Deoxynivalenol Affects Cell Metabolism and Increases Protein Biosynthesis in Intestinal Porcine Epithelial Cells (IPEC-J2): DON Increases Protein Biosynthesis

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
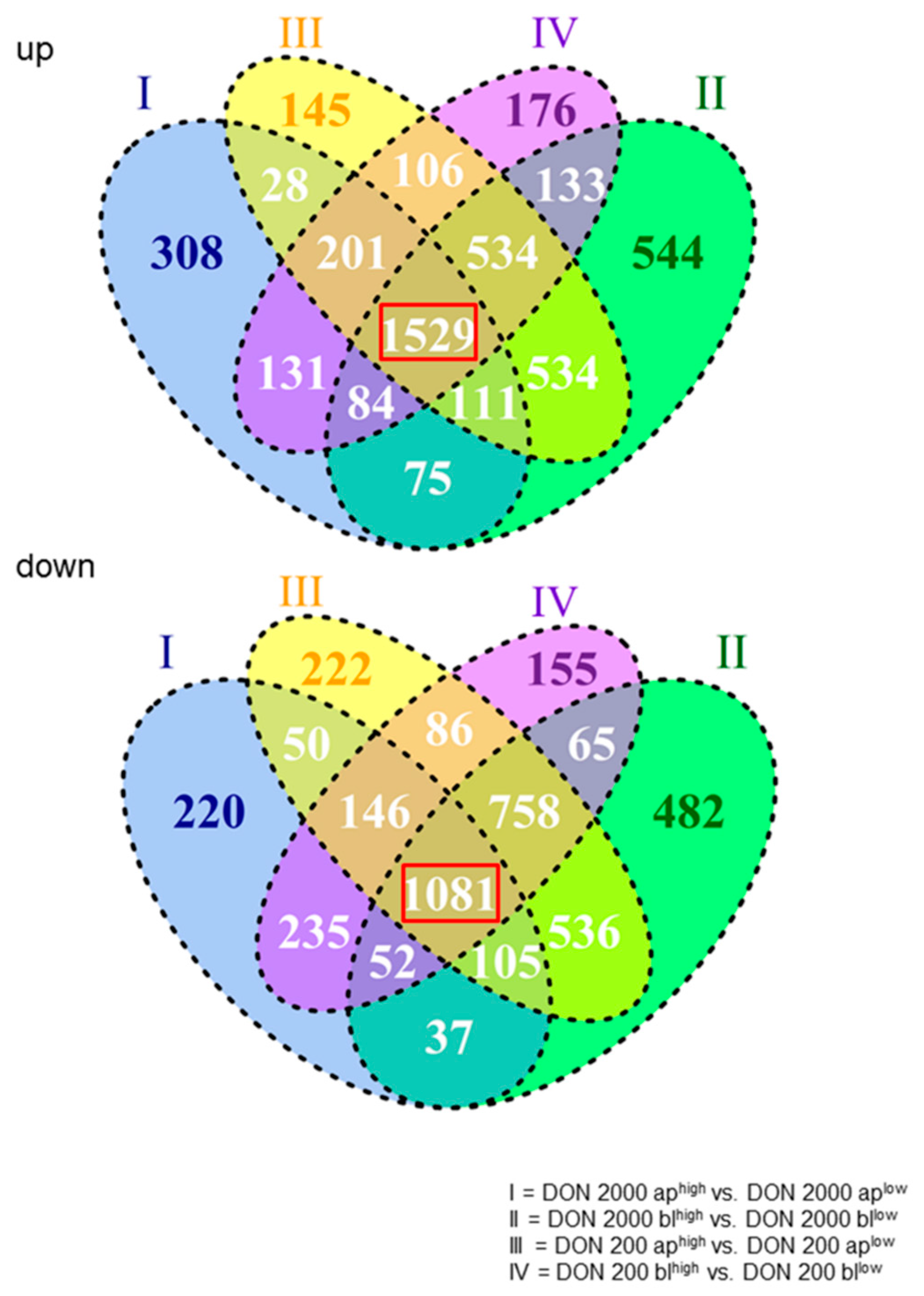
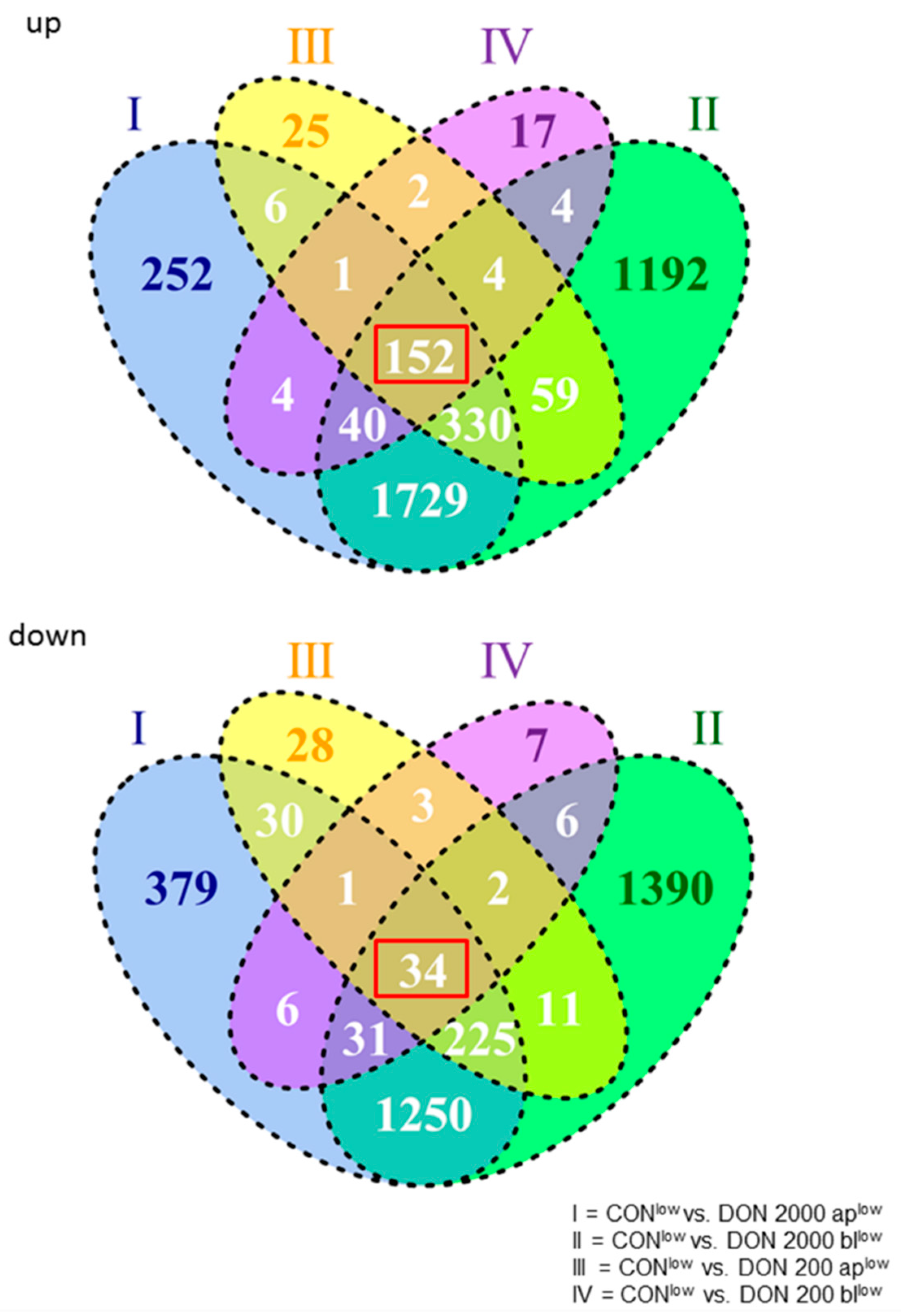
2.1. Constitutively Regulated Genes Depend on High Or Low Glucose Concentration
2.2. General Effect of DON on Gene Expression under Low Glucose Consumption
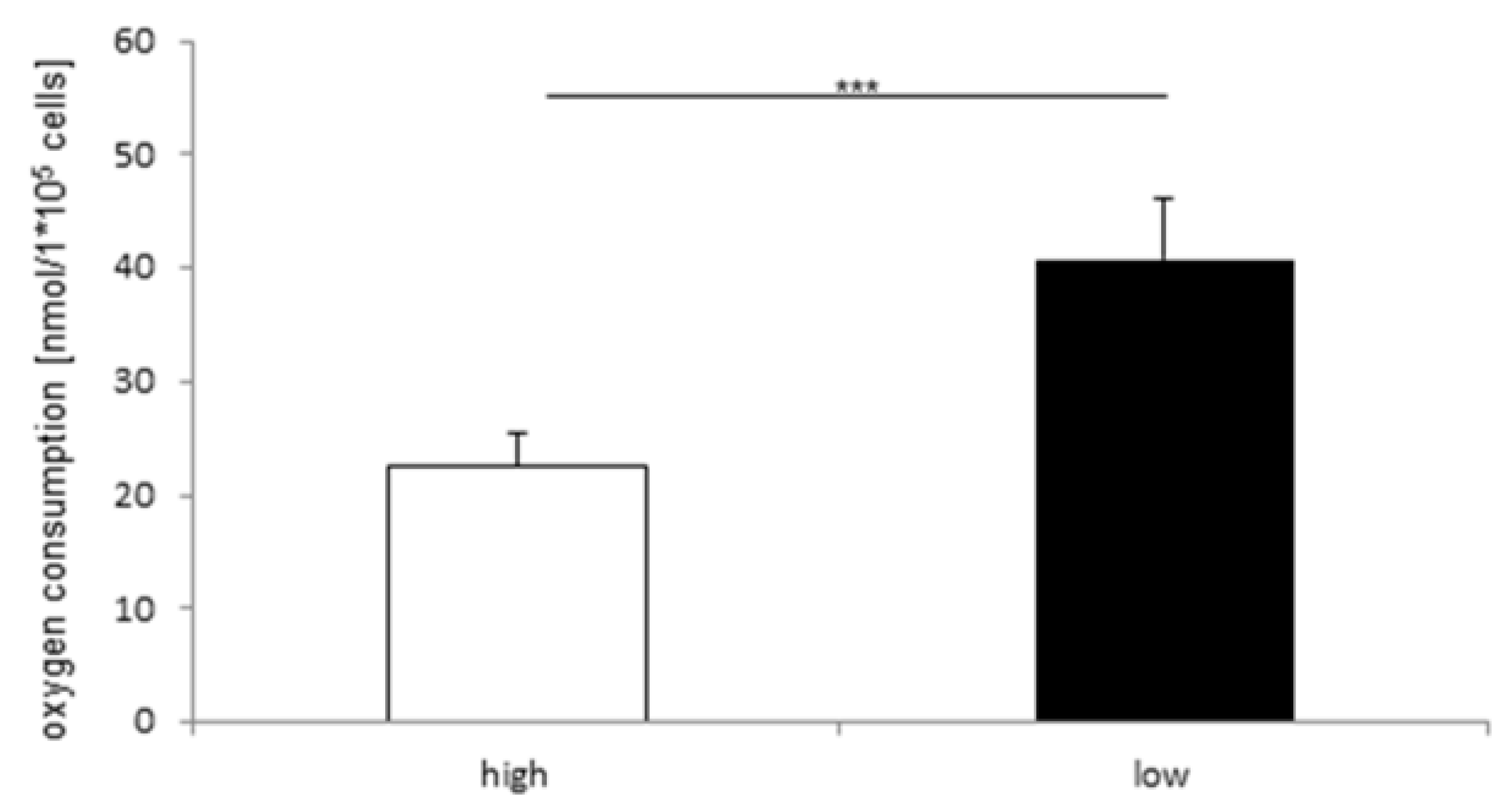
2.3. Effect of Glucose Concentration on Oxygen Consumption
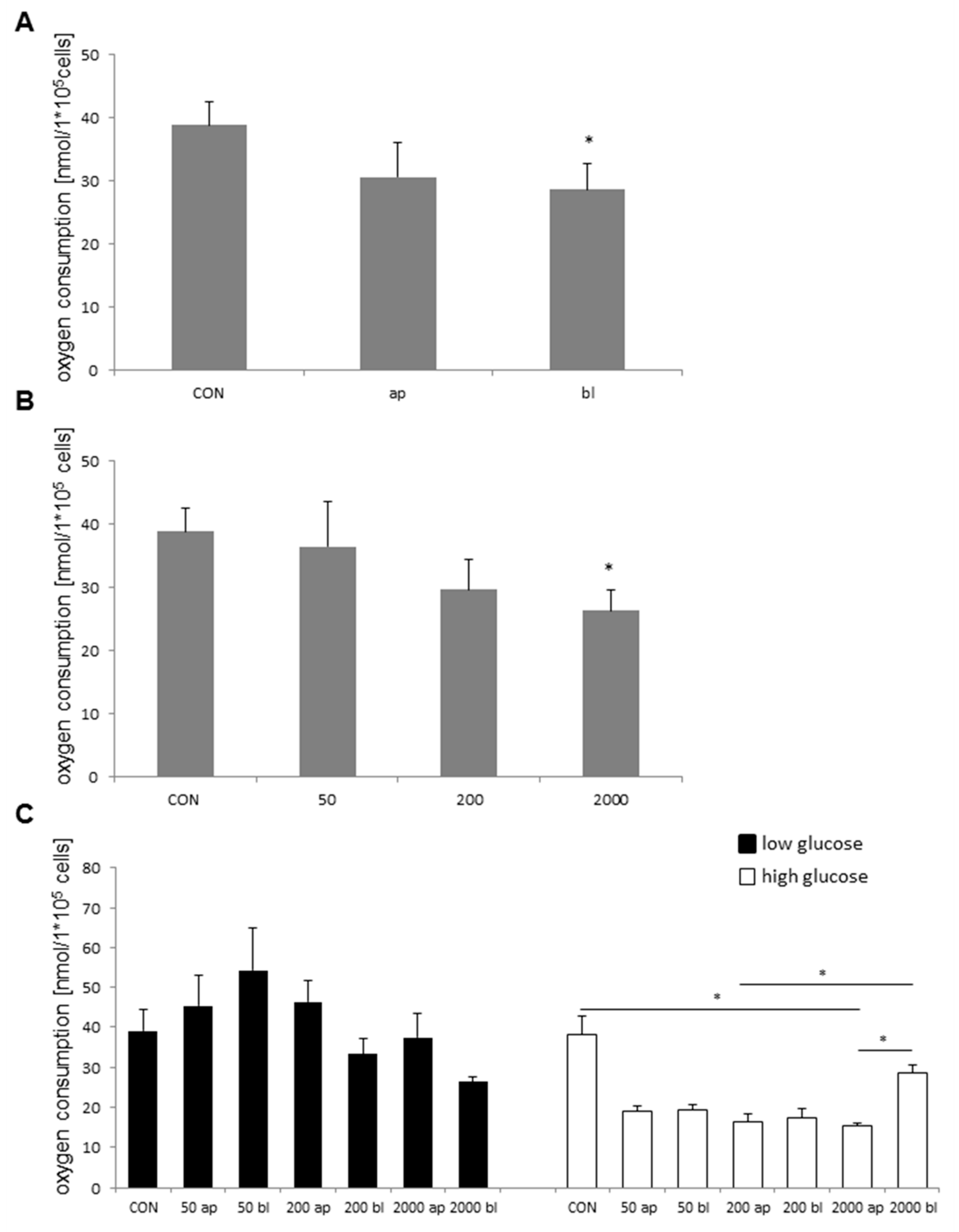
2.4. Influence of DON-Application on Oxygen Consumption
2.5. Oxygen Consumption and DON-Concentration
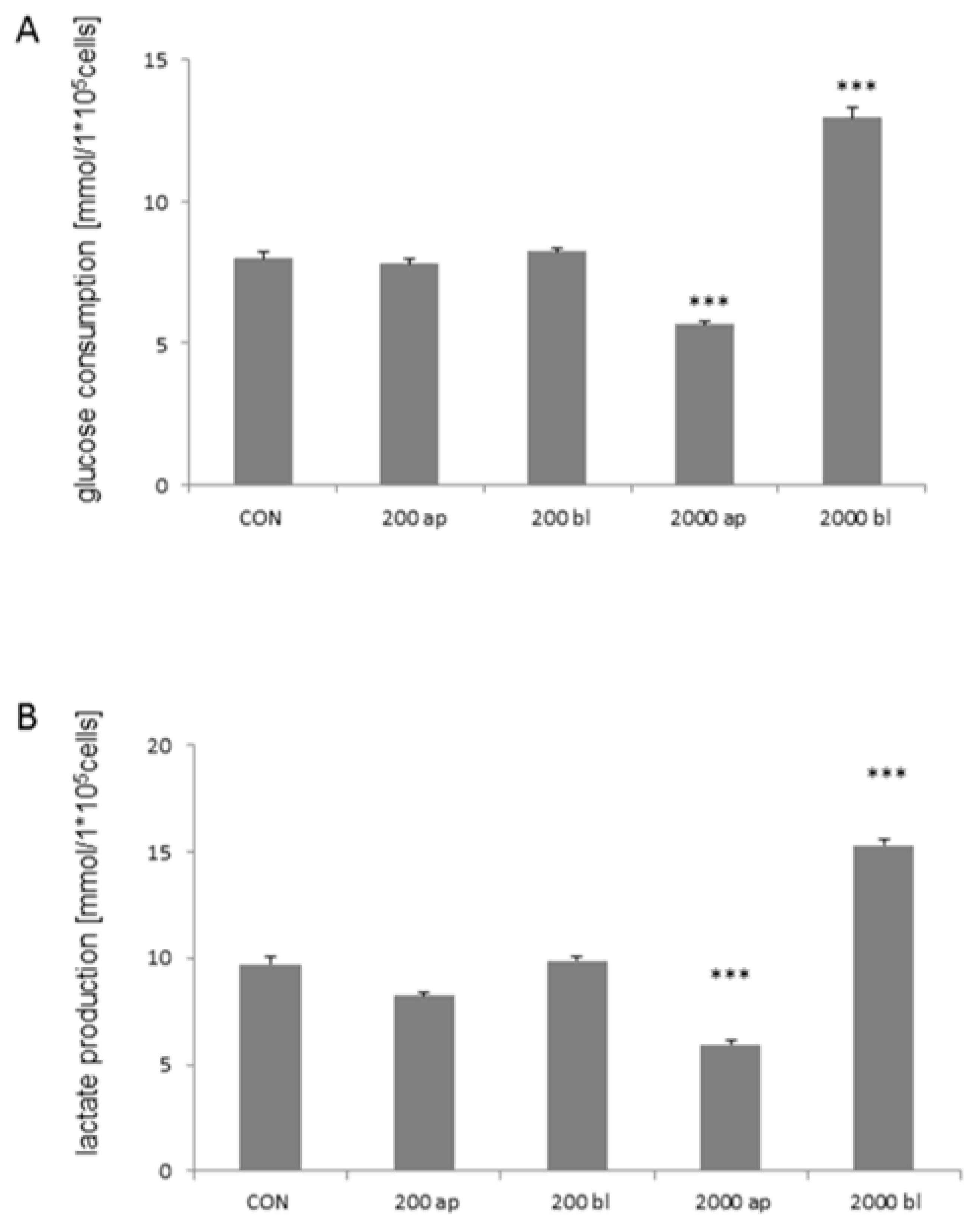
2.6. Glucose Consumption and Lactate Production under High Glucose Concentration
2.7. Metabolic Pathway in IPEC-J2
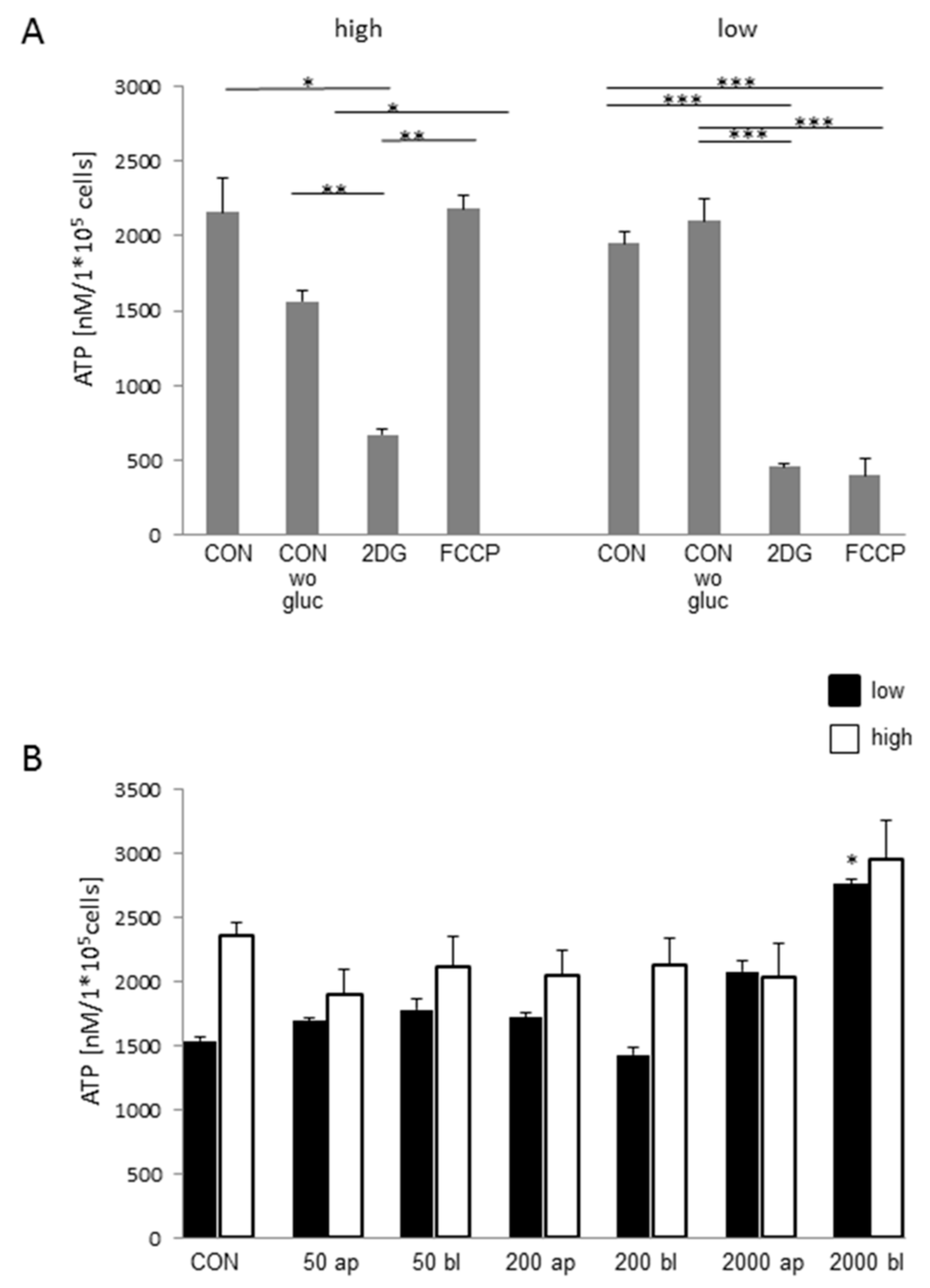
2.8. ATP-Concentration Depends on Glucose- and DON-Concentration
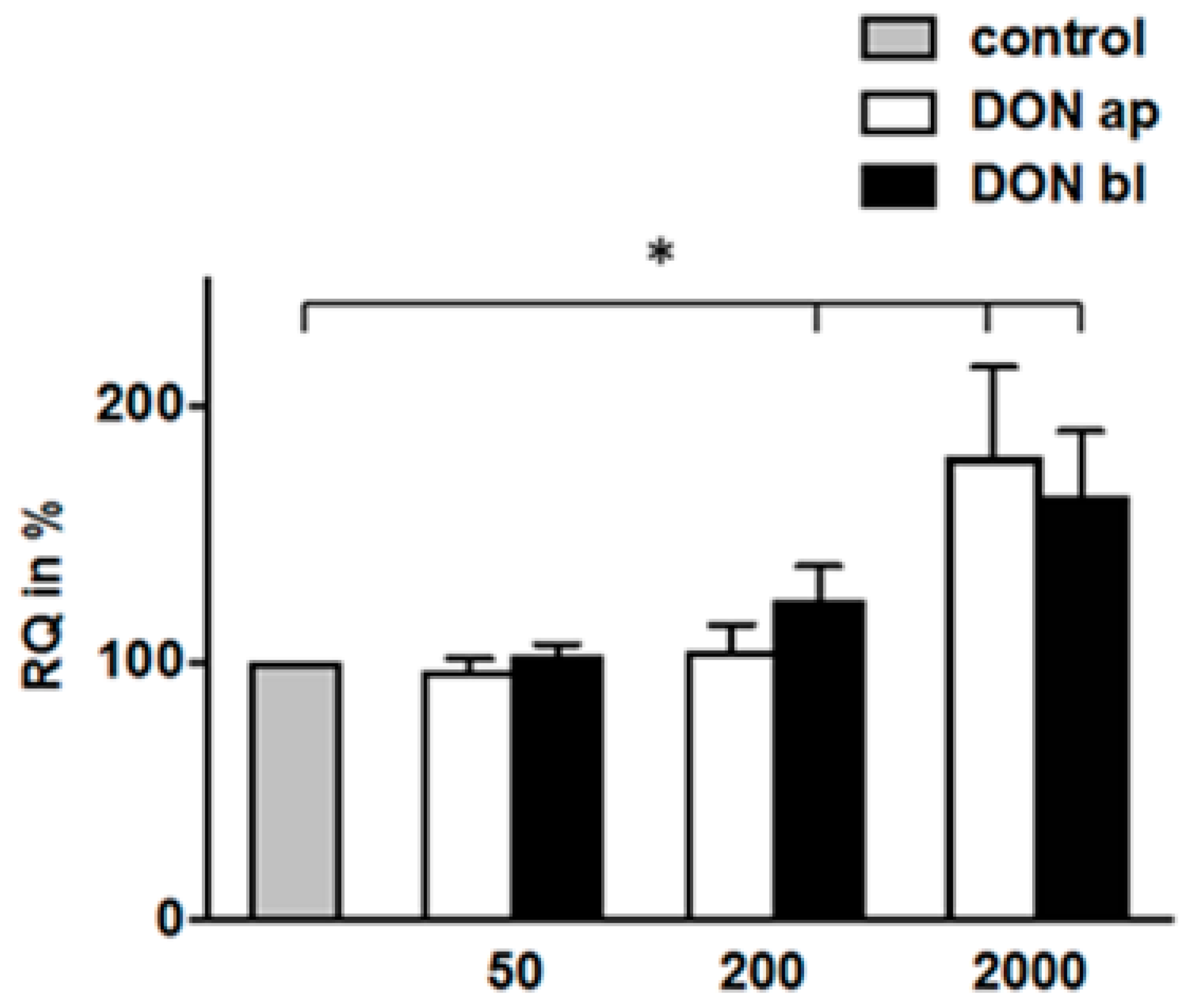
2.9. Protein Biosynthesis Rate Depend on DON-Concentration and Application under High Glucose Conditions
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. DON-Application
5.3. RNA Isolation
5.4. Microarray
5.4.1. Target Preparation and Hybridization
5.4.2. Microarray Data Processing
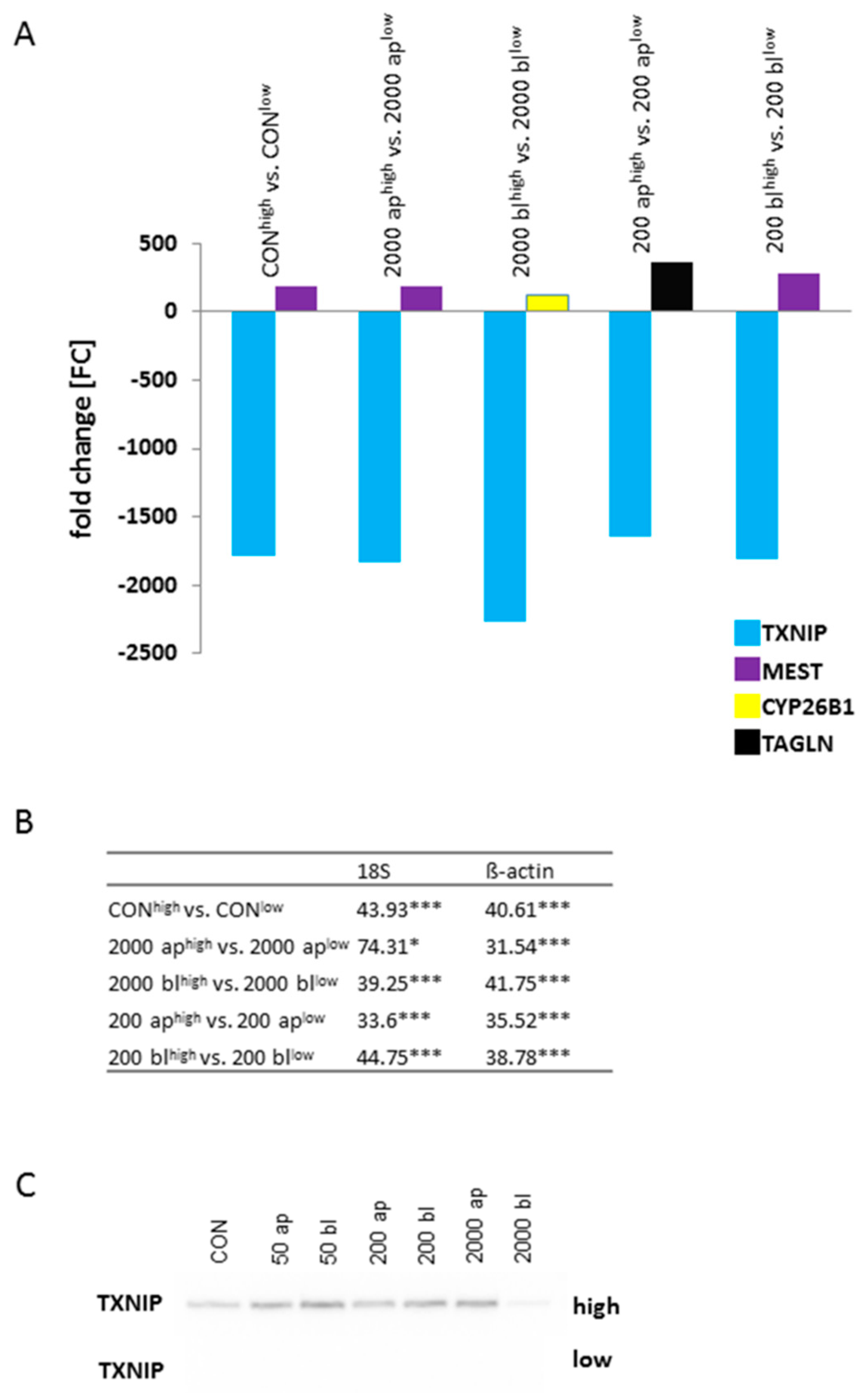
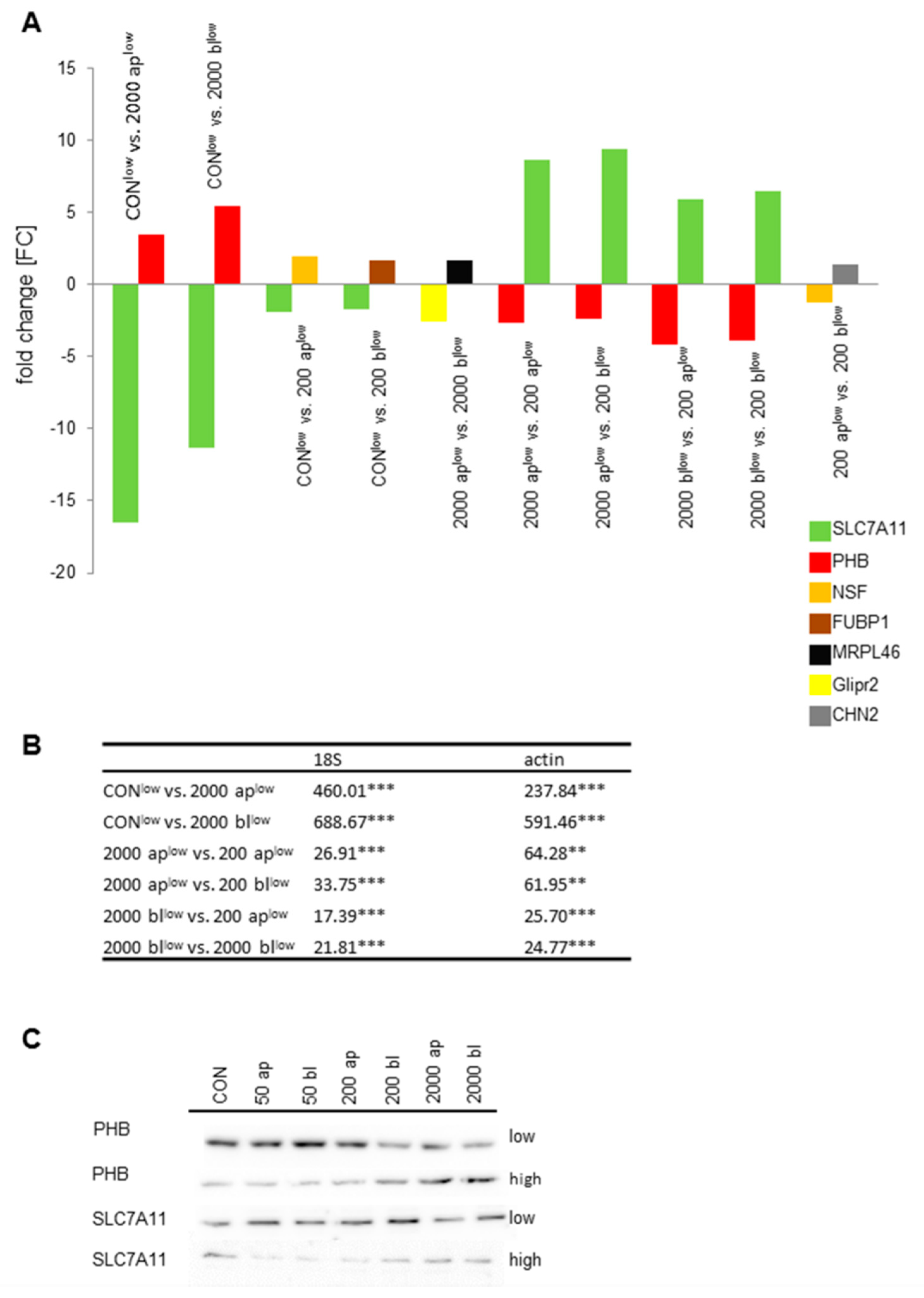
5.5. qPCR
5.6. Western Blot
5.7. Oxygen Measurement
5.8. Lactate and Glucose Measurement
5.9. ATP Measurement
5.10. Analyses of the Protein Biosynthesis
5.10.1. Metabolic Labeling and Treatment of IPEC-J2
5.10.2. Bioorthogonal Non-Canonical Amino Acid Tagging (BONCAT)
5.10.3. Western Blot Experiments and Quantitative Analysis
5.10.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AcCoA | acetyl coenzyme A |
| AHA | azidohomoalanine |
| ap | apical |
| ATP | adenosinetriphosphate |
| bl | basolateral |
| BONCAT | bioorthogonal non-canonical amino acid tagging |
| CAV2 | caveolin-2 |
| COX5B | cytochrome C oxidase subunit 5B |
| CYP26B1 | cytochrome P450 26B1 |
| 2DG | 2-deoxy-glucose |
| DON | deoxynivalenol |
| EGF | epidermal growth factor |
| EST | expressed sequence tags |
| ETC | electron transport chain |
| FADH | flavine adenine dinucleotide |
| FBS | fetal bovine serum |
| FCCP | carbonylcyanid-4-trifluormethoxyphenylhydrazon |
| GAPDH | glycerinaldehyd-3-phosphate dehydrogenase |
| GSH | glutathione |
| HEPES | 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid |
| IPEC-J2 | intestinal porcine epithelial cells |
| ITS | insulin-transferrin-selenium |
| Met | methionine |
| MEST | mesoderm-specific transcript homologues protein |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| OxPhos | oxidative phosphorylation |
| PB | phosphate buffer |
| PBS | phosphate buffered saline |
| PHB | prohibitin |
| PKR | protein kinase R |
| PVDF | polyvinylidenfluoride |
| ROS | reactive oxygen species |
| RQ | relative quantification |
| SDS | sodium dodecyl sulfate |
| SLC6A19 | system B(0) neutral amino acid transporter AT1 |
| SLC7A11 | solute carrier family 7 member 11 |
| TAGLN | transgelin |
| TBTA | tris[(1-benzyl-1H-1,2,3-triazol-4-yl)methyl]amine |
| TRA1 | tumor rejection antigen 1 (≙ GRP94, HSP90) |
| TRIS | tris(hydroxymethyl)aminomethane |
| VDUP1 | vitamin D3 upregulated protein1 |
References
- Dalli, J. The Rapid Alert System for Food and Feed; Office for Official Publications of the European Communities: Luxembourg, 2010; ISBN 978-92-79-19765-9. [Google Scholar]
- Schothorst, R.C.; Jekel, A.A. Determination of trichothecenes in beer by capillary gas chromatography with flame ionisation detection. Food Chem. 2003, 82, 475–479. [Google Scholar] [CrossRef]
- Rotter, B.; Prelusky, D.; Pestka, J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Mahfoud, R.; Garmy, N.; Fantini, J. The mycotoxin deoxynivalenol affects nutrient absorption in human intestinal epithelial cells. J. Nutr. 2002, 132, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liao, P.; He, L.; Ren, W.; Yin, J.; Duan, J.; Li, T. Growth performance serum biological profile, jejunal morphology, and the expression of nutrient transporter genes in deoxynivalenol-challenged growing pigs. BMC Vet. Res. 2015, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Kersten, S.; Valenta, H.; Breves, G. Inactivation of deoxinevalenol-contaminated cereal grains with sodium metabisulfite: A review of procedures and toxical aspects. Mycotoxin Res. 2012, 28, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Zerull, K.; Breves, G.; Schröder, B.; Goyarts, T.; Dänicke, S. The influence of the mycotoxin deoxynivalenol on the jejunal glucose transport in pigs. Mycotoxin Res. 2005, 21, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Diesing, A.K.; Nossol, C.; Dänicke, S.; Walk, N.; Post, A.; Kahlert, S.; Rothkötter, H.-J.; Kluess, J. Vulnerability of polarized Intestinal porcine epithelial cells to mycotoxin Deoxynivalenol depends on the route of Application. PLoS ONE 2011, 6, e17472. [Google Scholar] [CrossRef] [PubMed]
- Diesing, A.K.; Nossol, C.; Ponsuksili, S.; Wimmers, K.; Kluess, J.; Walk, N.; Post, A.; Rothkötter, H.-J.; Kahlert, S. Gene regulation of intestinal porcine epithelial cells IPEC-J2 is dependent on the site of Deoxynivalenol toxicology action. PLoS ONE 2012, 7, e34136. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol-induced proinflammatory gene expression: Mechanisms and pathological sequelae. Toxins 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Parys, M.; Garsou, S.; Pussemier, L.; Schneider, Y.J.; Larondelle, Y. Deoxynivalenol transport across human intestinal caco-2 cells and its effects on cellular metabolism at realistic intestinal concentrations. Toxicol. Lett. 2006, 164, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, B.; McLaughlin, C.S. Biochemical mechanism of action of trichothecene mycotoxins. In Trichothecene Mycotoxicosis: Pathophysiological Effects; Beasley, V.R., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 27–35. [Google Scholar]
- Dänicke, S.; Goyarts, T.; Döll, S.; Grove, N.; Spolders, M.; Flachowski, G. Effects of the Fusaium toxin deoxynivalenol on tissue protein synthesis in pig. Toxicol. Lett. 2006, 165, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.R.; Lau, A.L.; Pestka, J.J. Role of double-stranded RNA-activated protein kinase R (PKR) in deoxynivalenol-induced ribotoxic stress response. Toxicol. Sci. 2003, 74, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, R.; Devaraj, S.N.; Padma, V.V. Lutein protects HT-29 cells against deoxynivalenol-induced oxidative stress and apoptosis: Prevention of NF-κB nuclear localization and down regulation of NF-κB and cyclo-oxygenase—2 expression. Free Radic. Biol. Med. 2010, 49, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.; Rao, P.V. Brain oxidative stress after dermal and subcutaneous exposure of T-2 toxin in mice. Food. Chem. Toxicol. 2010, 48, 3436–3442. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotosky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nossol, C.; Barta-Böszörmènyi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.K.; Rothkötter, H.-J. Comparing two intestinal porcine epithelial cell lines (IPECs): Morphological differentiation, function and metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef] [PubMed]
- Nossol, C.; Diesing, A.K.; Kahlert, S.; Kersten, S.; Kluess, J.; Ponsuksili, S.; Hartig, R.; Wimmers, K.; Dänicke, S.; Rothköther, H.-J. Deoxynivalenol affects the composition of the basement membrane proteins and influences en route the migration of CD16+ cells into the intestinal epithelium. Mycotoxin Res. 2013, 29, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Klasvogt, S.; Zuschratter, W.; Schmidt, A.; Kröber, A.; Vorwerk, S.; Wolter, R.; Isermann, B.; Wimmers, K.; Rothkötter, H.-J.; Nossol, C. Air-liquid interface enhances oxidative phosphorylation in intestinal epithelial cell line IPEC-J2. Cell Death Discov. 2017, 3, 17001. [Google Scholar] [CrossRef] [PubMed]
- Penny, H.L.; Siew, J.L.; Adriani, G.; Yeao, W.H.; Ee, P.S.; Luis, B.S.; Lee, B.; Lee, T.; Mak, S.Y.; Ho, Y.S.; et al. Warburg metabolism in tumor-conditioned macrophages promotes metastasis in human pancreatic ductal adenocarcinoma. Oncoimmunology 2016, 5, e1191731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, R.; Wani, N.A.; Ghoshal, K.; Jacob, S.T.; Motiwala, T. Sorafenib and 2-Deoxyglucose synergistically inhibit proliferation of both Sorafenib sensitive and resistant HCC cells by inhibiting ATP production. Gene Expr. 2017, 17, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Matsui, M.; Iwata, S.; Hirota, K.; Masutani, H.; Nakamura, H.; Takagi, Y.; Sono, H.; Gon, Y.; Yodoi, J. Identification of thioredoxin-binding protein-1/vitamin D3) up-regulated protein 1 as a negative regulator of thioredoxin function and expression. J. Biol. Chem. 1999, 274, 21645–21650. [Google Scholar] [CrossRef] [PubMed]
- Yodoi, J.; Nakamura, H.; Masutani, H. Redox regulation of stress signals: Possible roles of dendritic stellate TRX producer cells (DST cell types). Biol. Chem. 2002, 383, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Hui, T.Y.; Sheth, S.S.; Diffley, J.M.; Potter, D.W.; Lusis, A.J.; Attie, A.D.; Davis, R.A. Mice lacking thioredoxin-interacting protein provide evidence linking cellular redox state to appropriate response to nutritional signals. J. Biol. Chem. 2004, 279, 24387–24393. [Google Scholar] [CrossRef] [PubMed]
- Chal-Molstad, H.; Saxena, G.; Chen, J.; Shalev, A. Glucose-stimulated expression of Txnip is mediated by carbohydrate response element-binding protein, p300, and histone H4 acetylation in pancreatic beta cells. J. Biol. Chem. 2009, 284, 16898–16905. [Google Scholar] [CrossRef] [PubMed]
- Pradelli, L.A.; Villa, E.; Zunino, E.; Marchetti, B.; Ricci, J.E. Glucose metabolism is inhibited by caspases upon the induction of apoptosis. Cell Death Dis. 2014, 5, e1460. [Google Scholar] [CrossRef] [PubMed]
- Assinder, S.J.; Stanton, J.A.; Prasad, P.D. Transgelin: An actin-binding protein and tumour suppressor. Int. J. Biochem. Cell Biol. 2009, 41, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Lees-Miller, J.P.; Heeley, D.H.; Smillie, L.B. An abundant and novel protein of 22 kDa (SM22) is widely distributed in smooth muscles: Purification from bovine aorta. Biochem. J. 1987, 244, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chang, Y.J.; Miyamoto, H.; Ni, J.; Niu, Y.; Chen, Z.; Chen, Y.L.; Yao, J.L.; di Sant Agnese, P.A.; Chang, C. Transgelin functions as a suppressor via inhibition of ARA54-enhanced androgen receptor transactivation and prostate cancer cell growth. Mol. Endocrinol. 2007, 21, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Huang, Q.; Chen, W.; Wang, L.; Lin, W.; Lin, J.; Lin, X. Identification of transgelin as a potential novel biomarker for gastric adenocarcinoma based on proteomics technology. J. Cancer Res. Clin. Oncol. 2008, 134, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Little, E.; Ramakrishnan, M.; Roy, B.; Gazit, G.; Lee, A.S. The glucose-regulated proteins (GRP78 and GRP94): Functions, gene regulation, and application. Crit. Rev. Eukaryot. Gene Expr. 1994, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Vahjen, W.; Aschenbach, J.R.; Zentek, J. A diet naturally contaminated with the Fusarium mycotoxin deoxynivalenol (DON) down regulates gene expression of glucose transporters in the intestine of broiler chickens. Livest. Sci. 2011, 140, 72–79. [Google Scholar] [CrossRef]
- Greiner, E.F.; Guppy, M.; Brand, K. Glucose is essential for proliferation and the glycolytic enzyme induction that provokes a transition to glycolytic energy production. J. Biol. Chem. 1994, 269, 31484–31490. [Google Scholar] [PubMed]
- Sato, H.; Tamba, M.; Kuriyama-Matsumura, K.; Okuno, S.; Bannai, S. Molecular cloning and expression of human xCT, the light chain of amino acid transport system xc. Antioxid. Redox Signal. 2000, 2, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Gatti, L.; Zunino, F. Overview of tumor cell chemoresistance mechanisms. Methods Mol. Med. 2005, 111, 127–148. [Google Scholar] [PubMed]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.E.; Park, S.Y.; Wong, S.S.; Pestka, J.J. Modulation of nitric oxide, hydrogen peroxide and cytokine production in a clonal macrophage model by the trichothecene vomitoxin (deoxynivalenol). Toxicology 1998, 125, 203–214. [Google Scholar] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Zielke, H.R.; Zielke, C.L.; Ozand, P.T. Glutamine: A major energy source for cultured mammalien cells. Fed. Proc. 1984, 43, 121–125. [Google Scholar] [PubMed]
- Curi, R.; Lagranha, C.J.; Doi, S.Q.; Sellitti, D.F.; Procopio, J.; Pithon-Curi, T.C.; Corless, M.; Newsholme, P. Molecular mechanisms of glutamin action. J. Cell Physiol. 2005, 204, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Bourges, I.; Ramus, C.; Mousson de Camaret, B.; Beugnot, R.; Remacle, C.; Cardol, P.; Hofhaus, G.; Issartel, J.P. Structural organization of mitochondrial human complex I: Role of the ND4 and ND5 mitochondria-encoded subunits and interaction with prohibitin. Biochem. J. 2004, 383, 491–499. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond aerobic glycolysis: Transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. PNAS 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Bannert, E.; Tesch, T.; Kluess, J.; Frahm, J.; Kersten, S.; Kahlert, S.; Renner, L.; Rothkötter, H.-J.; Dänicke, S. Metabolic and Hematological consequences of dietary deoxynivalenol interacting with systemic Escherichia coli lipopolysaccharide. Toxins 2015, 7, 4773–4796. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhang, X.; Liu, L.; Chen, J.; Nie, Z.; Sheng, Q.; Zhang, W.; Jiang, C.; Yu, W.; Wang, D.; et al. Characterization of a gene encoding prohibitin in silkworm, Bombyx Mori. Gene 2012, 502, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Liemburg-Apers, D.C.; Wagenaars, J.A.L.; Smeitink, J.A.M.; Willems, P.H.G.M.; Koopman, W.J.H. Acute stimulation of glucose influx upon mitoenergetic dysfunction requires LKB1, AMPK, Sirt2 and mTOR–RAPTOR. J. Cell Sci. 2011, 129, 4411–4423. [Google Scholar] [CrossRef] [PubMed]
- Nijtamns, L.G.; de Jong, L.; Artal Sanz, M.; Coates, P.J.; Berden, J.A.; Back, J.W.; Muijsers, A.O.; van der Spek, H.; Grivell, L.A. Prohibitins act as a membrane-bound chaperone for the stabilization of mitochondrial proteins. EMBO J. 2000, 19, 2444–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathiria, A.S.; Butcher, L.D.; Feagins, L.A.; Souza, R.F.; Boland, C.R.; Theiss, A.L. Prohihitin 1 modulates mitochondrial stress-related autophagy in human colonic epithelial cells. PLoS ONE 2012, 7, e31231. [Google Scholar] [CrossRef] [PubMed]
- Merkwirth, C.; Langer, T. Prohibitin function within mitochondria: Essential roles for cell proliferation and cristae morphogenesis. BBA-Mol. Cell. Res. 2009, 1793, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Dong, P.; Zhang, Z.; Li, C.; Li, Y.; Li, X.; Wu, Z.; Guo, S.; Mai, S.; Xia, D.; et al. Akt phosphorylates Prohibitin 1 to mediate its mitochondrial localization and promote proliferation of bladder cancer cells. Cell Death Dis. 2015, 6, e1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strub, G.M.; Paillard, M.; Liang, J.; Gomez, L.; Allegood, J.C.; Hait, N.C.; Maceyka, M.; Price, M.M.; Chen, Q.; Simpson, D.C.; et al. Sphingosine-1-phosphate produced by sphingosine kinase 2 in mitochondria interacts with prohibitin 2 to regulate complex IV assembly and respiration. FASEB J. 2011, 25, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, M.; Shepherd, B.R.; Suarez, Y.; Fernandez-Hernando, C.; Yu, J.; Pan, Y.; Acevedo, M.L.; Shadel, G.S.; Sessa, W.C. Prohibitin-1 maintains the angiogenic capacity of endothelial cells by regulating mitochondrial function and senescence. J. Cell Biol. 2008, 180, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steglich, G.; Neupert, W.; Langer, T. Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol. Cell. Biol. 1999, 19, 3435–3442. [Google Scholar] [CrossRef] [PubMed]
- Steube, K.G.; Koelz, A.L.; Uphoff, C.C.; Drexler, H.G.; Kluess, J.; Steinberg, P. The necessitiy of identity assessment of animal intestinal cell lines: A case report. Cytotechnology 2012, 64, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Berschneider, H.M. Development of normal cultured small intestinal epithelial cell lines which transport Na and Cl (Abstract). Gasteroenterology 1989, 96, A41. [Google Scholar]
- Edgar, R.; Domrachev, M.; Lash, A. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffmann, A.; Gentleman, R.; Huber, W. arrayQualityMetrics—A bioconductor package for quality assessment of microarray data. Bioinformatics 2009, 25, 415–416. [Google Scholar] [CrossRef] [PubMed]
- Bourgon, R.; Gentleman, R.; Huber, W. Independent filtering increases detection power for high-throughput experiments. Proc. Natl. Acad. Sci. USA 2010, 107, 9546–9551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acid Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Stellmacher, A.; Freitag, C.E.; Landgraf, P.; Dieterich, D.C. Monitoring Astrocytic Proteome Dynamics by Cell Type-Specific Protein Labeling. PLoS ONE 2015, 21, e0145451. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, D.C.; Lee, J.J.; Link, A.J.; Graumann, J.; Tirell, D.A.; Schuman, E.M. Labeling, detection and identification of newly synthesized proteomes with bioorthogonal noncanonical amino acid tagging. Nat. Protoc. 2007, 2, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, P.; Antileo, E.R.; Schumann, E.M.; Dieterich, D.C. BONCAT: Metabolic labeling, click chemistry, and affinity purification of newly synthesized proteoms. Meth. Mol. Biol. 2014, 1266, 199–215. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Pathways |
|---|---|
| 1 | Metabolic pathways |
| 2 | Pathways in cancer |
| 3 | Endocytosis |
| 4 | PI3K-Akt signaling pathway |
| 5 | Biosynthesis of antibiotics |
| TXNIP | ||||
| 18S | Actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 109.68 | 112.25 | 138.83 | 128.94 * |
| 50 bl | 100.46 | 115.4 | 123.68 | 130.74 * |
| 200 ap | 86,65 | 113.29 | 113.81 | 130.13 * |
| 200 bl | 125.99 | 123.68 | 127.16 | 133.18 * |
| 2000 ap | 216.1 *** | 127.75 | 110.33 * | 142.08 * |
| 2000 bl | 111.03 | 124.26 | 96.82 | 94.17 *** |
| COX5B | ||||
| 18S | Actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 102.34 | 89.09 | 129.53 | 102.45 |
| 50 bl | 105.6 | 92.02 | 127.62 | 104.36 |
| 200 ap | 85.86 | 96.37 | 112.77 | 110.82 |
| 200 bl | 109.18 | 89.09 | 110.19 | 96.03 |
| 2000 ap | 242 *** | 144.47 | 125.12 *** | 158.91 *** |
| 2000 bl | 172.4 | 140.03 *** | 149.48 | 105.92 |
| GLUT1 | ||||
| 18S | Actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 127.16 | 91.59 | 160.96 ** | 105.21 |
| 50 bl | 126.58 | 97.72 | 155.83 ** | 110.7 |
| 200 ap | 119.75 | 138.84 | 157.28 ** | 159.48 * |
| 200 bl | 150.18 | 115.4 | 151.57 * | 124.26 |
| 2000 ap | 460.01 *** | 891.61 *** | 237.84 *** | 985.62 *** |
| 2000 bl | 688.67 *** | 483.44 *** | 591.46 *** | 366.38 *** |
| MCT1 | ||||
| 18S | Actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 115.94 | 85.46 | 146.75 *** | 94.95 |
| 50 bl | 119.27 | 88.27 | 145.4 *** | 96.72 |
| 200 ap | 93.74 | 92.99 | 123.11 | 104.56 |
| 200 bl | 99.08 | 88.27 | 100 | 91.93 |
| 2000 ap | 194.08 * | 112.8 | 99.66 | 125.58 |
| 2000 bl | 154.3 | 100 | 133.79 ** | 73.3 |
| SGLT1 | ||||
| 18S | Actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 118.65 | 107.14 | 150.18 *** | 123.32 |
| 50 bl | 123.11 | 114.87 | 151.57 *** | 130.13 |
| 200 ap | 88.68 | 80.8 | 116.47 | 92.08 |
| 200 bl | 118.41 | 108.21 | 117.98 | 118.53 |
| 2000 ap | 176.23 ** | 78.43 | 85.75 | 90.27 |
| 2000 bl | 81.78 | 62.2 | 67.43 *** | 46.12 *** |
| PHB | ||||
| 18S | β-actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 127.16 | 91.59 | 160.96 | 105.21 |
| 50 bl | 126.58 | 97.72 | 155.83 | 110.7 |
| 200 ap | 119.75 | 138.84 | 157.28 | 159.48 |
| 200 bl | 150.18 | 115.4 | 151.57 | 124.26 |
| 2000 ap | 460.01 *** | 891.61 *** | 237.84 *** | 985.62 *** |
| 2000 bl | 688.67 *** | 483.439 *** | 591.46 *** | 366.38 *** |
| SLC7A11 | ||||
| 18S | β-actin | |||
| Low | High | Low | High | |
| CON | 100 | 100 | 100 | 100 |
| 50 ap | 127.75 | 110.19 | 161.7 | 126.58 |
| 50 bl | 126.99 | 136.29 | 153.48 | 154.4 *** |
| 200 ap | 92.87 | 84.67 | 121.98 | 97.27 |
| 200 bl | 97.72 | 108.17 | 98.62 | 116.47 |
| 2000 ap | 30.57 *** | 62.31 | 15.8 *** | 68.87 ** |
| 2000 bl | 31.59 *** | 92.02 | 28.32 *** | 69.74 |
| Number | Pathways |
|---|---|
| 1 | Spliceosomes |
| 2 | RNA transport |
| 3 | Epstein–Barr virus infection |
| 4 | Ribosome biogenesis in eukaryotes |
| 5 | mRNA surveillance pathway |
| Gene | Left (5′-3′) | Right (5′-3′) | Product Size | Temperature [°C] |
|---|---|---|---|---|
| 18S | GCAATTATTCCCCATGAACG | GGCCTCACTAAACCATCCAA | 123 | 56.5 |
| beta-actin | GATGAGATTGGCATGGCTTT | CACCTTCACCGTTCCAGTTT | 122 | 58.3 |
| TXNIP | AGCAGCCAAGAGAACAGAGA | TCCACGGACACAATACCCA | 118 | 57.2 |
| PHB | TGA AAA CTC TGC CCC TGT GA | TCT GCA GGA CTC ACA TCT CG | 119 | 57.3 |
| SLC7A11 | TAA ATT TGG GTG CAA TGT GAT GT | TTG AAG CAA CTA GAA GCA TGA CA | 99 | 54.9 |
| MCT1 | TCCATCATGTTGGCTGTCAT | GAAGGAAGCTGCAATCAAGC | 129 | 58.5 |
| COX5B | GGAGAGGGAGGTCATGATGG | CCACTATCCGCTTGTTGGTG | 128 | 59.4 |
| SGLT-1 | AAGCTGGTCATGGAGCTGAT | AGACGTCCATGGTGAAGAGG | 127 | 61.5 |
| GLUT1 | GAGCCCTGCCTAGACACTTG | CCACCTCTTGGGGTAGAAGA | 112 | 62 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nossol, C.; Landgraf, P.; Kahlert, S.; Oster, M.; Isermann, B.; Dieterich, D.C.; Wimmers, K.; Dänicke, S.; Rothkötter, H.-J. Deoxynivalenol Affects Cell Metabolism and Increases Protein Biosynthesis in Intestinal Porcine Epithelial Cells (IPEC-J2): DON Increases Protein Biosynthesis. Toxins 2018, 10, 464. https://doi.org/10.3390/toxins10110464
Nossol C, Landgraf P, Kahlert S, Oster M, Isermann B, Dieterich DC, Wimmers K, Dänicke S, Rothkötter H-J. Deoxynivalenol Affects Cell Metabolism and Increases Protein Biosynthesis in Intestinal Porcine Epithelial Cells (IPEC-J2): DON Increases Protein Biosynthesis. Toxins. 2018; 10(11):464. https://doi.org/10.3390/toxins10110464
Chicago/Turabian StyleNossol, Constanze, Peter Landgraf, Stefan Kahlert, Michael Oster, Berend Isermann, Daniela C. Dieterich, Klaus Wimmers, Sven Dänicke, and Hermann-Josef Rothkötter. 2018. "Deoxynivalenol Affects Cell Metabolism and Increases Protein Biosynthesis in Intestinal Porcine Epithelial Cells (IPEC-J2): DON Increases Protein Biosynthesis" Toxins 10, no. 11: 464. https://doi.org/10.3390/toxins10110464
APA StyleNossol, C., Landgraf, P., Kahlert, S., Oster, M., Isermann, B., Dieterich, D. C., Wimmers, K., Dänicke, S., & Rothkötter, H. -J. (2018). Deoxynivalenol Affects Cell Metabolism and Increases Protein Biosynthesis in Intestinal Porcine Epithelial Cells (IPEC-J2): DON Increases Protein Biosynthesis. Toxins, 10(11), 464. https://doi.org/10.3390/toxins10110464