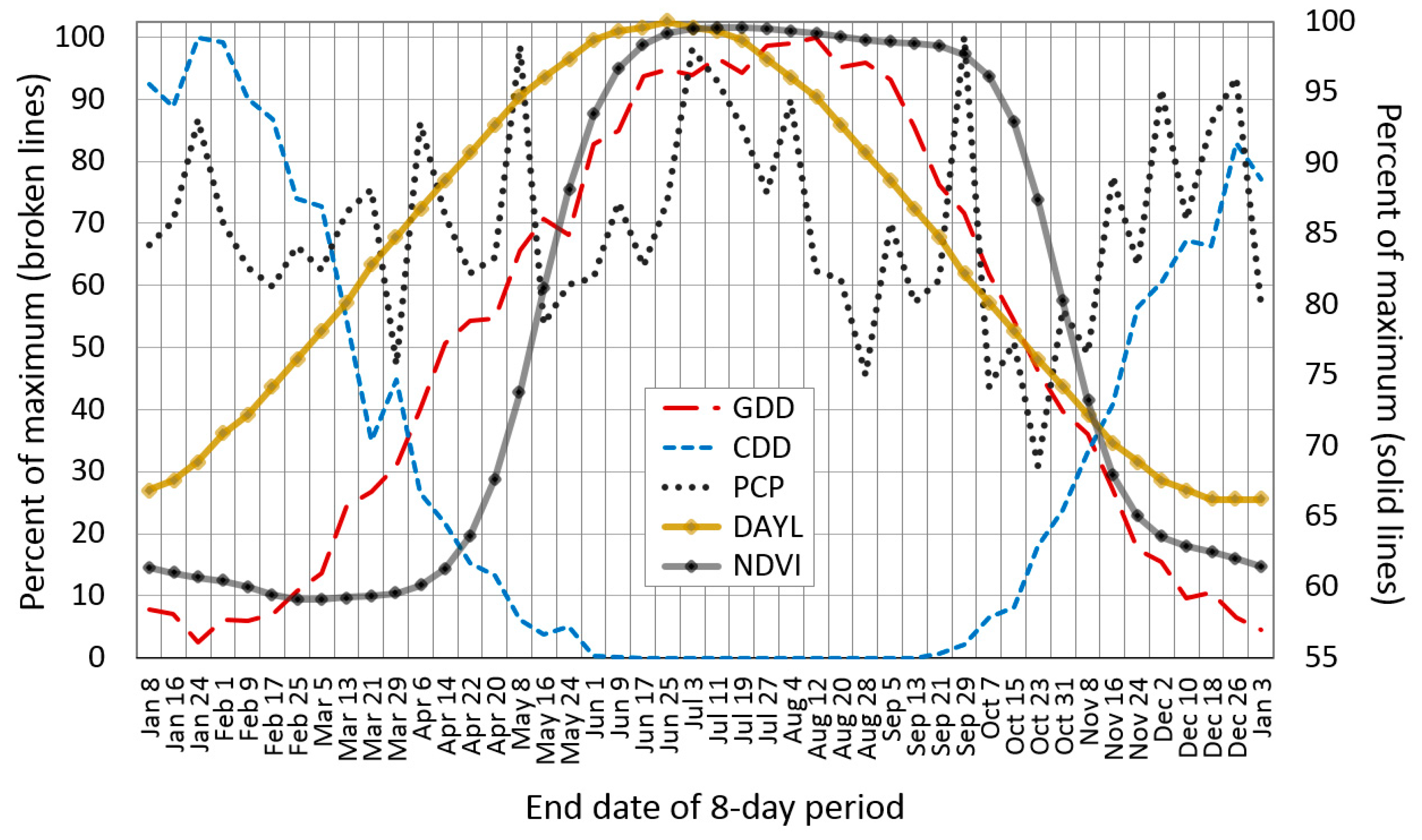
2.5. Pheno-Climatological Relationships
3. Results
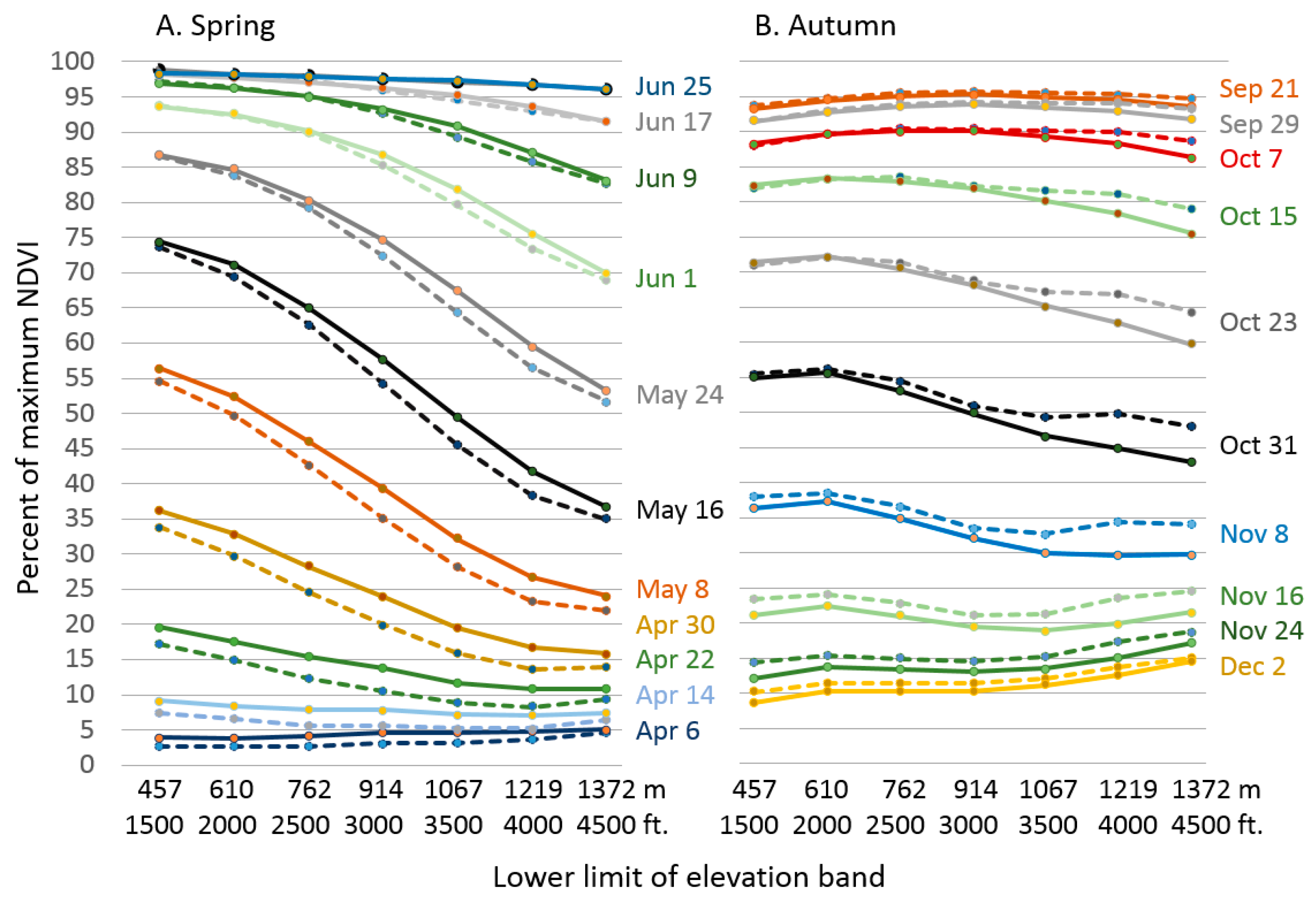
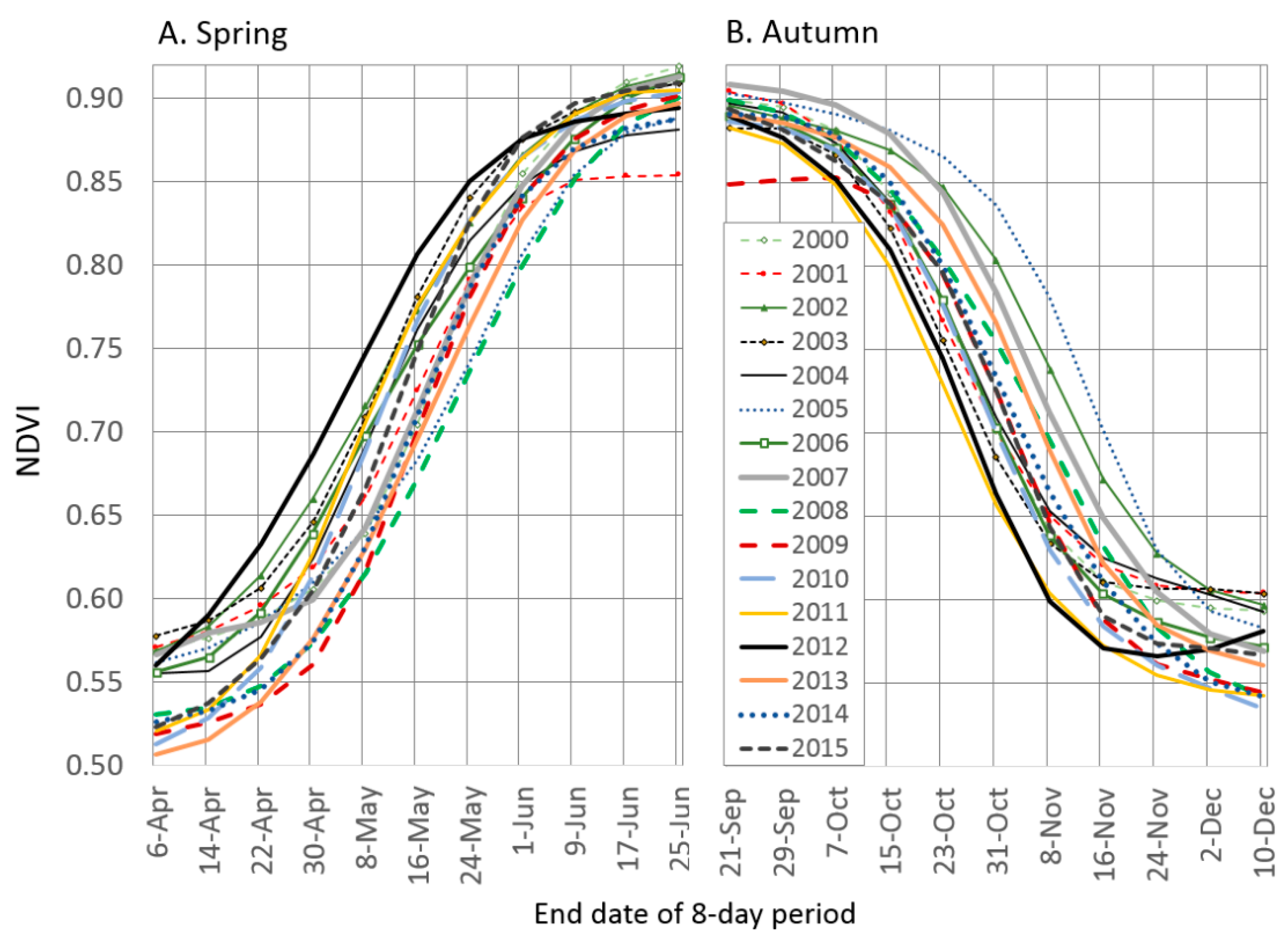
3.1. Land Surface Phenology
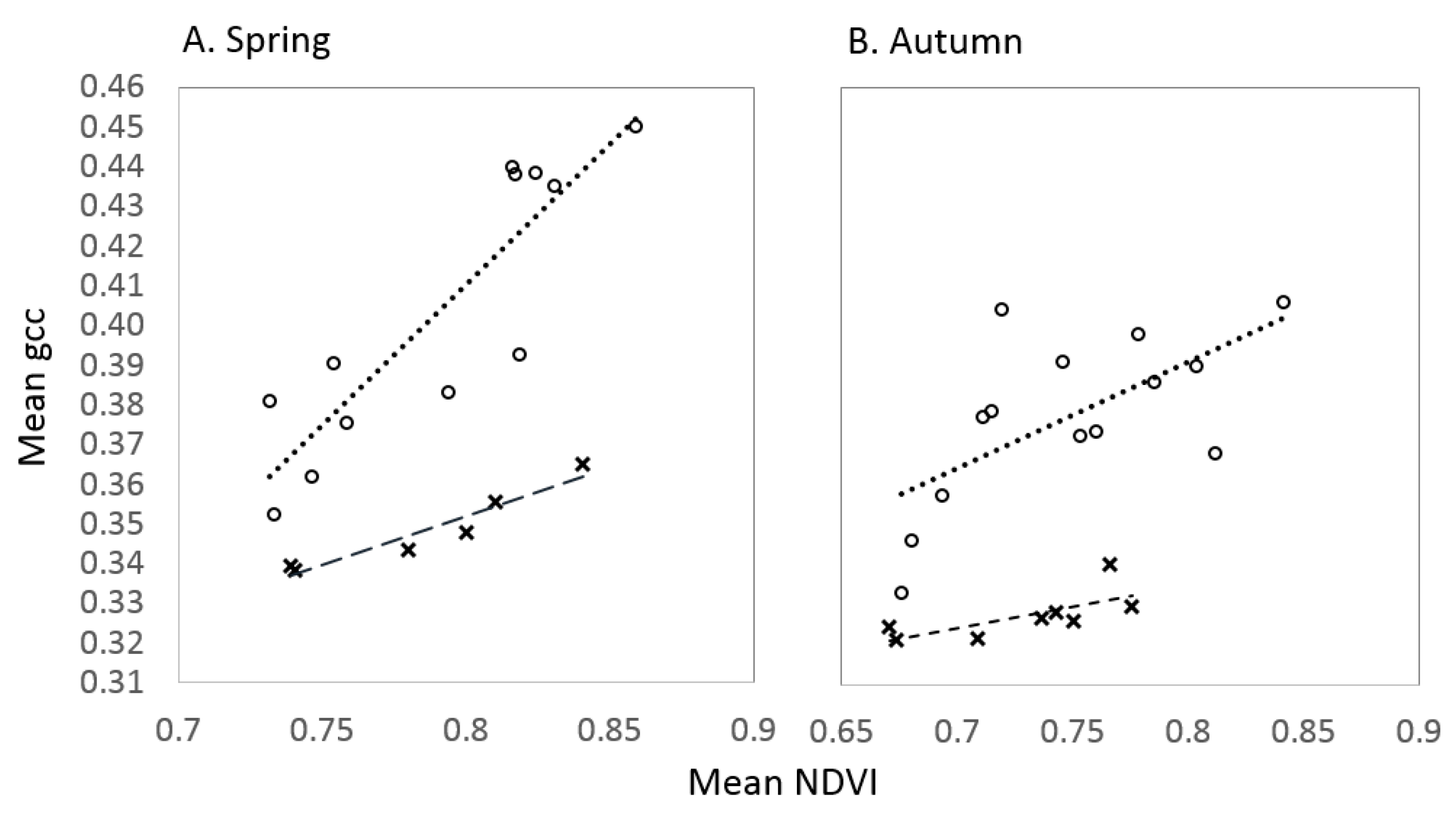
3.2. Comparison with Independent Data
3.3. Pheno-Climatology
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- White, K.; Pontius, J.; Schaberg, P. Remote sensing of spring phenology in northeastern forests: A comparison of methods, field metrics and sources of uncertainty. Remote Sens. Environ. 2014, 148, 97–107. [Google Scholar] [CrossRef]
- Norman, S.P.; Koch, F.H.; Hargrove, W.W. Review of broad-scale drought monitoring of forests: Toward an integrated data mining approach. For. Ecol. Manag. 2016, 380, 346–358. [Google Scholar] [CrossRef]
- Hargrove, W.W.; Spruce, J.P.; Gasser, G.E.; Hoffman, F.M. Toward a national early warning system for forest disturbances using remotely sensed canopy phenology. Photogramm. Eng. Remote Sens. 2009, 75, 1150–1156. [Google Scholar]
- Noormets, A.; Chen, J.; Gu, L.; Desai, A. The phenology of gross ecosystem productivity and ecosystem respiration in temperate hardwood and conifer chronosequences. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 59–85. [Google Scholar]
- Tang, X.G.; Wang, X.; Wang, Z.M.; Liu, D.W.; Jia, M.M.; Dong, Z.Y.; Xie, J.; Ding, Z.; Wang, H.R.; Liu, X.P. Influence of vegetation phenology on modelling carbon fluxes in temperate deciduous forest by exclusive use of MODIS time-series data. Int. J. Remote Sens. 2013, 34, 8373–8392. [Google Scholar] [CrossRef]
- Estiarte, M.; Penuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: Effects on nutrient proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.; Song, C.H.; Bolstad, P.V.; Band, L.E. Downscaling real-time vegetation dynamics by fusing multi-temporal MODIS and Landsat NDVI in topographically complex terrain. Remote Sens. Environ. 2011, 115, 2499–2512. [Google Scholar] [CrossRef]
- Abarca, M.; Lill, J.T. Warming affects hatching time and early season survival of eastern tent caterpillars. Oecologia 2015, 179, 901–912. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, A.; Bajocco, S.; Ricotta, C. Phenological variability drives the distribution of wildfires in Sardinia. Landsc. Ecol. 2012, 27, 1535–1545. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Kapari, L.; Hagen, S.B.; Schott, T.; Vindstad, O.P.L.; Nilssen, A.C.; Ims, R.A. Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-arctic birch. Glob. Chang. Biol. 2011, 17, 2071–2083. [Google Scholar] [CrossRef]
- Westerling, A.L. Increasing western US forest wildfire activity: Sensitivity to changes in the timing of spring. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Reed, B.C. Surface phenology and satellite sensor-derived onset of greenness: An initial comparison. Int. J. Remote Sens. 1999, 20, 3451–3457. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Muller, A.; Kuchler, W. Possible impacts of climate change on natural vegetation in Saxony (Germany). Int. J. Biometeorol. 2005, 50, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, F.M.; Rotzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Polgar, C.A.; Primack, R.B. Leaf-out phenology of temperate woody plants: From trees to ecosystems. New Phytol. 2011, 191, 926–941. [Google Scholar] [CrossRef] [PubMed]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Reed, B.; Schwartz, M.D.; Xiao, X. Remote sensing phenology: Status and the way forward. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 231–246. [Google Scholar]
- Fisher, J.I.; Mustard, J.F. Cross-scalar satellite phenology from ground, Landsat, and MODIS data. Remote Sens. Environ. 2007, 109, 261–273. [Google Scholar] [CrossRef]
- Gu, L.; Hanson, P.J.; Mac Post, W.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 eastern us spring freezes: Increased cold damage in a warming world? Bioscience 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Fridley, J.D. Downscaling climate over complex terrain: High finescale (<1000 m) spatial variation of near-ground temperatures in a montane forested landscape (Great Smoky Mountains). J. Appl. Meteorol. Climatol. 2009, 48, 1033–1049. [Google Scholar]
- Hwang, T.; Band, L.E.; Miniat, C.F.; Song, C.H.; Bolstad, P.V.; Vose, J.M.; Love, J.P. Divergent phenological response to hydroclimate variability in forested mountain watersheds. Glob. Chang. Biol. 2014, 20, 2580–2595. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res. 2009, 39, 1259–1269. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef] [PubMed]
- Fahey, R.T. Variation in responsiveness of woody plant leaf out phenology to anomalous spring onset. Ecosphere 2016, 7, 15–17. [Google Scholar] [CrossRef]
- Prebyl, T.J. An Analysis of the Patterns and Processes Associated with Spring Forest Phenology in a Southern Appalachian Landscape Using Remote Sensing. Master’s Thesis, University of Georgia, Athens, Greece, 2012. [Google Scholar]
- Lechowicz, M.J. Why do temperate deciduous trees leaf out at different times? Adaptation and ecology of forest communities. Am. Nat. 1984, 124, 821–842. [Google Scholar] [CrossRef]
- Morin, X.; Lechowicz, M.J.; Augspurger, C.; O’ Keefe, J.; Viner, D.; Chuine, I. Leaf phenology in 22 North American tree species during the 21st century. Glob. Chang. Biol. 2009, 15, 961–975. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Lobo, A.; Maisongrande, P. Stratified analysis of satellite imagery of sw Europe during summer 2003: The differential response of vegetation classes to increased water deficit. Hydrol. Earth Syst. Sci. 2006, 10, 151–164. [Google Scholar] [CrossRef]
- Richardson, A.D.; O’Keefe, J. Phenological differences between understory and overstory: A case study using the long-term Harvard Forest records. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 87–117. [Google Scholar]
- Ryu, Y.; Lee, G.; Jeon, S.; Song, Y.; Kimm, H. Monitoring multi-layer canopy spring phenology of temperate deciduous and evergreen forests using low-cost spectral sensors. Remote Sens. Environ. 2014, 149, 227–238. [Google Scholar] [CrossRef]
- Spruce, J.P.; Sader, S.; Ryan, R.E.; Smoot, J.; Kuper, P.; Ross, K.; Prados, D.; Russell, J.; Gasser, G.; McKellip, R.; et al. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks. Remote Sens. Environ. 2011, 115, 427–437. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D.; Casady, G.M.; Neary, D.G.; Bautista, S.; Alloza, J.A.; Carmel, Y.; Wittenberg, L.; Malkinson, D.; Orr, B.J. Monitoring post-wildfire vegetation response with remotely sensed time-series data in Spain, USA and Israel. Int. J. Wildland Fire 2010, 19, 75–93. [Google Scholar] [CrossRef]
- Garonna, I.; de Jong, R.; Schaepman, M.E. Variability and evolution of global land surface phenology over the past three decades (1982–2012). Glob. Chang. Biol. 2016, 22, 1456–1468. [Google Scholar] [CrossRef] [PubMed]
- Ault, T.R.; Schwartz, M.D.; Zurita-Milla, R.; Weltzin, J.F.; Betancourt, J.L. Trends and natural variability of spring onset in the coterminous United States as evaluated by a new gridded dataset of spring indices. J. Clim. 2015, 28, 8363–8378. [Google Scholar] [CrossRef]
- Gunderson, C.A.; Edwards, N.T.; Walker, A.V.; O’Hara, K.H.; Campion, C.M.; Hanson, P.J. Forest phenology and a warmer climate—Growing season extension in relation to climatic provenance. Glob. Chang. Biol. 2012, 18, 2008–2025. [Google Scholar] [CrossRef]
- Dragoni, D.; Rahman, A.F. Trends in fall phenology across the deciduous forests of the eastern USA. Agric. For. Meteorol. 2012, 157, 96–105. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.F.; Sanchez-Castillo, M.; Dash, J.; Atkinson, P.M.; Ojeda-Zujar, J. Modelling interannual variation in the spring and autumn land surface phenology of the European forest. Biogeosciences 2016, 13, 3305–3317. [Google Scholar] [CrossRef]
- Yue, X.; Unger, N.; Keenan, T.F.; Zhang, X.; Vogel, C.S. Probing the past 30-year phenology trend of us deciduous forests. Biogeosciences 2015, 12, 4693–4709. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.H.; Wang, M.C.; Xu, J.C.; Luedeling, E. Responses of spring phenology in temperate zone trees to climate warming: A case study of apricot flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef]
- Allona, I.; Ramos, A.; Ibanez, C.; Contreras, A.; Casado, R.; Aragoncillo, C. Molecular control of winter dormancy establishment in trees. Span. J. Agric. Res. 2008, 6, 201–210. [Google Scholar] [CrossRef]
- Tanino, K.K.; Kalcsits, L.; Silim, S.; Kendall, E.; Gray, G.R. Temperature-driven plasticity in growth cessation and dormancy development in deciduous woody plants: A working hypothesis suggesting how molecular and cellular function is affected by temperature during dormancy induction. Plant Mol. Biol. 2010, 73, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Gallinat, A.S.; Primack, R.B.; Wagner, D.L. Autumn, the neglected season in climate change research. Trends Ecol. Evolut. 2015, 30, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.Y.; Wang, X.J.; Silander, J.A. Deciduous forest responses to temperature, precipitation, and drought imply complex climate change impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef] [PubMed]
- Shanks, R.E. Climates of the Great Smoky Mountains. Ecology 1954, 35, 354–361. [Google Scholar] [CrossRef]
- Lesser, M.R.; Fridley, J.D. Global change at the landscape level: Relating regional and landscape-scale drivers of historical climate trends in the Southern Appalachians. Int. J. Climatol. 2016, 36, 1197–1209. [Google Scholar] [CrossRef]
- Pierce, D.S. The Great Smokies: From Natural Habitat to National Park; The University of Tennessee Press: Knoxville, TN, USA, 2000. [Google Scholar]
- Whittaker, R.H. Vegetation of the Great Smoky Mountains. Ecol. Monogr. 1956, 26, 1–69. [Google Scholar] [CrossRef]
- Busing, R.T.; White, P.S.; Mackenzie, M.D. Gradient analysis of old spruce-fir forest of the Great Smoky Mountains circa 1935. Can. J. Bot. Rev. Can. De Bot. 1993, 71, 951–958. [Google Scholar]
- Hayes, M.; Moody, A.; White, P.S.; Costanza, J.L. The influence of logging and topography on the distribution of spruce-fir forests near their southern limits in Great Smoky Mountains National Park, USA. Plant Ecol. 2007, 189, 59–70. [Google Scholar] [CrossRef]
- Krapfl, K.J.; Holzmueller, E.J.; Jenkins, M.A. Early impacts of hemlock woolly adelgid in Tsuga canadensis forest communities of the Southern Appalachian Mountains. J. Torr. Bot. Soc. 2011, 138, 93–106. [Google Scholar] [CrossRef]
- Vose, J.M.; Wear, D.N.; Mayfield, A.E.; Nelson, C.D. Hemlock woolly adelgid in the Southern Appalachians: Control strategies, ecological impacts, and potential management responses. For. Ecol. Manag. 2013, 291, 209–219. [Google Scholar] [CrossRef]
- Welch, R.; Madden, M.; Jordan, T. Photogrammetric and GIS techniques for the development of vegetation databases of mountainous areas: Great Smoky Mountains National Park. ISPRS J. Photogramm. Remote Sens. 2002, 57, 53–68. [Google Scholar] [CrossRef]
- Harmon, M. Fire history of the western-most portion of Great Smoky Mountains National Park. Bull. Torr. Bot. Club 1982, 109, 74–79. [Google Scholar] [CrossRef]
- Flatley, W.T.; Lafon, C.W.; Grissino-Mayer, H.D.; LaForest, L.B. Fire history, related to climate and land use in three Southern Appalachian landscapes in the eastern United States. Ecol. Appl. 2013, 23, 1250–1266. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.B.; Hepinstall-Cymerman, J.; Godfrey, C.M.; Peterson, C.J. Landscape-scale characteristics of forest tornado damage in mountainous terrain. Landsc. Ecol. 2016, 31, 2097–2114. [Google Scholar] [CrossRef]
- Spruce, J.P.; Gasser, G.E.; Hargrove, W.W. MODIS NDVI Data, Smoothed and Gap-Filled, for the Conterminous Us: 2000–2015. USDA Forest Service and NASA Stennis; ORNL DAAC: Oak Ridge, TN, USA, 2016.
- Savitzky, A.; Golay, M.J.E. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-surface remote sensing of spatial and temporal variation in canopy phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite remote sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Sonnetag, O.K.; Hofkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Fry, J.; Xian, G.; Jin, S.; Dewitz, J.; Homer, C.; Yang, L.; Barnes, C.; Herold, N.; Wickham, J. Completion of the 2006 national land cover database for the conterminous United States. Photogramm. Eng. Remote Sens. 2011, 77, 858–864. [Google Scholar]
- Fu, P.; Rich, P.M. A geometric solar radiation model with applications in agriculture and forestry. Comput. Electron. Agric. 2002, 37, 25–35. [Google Scholar] [CrossRef]
- Thornton, P.E.; Running, S.W.; White, M.A. Generating surfaces of daily meteorological variables over large regions of complex terrain. J. Hydrol. 1997, 190, 214–251. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Hwang, T.; Song, C.H.; Vose, J.M.; Band, L.E. Topography-mediated controls on local vegetation phenology estimated from MODIS vegetation index. Landsc. Ecol. 2011, 26, 541–556. [Google Scholar] [CrossRef]
- Richardson, A.D.; Bailey, A.S.; Denny, E.G.; Martin, C.W.; O’Keefe, J. Phenology of a northern hardwood forest canopy. Glob. Chang. Biol. 2006, 12, 1174–1188. [Google Scholar] [CrossRef]
- Qader, S.H.; Atkinson, P.M.; Dash, J. Spatiotemporal variation in the terrestrial vegetation phenology of Iraq and its relation with elevation. Int. J. Appl. Earth Obs. Geoinf. 2015, 41, 107–117. [Google Scholar] [CrossRef]
- Kariyeva, J.; van Leeuwen, W.J.D. Environmental drivers of NDVI-based vegetation phenology in central Asia. Remote Sens. 2011, 3, 203–246. [Google Scholar] [CrossRef]
- Jochner, S.; Ziello, C.; Bock, A.; Estrella, N.; Buters, J.; Weichenmeier, I.; Behrendt, H.; Menzel, A. Spatio-temporal investigation of flowering dates and pollen counts in the topographically complex zugspitze area on the German-Austrian border. Aerobiologia 2012, 28, 541–556. [Google Scholar] [CrossRef]
- Pellerin, M.; Delestrade, A.; Mathieu, G.; Rigault, O.; Yoccoz, N.G. Spring tree phenology in the Alps: Effects of air temperature, altitude and local topography. Eur. J. For. Res. 2012, 131, 1957–1965. [Google Scholar] [CrossRef]
- Elmore, A.J.; Guinn, S.M.; Minsley, B.J.; Richardson, A.D. Landscape controls on the timing of spring, autumn, and growing season length in Mid-Atlantic forests. Glob. Chang. Biol. 2012, 18, 656–674. [Google Scholar] [CrossRef]
- Fisher, J.I.; Richardson, A.D.; Mustard, J.F. Phenology model from surface meteorology does not capture satellite-based greenup estimations. Glob. Chang. Biol. 2007, 13, 707–721. [Google Scholar] [CrossRef]
- Tuanmu, M.N.; Vina, A.; Bearer, S.; Xu, W.H.; Ouyang, Z.Y.; Zhang, H.M.; Liu, J.G. Mapping understory vegetation using phenological characteristics derived from remotely sensed data. Remote Sens. Environ. 2010, 114, 1833–1844. [Google Scholar] [CrossRef]
- Schwartz, N.B.; Urban, D.L.; White, P.S.; Moody, A.; Klein, R.N. Vegetation dynamics vary across topographic and fire severity gradients following prescribed burning in Great Smoky Mountains National Park. For. Ecol. Manag. 2016, 365, 1–11. [Google Scholar] [CrossRef]
- Bunn, W.A.; Jenkins, M.A.; Brown, C.B.; Sanders, N.J. Change within and among forest communities: The influence of historic disturbance, environmental gradients, and community attributes. Ecography 2010, 33, 425–434. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Spring | Rank | Autumn | Rank | Season | Rank |
|---|---|---|---|---|---|---|
| 2000 | 16 April | 13 | 13 October | 1 | 198 | 6 |
| 2001 | 19 April | 16 | 13 October | 1 | 182 | 14 |
| 2002 | 16 April | 12 | 3 November | 13 | 191 | 8 |
| 2003 | 1 April | 2 | 3 November | 13 | 202 | 3 |
| 2004 | 15 April | 11 | 7 November | 16 | 190 | 10 |
| 2005 | 14 April | 9 | 30 October | 10 | 206 | 1 |
| 2006 | 5 April | 3 | 17 October | 3 | 189 | 11 |
| 2007 | 12 April | 8 | 1 November | 11 | 185 | 13 |
| 2008 | 18 April | 15 | 26 October | 8 | 199 | 5 |
| 2009 | 11 April | 6 | 23 October | 5 | 191 | 8 |
| 2010 | 29 March | 1 | 1 November | 11 | 192 | 7 |
| 2011 | 14 April | 9 | 24 October | 6 | 200 | 4 |
| 2012 | 10 April | 5 | 4 November | 15 | 203 | 2 |
| 2013 | 11 April | 6 | 24 October | 6 | 188 | 12 |
| 2014 | 16 April | 12 | 22 October | 4 | 174 | 15 |
| 2015 | 5 April | 3 | 27 October | 9 | 171 | 16 |
| Mean | 11 April | - | 26 October | - | 191 | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norman, S.P.; Hargrove, W.W.; Christie, W.M. Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA. Remote Sens. 2017, 9, 407. https://doi.org/10.3390/rs9050407
Norman SP, Hargrove WW, Christie WM. Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA. Remote Sensing. 2017; 9(5):407. https://doi.org/10.3390/rs9050407
Chicago/Turabian StyleNorman, Steven P., William W. Hargrove, and William M. Christie. 2017. "Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA" Remote Sensing 9, no. 5: 407. https://doi.org/10.3390/rs9050407
APA StyleNorman, S. P., Hargrove, W. W., & Christie, W. M. (2017). Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA. Remote Sensing, 9(5), 407. https://doi.org/10.3390/rs9050407