A Time Series of Water Column Distributions and Sinking Particle Flux of Pseudo-Nitzschia and Domoic Acid in the Santa Barbara Basin, California


Abstract
:1. Introduction
2. Results
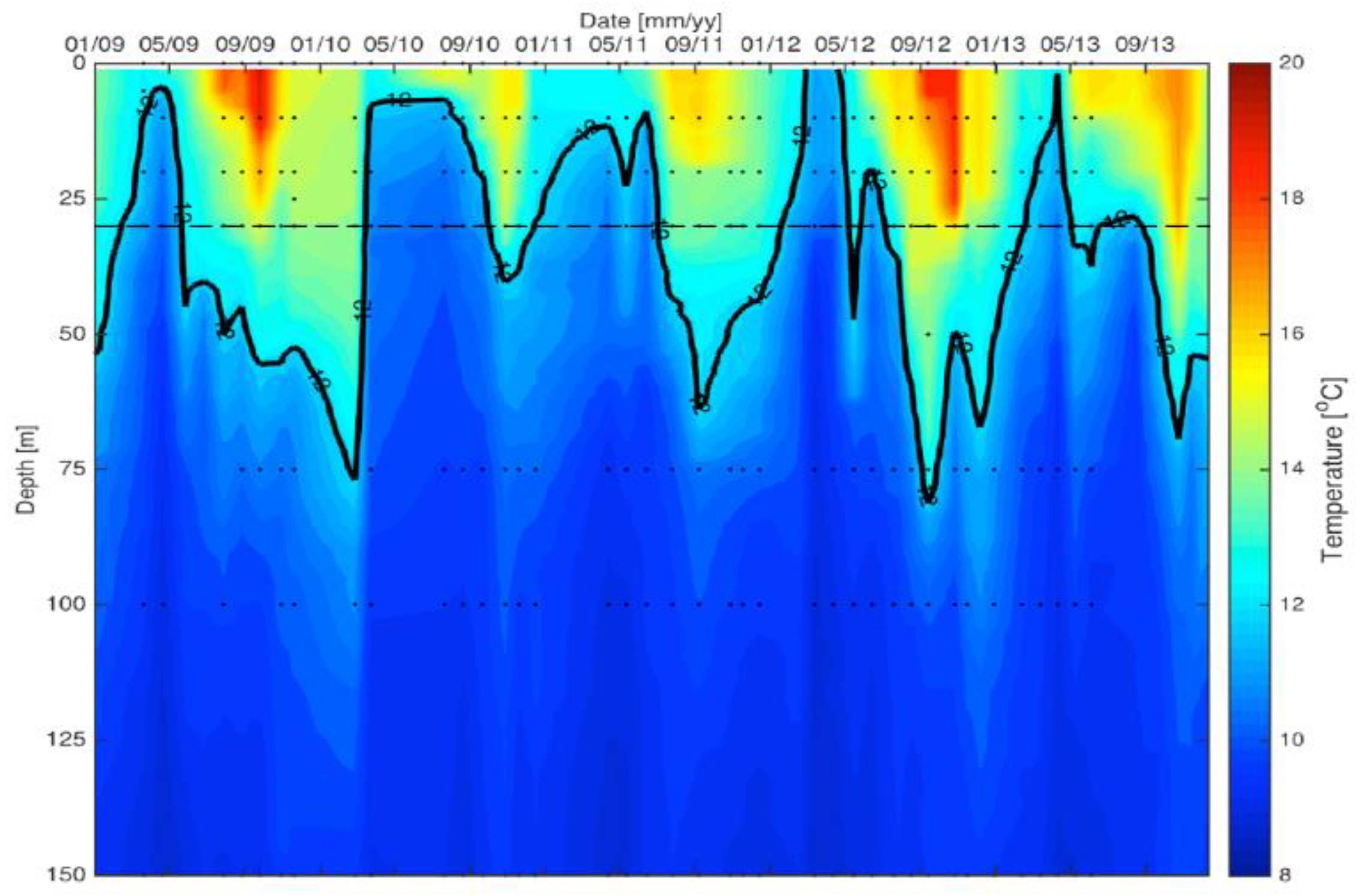
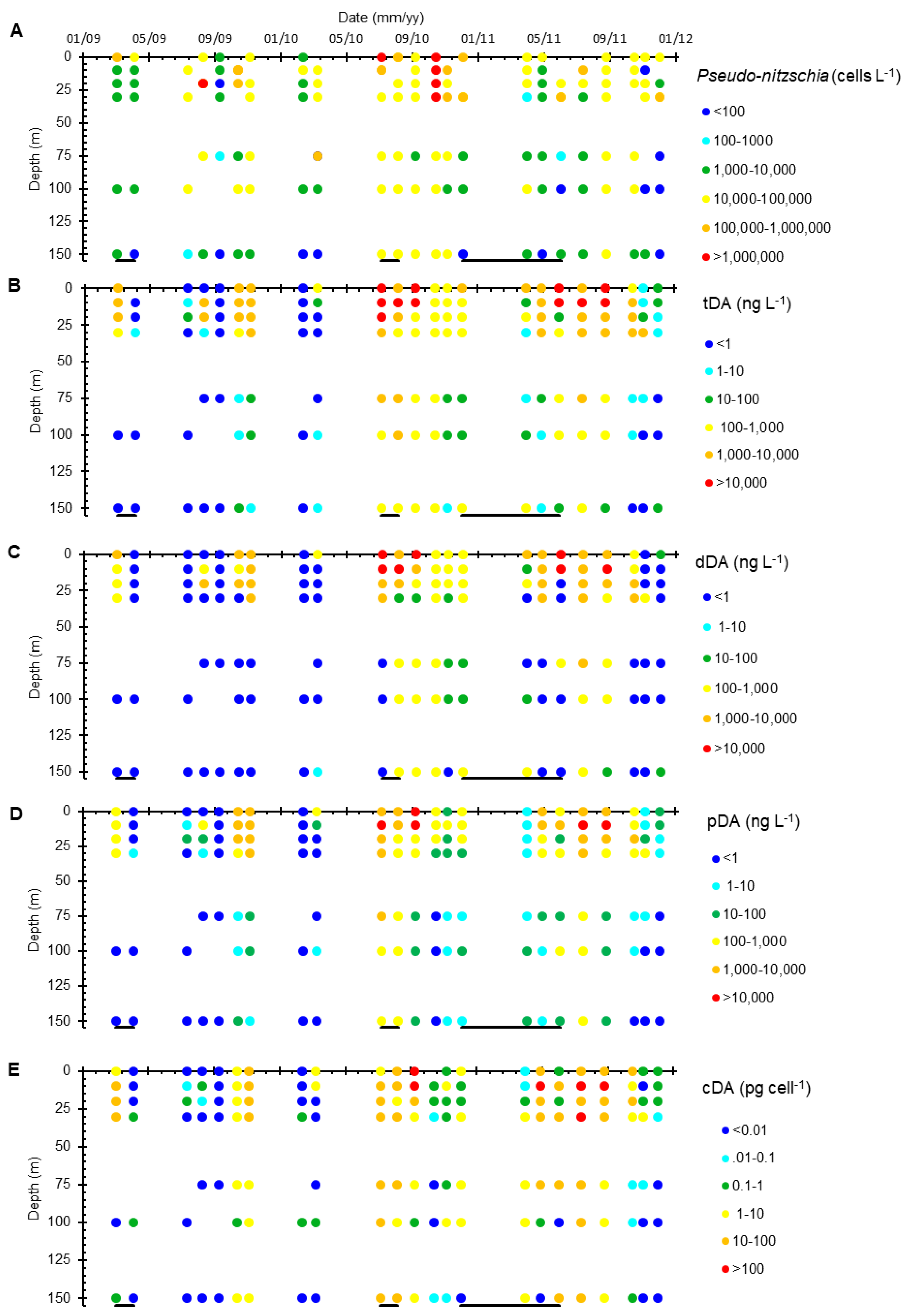
2.1. Water Column
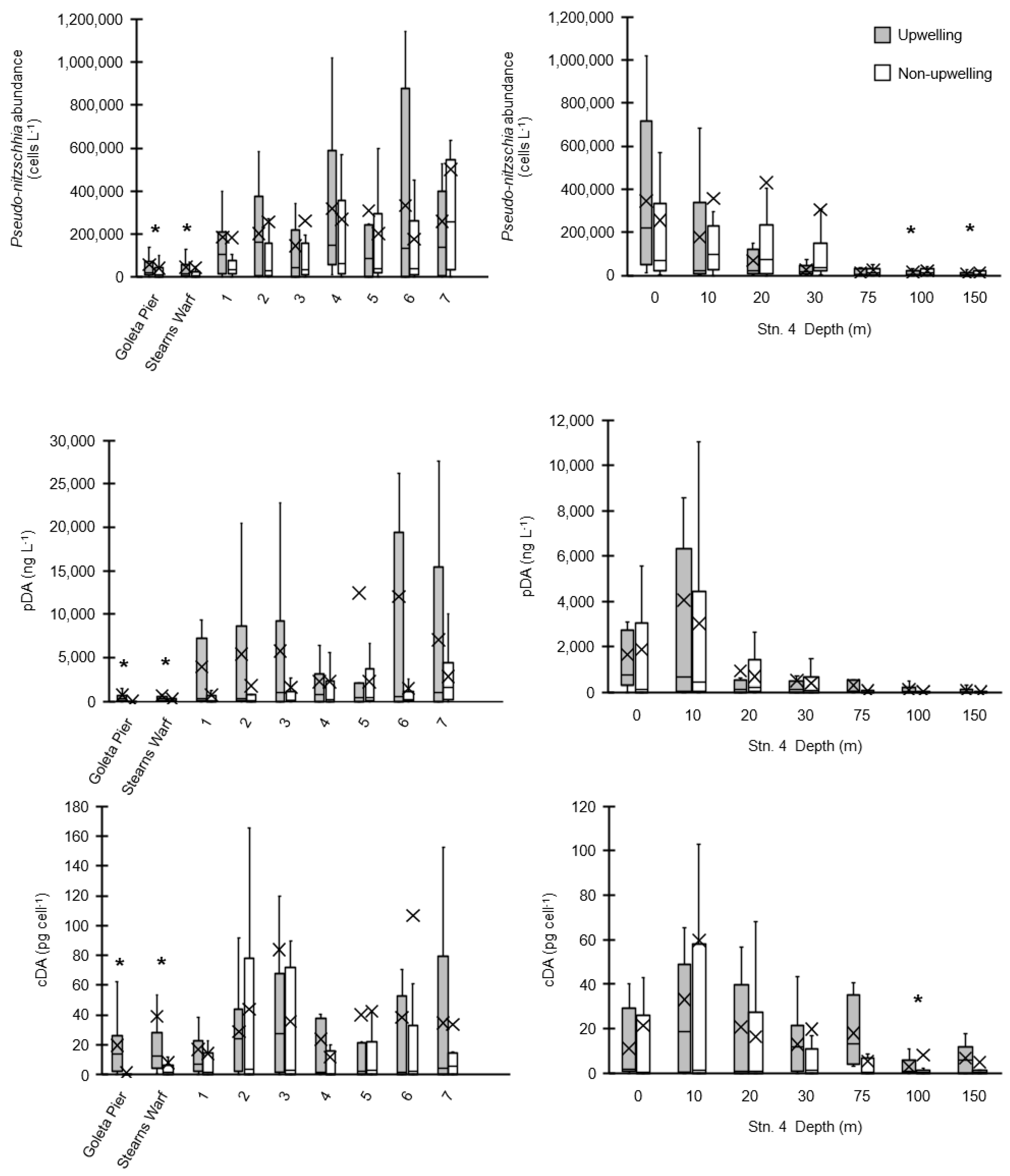
2.1.1. Surface Waters
2.1.2. Depth Distributions
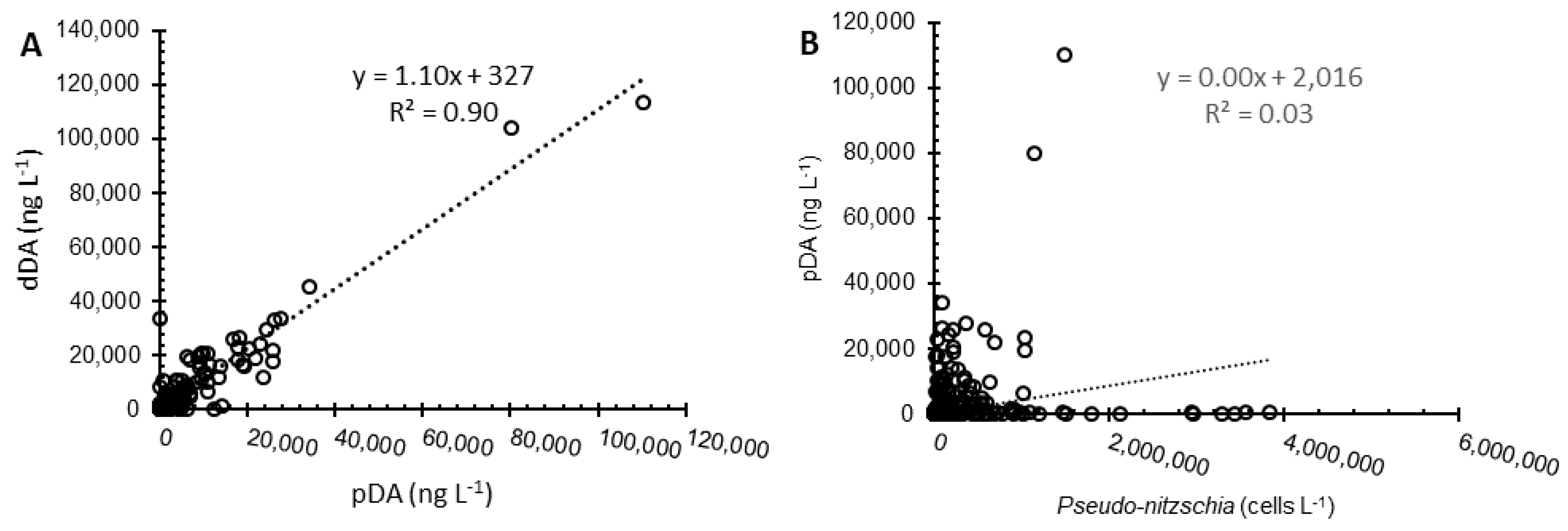
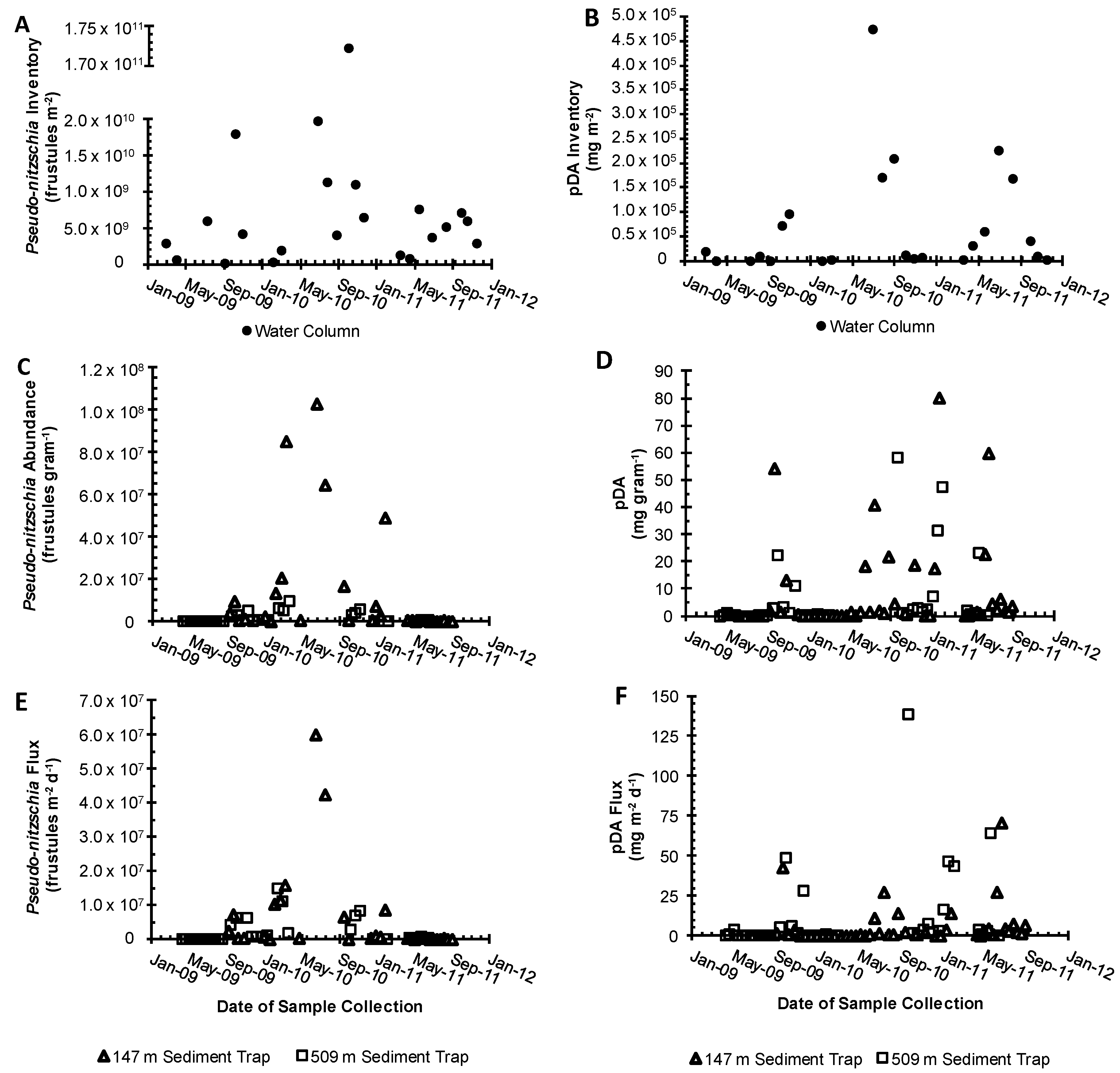
2.2. Water Column Inventories and Flux Comparison
2.3. Sediment Traps
3. Discussion
3.1. Water Column
3.2. Surface to Depth Transport
4. Conclusions
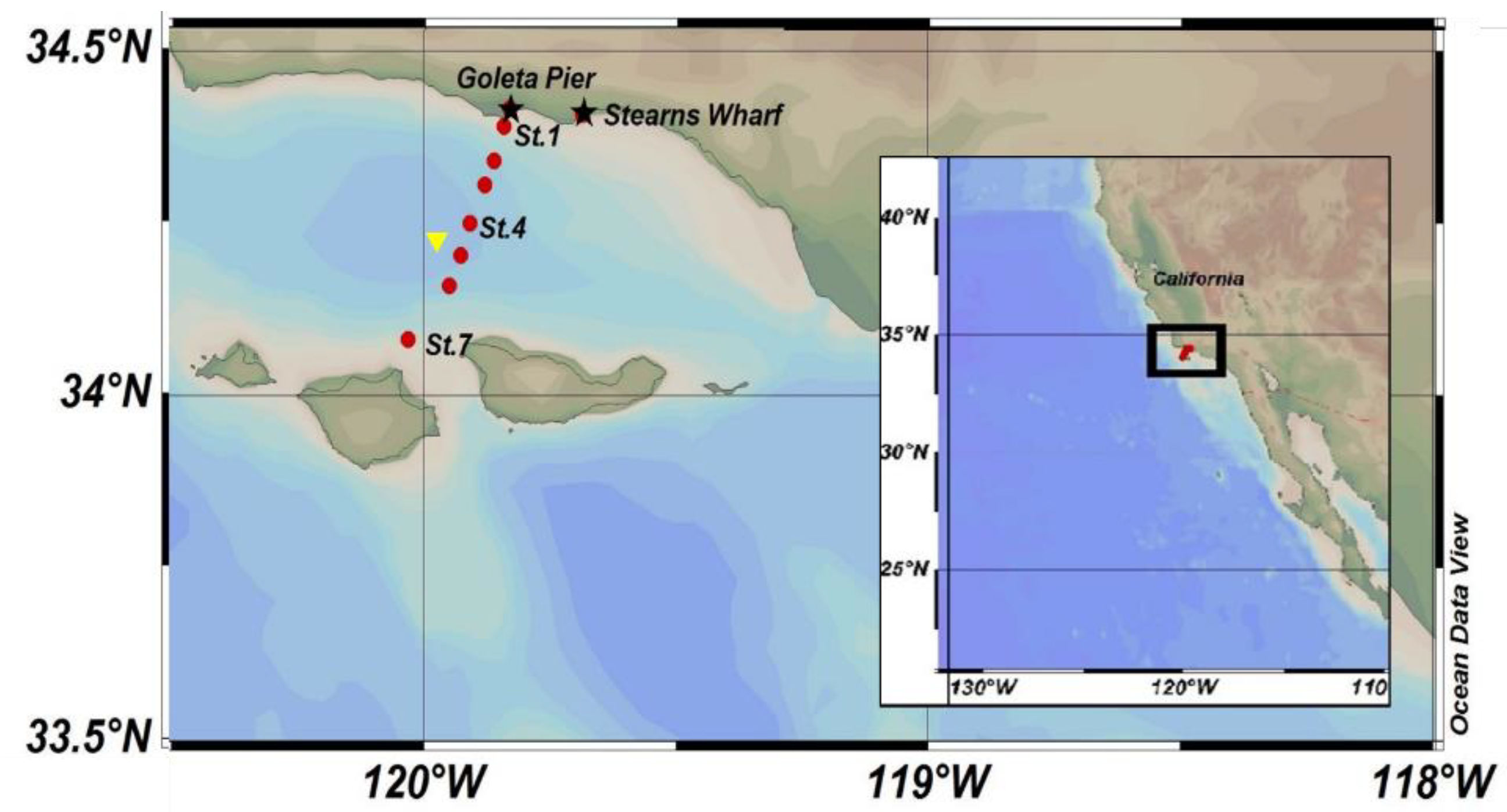
5. Materials and Methods
5.1. Pier Stations
5.2. Offshore Stations
5.3. Sediment Traps
5.4. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Robertson, A.; Frame, E.R.; Colegrove, K.M.; Nance, S.; Baugh, K.A.; Wiedenhoft, H.; Gulland, F.M.D. Clinical signs and histopathology associated with domoic acid poisoning in northern fur seals (Callorhinus ursinus) and comparison of toxin detection methods. Harmful Algae 2010, 9, 374–383. [Google Scholar] [CrossRef]
- Trainer, V.L.; Adams, N.G.; Bill, B.D.; Stehr, C.M.; Wekell, J.C.; Moeller, P.; Busman, M.; Woodruff, D.; Helena, M.; Sellner, K.; et al. Domoic acid production near California coastal upwelling zones, June 1998. Limnol. Oceanogr. 2000, 45, 1818–1833. [Google Scholar] [CrossRef] [Green Version]
- Bargu, S.; Powell, C.; Coale, S.; Busman, M.; Doucette, G.; Silver, M. Domoic acid detection in krill: A potential vector in marine food webs. Mar. Ecol. Prog. Ser. 2002, 237, 209–219. [Google Scholar] [CrossRef]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; Delong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Lett. Nat. 2000, 403, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, T.; Mazet, J.A.K.; Zabka, T.S.; Langlois, G.; Colegrove, K.M.; Silver, M.; Bargu, S.; Van Dolah, F.; Leighfield, T.; Conrad, P.A.; et al. Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): An increasing risk to marine mammal health. Proc. Biol. Sci. 2008, 275, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Schnetzer, A.; Jones, B.H.; Schaffner, R.A.; Cetinic, I.; Fitzpatrick, E.; Miller, P.E.; Seubert, E.L.; Caron, D.A. Coastal upwelling linked to toxic Pseudo-nitzschia australis blooms in Los Angeles coastal waters, 2005–2007. J. Plankton Res. 2013, 35, 1080–1092. [Google Scholar] [CrossRef]
- Bengtson Nash, S.M.; Baddock, M.C.; Takahashi, E.; Dawson, A.; Cropp, R. Domoic Acid Poisoning as a Possible Cause of Seasonal Cetacean Mass Stranding Events in Tasmania, Australia. Bull. Environ. Contam. Toxicol. 2017, 98, 8–13. [Google Scholar] [CrossRef] [PubMed]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Faustman, E.M. Domoic acid as a developmental neurotoxin. Neurotoxicology 2010, 31, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.H.; Andersen, M.E.; Basu, N.; Carvan, M.J.; Crofton, K.M.; King, K.A.; Suñol, C.; Tiffany-Castiglioni, E.; Schultz, I.R. Defining and modeling known adverse outcome pathways: Domoic acid and neuronal signaling as a case study. Environ. Toxicol. Chem. 2011, 30, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelong, A.; Hegaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia ( Bacillariophyceae ) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Ryan, J.P.; Kudela, R.M.; Birch, J.M.; Blum, M.; Bowers, H.A.; Chavez, F.P.; Doucette, G.J.; Hayashi, K.; Marin, R., III; Mikulski, C.M.; et al. Causality of an extreme harmful algal bloom in Monterey Bay, California, during the 2014–2016 northeast Pacific warm anomaly. Geophys. Res. Lett. 2017, 44, 5571–5579. [Google Scholar] [CrossRef]
- Macintyre, H.L.; Stutes, A.L.; Smith, W.L.; Dorsey, C.P.; Abraham, A.; Dickey, R.W. Environmental correlates of community composition and toxicity during a bloom of Pseudo-nitzschia spp. in the northern Gulf of Mexico. J. Plankton Res. 2011, 33, 273–295. [Google Scholar] [CrossRef]
- Anderson, C.R.; Brzezinski, M.A.; Washburn, L.; Kudela, R. Circulation and environmental conditions during a toxigenic Pseudo-nitzschia australis bloom in the Santa Barbara Channel, California. Mar. Ecol. Prog. Ser. 2006, 327, 119–133. [Google Scholar] [CrossRef]
- Anderson, C.R.; Siegel, D.A.; Kudela, R.M.; Brzezinski, M.A. Empirical models of toxigenic Pseudo-nitzschia blooms: Potential use as a remote detection tool in the Santa Barbara Channel. Harmful Algae 2009, 8, 478–492. [Google Scholar] [CrossRef]
- Schnetzer, A.; Miller, P.E.; Schaffner, R.A.; Stauffer, B.A.; Jones, B.H.; Weisberg, S.B.; DiGiacomo, P.M.; Berelson, W.M.; Caron, D.A. Blooms of Pseudo-nitzschia and domoic acid in the San Pedro Channel and Los Angeles harbor areas of the Southern California Bight, 2003–2004. Harmful Algae 2007, 6, 372–387. [Google Scholar] [CrossRef]
- Lane, J.Q.; Raimondi, P.T.; Kudela, R.M. Development of a logistic regression model for the prediction of toxigenic Pseudo-nitzschia blooms in monterey bay, California. Mar. Ecol. Prog. Ser. 2009, 383, 37–51. [Google Scholar] [CrossRef]
- Howard, M.D.A.; Cochlan, W.P.; Ladizinsky, N.; Kudela, R.M. Nitrogenous preference of toxigenic Pseudo-nitzschia australis (Bacillariophyceae) from field and laboratory experiments. Harmful Algae 2007, 6, 206–217. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G.; Cochlan, W.P.; Hughes, M.P.; Trainer, V.L. Domoic acid: The synergy of iron, copper, and the toxicity of diatoms. Limnol. Oceanogr. 2005, 50, 1908–1917. [Google Scholar] [CrossRef] [Green Version]
- Tatters, A.O.; Fu, F.X.; Hutchins, D.A. High CO2 and silicate limitation synergistically increase the toxicity of Pseudo-nitzschia fraudulenta. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Haroardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous relations in the arctic marine food web: Interactions between toxin producing Pseudo-nitzschia diatoms and Calanus copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sison-Mangus, M.P.; Jiang, S.; Tran, K.N.; Kudela, R.M. Host-specific adaptation governs the interaction of the marine diatom, Pseudo-nitzschia and their microbiota. ISME J. 2014, 8, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Sison-Mangus, M.P.; Jiang, S.; Kudela, R.M.; Mehic, S. Phytoplankton-associated bacterial community composition and succession during toxic diatom bloom and non-bloom events. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thorel, M.; Fauchot, J.; Morelle, J.; Raimbault, V.; Le Roy, B.; Miossec, C.; Kientz-Bouchart, V.; Claquin, P. Interactive effects of irradiance and temperature on growth and domoic acid production of the toxic diatom Pseudo-nitzschia australis (Bacillariophyceae). Harmful Algae 2014, 39, 232–241. [Google Scholar] [CrossRef]
- Schnetzer, A.; Lampe, R.H.; Benitez-Nelson, C.R.; Marchetti, A.; Osburn, C.L.; Tatters, A.O. Marine snow formation by the toxin-producing diatom, Pseudo-nitzschia australis. Harmful Algae 2017, 61, 23–30. [Google Scholar] [CrossRef]
- Bouillon, R.-C.; Knierim, T.L.; Kieber, R.J.; Skrabal, S.A.; Wright, J.L.C. Photodegradation of the algal toxin domoic acid in natural water matrices. Limnol. Oceanogr. 2006, 51, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, A.H.V.; McManus, M.A.; Cheriton, O.M.; Cowen, R.K.; Greer, A.T.; Kudela, R.M.; Ruttenberg, K.; Sevadjian, J. Hidden thin layers of toxic diatoms in a coastal bay. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 101, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Sekula-Wood, E.; Schnetzer, A.; Benitez-Nelson, C.R.; Anderson, C.; Berelson, W.M.; Brzezinski, M.A.; Burns, J.M.; Caron, D.A.; Cetinic, I.; Ferry, J.L.; et al. Rapid downward transport of the neurotoxin domoic acid in coastal waters. Nat. Geosci. 2009, 2, 272–275. [Google Scholar] [CrossRef]
- Hagström, J.A.; Granéli, E.; Maneiro, I.; Barreiro, A.; Petermann, A.; Svensen, C. Release and degradation of amnesic shellfish poison from decaying Pseudo-nitzschia multiseries in presence of bacteria and organic matter. Harmful Algae 2007, 6, 175–188. [Google Scholar] [CrossRef]
- Sekula-Wood, E.; Benitez-Nelson, C.; Morton, S.; Anderson, C.; Burrell, C.; Thunell, R. Pseudo-nitzschia and domoic acid fluxes in Santa Barbara Basin (CA) from 1993 to 2008. Harmful Algae 2011, 10, 567–575. [Google Scholar] [CrossRef]
- Maneiro, I.; Frangópulos, M.; Guisande, C.; Fernández, M.; Reguera, B.; Riveiro, I. Zooplankton as a potential vector of diarrhetic shellfish poisoning toxins through the food web. Mar. Ecol. Prog. Ser. 2000, 201, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.M.; Ferry, J.L. Adsorption of domoic acid to marine sediments and clays. J. Environ. Monit. 2007, 9, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Alldredge, A.L.; Gotschalk, C.C. Direct observations of the mass flocculation of diatom blooms: Characteristics, settling velocities and formation of diatom aggregates. Deep Sea Res. Part A. Oceanogr. Res. Pap. 1989, 36, 159–171. [Google Scholar] [CrossRef]
- Vigilant, V.L.; Silver, M.W. Domoic acid in benthic flatfish on the continental shelf of Monterey Bay, California, USA. Mar. Biol. 2007, 151, 2053–2062. [Google Scholar] [CrossRef]
- Kvitek, R.; Goldberg, J.; Smith, G.; Doucette, G.; Silver, M. Domoic acid contamination within eight representative species from the benthic food web of Monterey Bay, California, USA. Mar. Ecol. Prog. Ser. 2008, 367, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Kelly, M.S.; Campbell, D.A.; Dong, S.L.; Zhu, J.X.; Wang, S.F. Exposure to domoic acid affects larval development of king scallop Pecten maximus (Linnaeus, 1758). Aquat. Toxicol. 2007, 81, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Bargu, S.; Lefebvre, K.; Silver, M.W. Effect of dissolved domoic acid on the grazing rate of krill Euphausia pacifica. Mar. Ecol. Prog. Ser. 2006, 312, 169–175. [Google Scholar] [CrossRef]
- Hong, Z.; Zhang, Y.; Zuo, Z.; Zhu, R.; Gao, Y. Influences of Domoic Acid Exposure on Cardiac Development and the Expression of Cardiovascular Relative Genes in Zebrafish (Daniorerio) Embryos. J. Biochem. Mol. Toxicol. 2015, 29, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Van Meerssche, E.; Pinckney, J.L. The influence of salinity in the domoic acid effect on estuarine phytoplankton communities. Harmful Algae 2017, 69, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rue, E.; Bruland, K. Domoic acid binds iron and copper: A possible role for the toxin produced by the marine diatom Pseudo-nitzschia. Mar. Chem. 2001, 76, 127–134. [Google Scholar] [CrossRef]
- Prince, E.K.; Irmer, F.; Pohnert, G. Domoic acid improves the competitive ability of Pseudo-nitzschia delicatissima against the diatom Skeletonema marinoi. Mar. Drugs 2013, 11, 2398–2412. [Google Scholar] [CrossRef] [PubMed]
- Husson, B.; Hernández-Fariñas, T.; Le Gendre, R.; Schapira, M.; Chapelle, A. Two decades of Pseudo-nitzschia spp. blooms and king scallop (Pecten maximus) contamination by domoic acid along the French Atlantic and English Channel coasts: Seasonal dynamics, spatial heterogeneity and interannual variability. Harmful Algae 2016, 51, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Louw, D.C.; Doucette, G.J.; Voges, E. Annual patterns, distribution and long-term trends of Pseudo-nitzschia species in the northern Benguela upwelling system. J. Plankton Res. 2017, 39, 35–47. [Google Scholar] [CrossRef]
- McKibben, S.M.; Peterson, W.; Wood, A.M.; Trainer, V.L.; Hunter, M.; White, A.E. Climatic regulation of the neurotoxin domoic acid. PNAS 2017, 114, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odebrecht, C.; Ferrario, M.E.; Ciotti, A.M.; Kitzmann, D.; Moreira, M.O.; Hinz, F. The distribution of the diatom Pseudo-nitzshia off southern Brazil and relationships with oceanographic conditions. Harmful Algal Bloom. 2001, 42–45. [Google Scholar]
- Kudela, R.; Pitcher, G.; Probyn, T.; Figueiras, F.; Moita, T.; Trainer, V. Harmful Algal Blooms in Coastal Upwelling Systems. Oceanography 2005, 18, 184–197. [Google Scholar] [CrossRef]
- Fryxell, G.A.; Villac, M.C.; Shapiro, L.P. The occurrence of the toxic diatom genus Pseudo-nitzschia (Bacillariophyceae) on the West Coast of the USA, 1920-1996: A review. Phycologia 1997, 36, 419–437. [Google Scholar] [CrossRef]
- Barron, J.A.; Bukry, D.; Field, D.B.; Finney, B. Response of diatoms and silicoflagellates to climate change and warming in the California Current during the past 250 years and the recent rise of the toxic diatom Pseudo-nitzschia australis. Quat. Int. 2013, 310, 140–154. [Google Scholar] [CrossRef]
- Anderson, C.R.; Siegel, D.A.; Brzezinski, M.A.; Guillocheau, N. Controls on temporal patterns in phytoplankton community structure in the Santa Barbara channel, California. J. Geophys. Res. 2008, 113, 1–16. [Google Scholar] [CrossRef]
- Seubert, E.L.; Gellene, A.G.; Howard, M.D.A.; Connell, P.; Ragan, M.; Jones, B.H.; Runyan, J.; Caron, D.A. Seasonal and annual dynamics of harmful algae and algal toxins revealed through weekly monitoring at two coastal ocean sites off southern California, USA. Environ. Sci. Pollut. Res. 2013, 20, 6878–6895. [Google Scholar] [CrossRef] [PubMed]
- Kahru, M.; Kudela, R.M.; Manzano-Sarabia, M.; Greg Mitchell, B. Trends in the surface chlorophyll of the California Current: Merging data from multiple ocean color satellites. Deep. Res. Part II Top. Stud. Oceanogr. 2012, 77–80, 89–98. [Google Scholar] [CrossRef]
- Krause, J.W.; Brzezinski, M.A.; Siegel, D.A.; Thunell, R.C. Biogenic silica standing stock and export in the Santa Barbara Channel ecosystem. J. Geophys. Res. Ocean. 2013, 118, 736–749. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, M.A.; Washburn, L. Phytoplankton primary productivity in the Santa Barbara Channel: Effects of wind-driven upwelling and mesoscale eddies. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Hickey, B.M. The California current system—Hypotheses and facts. Prog. Oceanogr. 1979, 8, 191–279. [Google Scholar] [CrossRef]
- Lynn, R.J.; Simpson, J.J. The California Current System: The seasonal variability of its physical characteristics. J. Geophys. Res. 1987, 92, 12947–12966. [Google Scholar] [CrossRef]
- Hickey, B.M. Ideas and Observations on Progress in the Study of the Seas; The Sea; Wiley and Sons Inc.: New York, NY, USA, 1998; pp. 345–393. [Google Scholar]
- Bograd, S.J.; Buil, M.P.; Di Lorenzo, E.; Castro, C.G.; Schroeder, I.D.; Goericke, R.; Anderson, C.R.; Benitez-Nelson, C.; Whitney, F.A. Changes in source waters to the Southern California Bight. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 112, 42–52. [Google Scholar] [CrossRef]
- Henderikx Freitas, F.; Siegel, D.A.; Washburn, L.; Halewood, S.; Stassinos, E. Assessing controls on cross-shelf phytoplankton and suspended particle distributions using repeated bio-optical glider surveys. J. Geophys. Res. Ocean. 2016, 121, 7776–7794. [Google Scholar] [CrossRef]
- Horner, R.A.; Hickey, B.M.; Postel, J.R. Pseudo-nitzschia blooms and physical oceanography off Washington State, USA. South African J. Mar. Sci. 2000, 22, 299–308. [Google Scholar] [CrossRef]
- Marchetti, A.; Trainer, V.L.; Harrison, P.J. Environmental conditions and phytoplankton dynamics associated with Pseudo-nitzschia abundance and domoic acid in the Juan de Fuca eddy. Mar. Ecol. Prog. Ser. 2004, 281, 1–12. [Google Scholar] [CrossRef]
- Trainer, V.L.; Hickey, B.M.; Lessard, E.J.; Cochlan, W.P.; Trick, C.G.; Wells, M.L.; MacFadyen, A.; Moore, S.K. Variability of Pseudo-nitzschia and domoic acid in the Juan de Fuca eddy region and its adjacent shelves. Limnol. Oceanogr. 2009, 54, 289–308. [Google Scholar] [CrossRef]
- Anderson, C.R.; Kudela, R.M.; Kahru, M.; Chao, Y.; Rosenfeld, L.K.; Bahr, F.L.; Anderson, D.M.; Norris, T.A. Initial skill assessment of the California Harmful Algae Risk Mapping ( C-HARM ) system. Harmful Algae 2016, 59, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Peterson, W.; Fisher, J.; Hunter, M.; Peterson, J. Initiation and development of a toxic and persistent Pseudo-nitzschia bloom off the Oregon coast in spring/summer2015. PLoS ONE 2016, 11, e0163977. [Google Scholar] [CrossRef] [PubMed]
- Bialonski, S.; Caron, D.A.; Schloen, J.; Feudel, U.; Kantz, H.; Moorthi, S.D. Phytoplankton dynamics in the Southern California Bight indicate a complex mixture of transport and biology. J. Plankton Res. 2016, 0, 1–15. [Google Scholar] [CrossRef]
- Seegers, B.N.; Birch, J.M.; Iii, R.M.; Scholin, C.A.; Caron, D.A.; Seubert, E.L.; Howard, M.D.A.; Robertson, G.L.; Jones, B.H. Subsurface seeding of surface harmful algal blooms observed through the integration of autonomous gliders, moored environmental sample processors, and satellite remote sensing in southern California. Limnol. Oceanogr. 2015, 60, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Mcmanus, M.A.; Kudela, R.M.; Silver, M.W.; Steward, G.F.; Donaghay, P.L.; Sullivan, J.M. Cryptic Blooms: Are Thin Layers the Missing Connection? Estuaries Coasts J. CERF 2008, 31, 396–401. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Gotschalk, C.C. The relative contribution of marine snow of different origins to biological processes in coastal waters. Cont. Shelf Res. 1990, 10, 41–58. [Google Scholar] [CrossRef]
- Baugh, K.; Bush, J.; Bill, B.; Lefebvre, K.; Trainer, V. Estimates of specific toxicity of several Pseudo-nitzschia species from the Washington coast, based on culture and field studies. African J. Mar. Sci. 2006, 28, 403–407. [Google Scholar] [CrossRef]
- Dursun, F.; Ünlü, S.; Yurdun, T. Determination of Domoic Acid in Plankton Net Samples from Golden Horn Estuary, Turkey, Using HPLC with Fluorescence Detection. Bull. Environ. Contam. Toxicol. 2018, 100, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Thessen, A.E.; Bowers, H.A.; Stoecker, D.K. Intra- and interspecies differences in growth and toxicity of Pseudo-nitzschia while using different nitrogen sources. Harmful Algae 2009, 8, 792–810. [Google Scholar] [CrossRef]
- Martin-Jezequel, V.; Calu, G.; Candela, L.; Amzil, Z.; Jauffrais, T.; Sechet, V.; Weigel, P. Effects of organic and inorganic nitrogen on the growth and production of domoic acid by Pseudo-nitzschia multiseries and P. australis (bacillariophyceae) in culture. Mar. Drugs 2015, 13, 7067–7086. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Gellene, A.G.; Hubbard, K.A.; Bowers, H.A.; Kudela, R.M.; Hayashi, K.; Caron, D.A. Pseudo-nitzschia species composition varies concurrently with domoic acid concentrations during two different bloom events in the Southern California Bight. J. Plankton Res. 2018, 40, 29–45. [Google Scholar] [CrossRef]
- Lail, E.M.; Skrabal, S.A.; Kieber, R.J.; Wright, J.L.C. The role of particles on the biogeochemical cycling of domoic acid and its isomers in natural water matrices. Harmful Algae 2007, 6, 651–657. [Google Scholar] [CrossRef]
- Bates, S.S.; Léger, C.; Wells, M.L.; Hardy, K. Photodegradation of Domoic Acid. Proc. Eight Can. Work. Harmful Mar. Algae 2003, 30–35. [Google Scholar]
- Fisher, J.M.; Reese, J.G.; Pellechia, P.J.; Moeller, P.L. Role of Fe ( III ), Phosphate, Dissolved Organic Matter, and Nitrate during the Photodegradation of Domoic Acid in the Marine Environment. Environ. Sci. Technol. 2006, 40, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.E. Postconsumption domoic acid generation by the diatom Pseudo-nitzschia multiseries as a factor in depuration models. Can. J. Fish. Aquat. Sci. 2008, 65, 1797–1799. [Google Scholar] [CrossRef]
- Stewart, J.E.; Marks, L.J.; Gilgan, M.W.; Pfeiffer, E.; Zwicker, B.M. Microbial utilization of the neurotoxin domoic acid: Blue mussels (Mytilus edulis) and soft shell clams (Mya arenaria) as sources of the microorganisms. Can. J. Microbiol. 1998, 44, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.Q.; Roddam, C.M.; Langlois, G.W.; Kudela, R.M. Application of Solid Phase Adsorption Toxin Tracking ( SPATT ) for field detection of the hydrophilic phycotoxins domoic acid and saxitoxin in coastal California. Limnol. Oceanogr. Methods 2010, 8, 645–660. [Google Scholar] [CrossRef]
- Colas, F.; Crassous, M.; Laurent, S.; Litaker, R.W.; Rinnert, E.; Le Gall, E.; Lunven, M.; Delauney, L.; Compere, C. A surface plasmon resonance system for the underwater detection of domoic acid. Limnol. Oceanogr. Methods 2016, 14, 456–465. [Google Scholar] [CrossRef]
- Shipe, R.F.; Brzezinski, M.A. A time series study of silica production and flux in an eastern boundary region: Santa Barbara Basin, California. Global Biogeochem. Cycles 2001, 15, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Venrick, E.L.; Lange, C.B.; Reid, F.M.H.; Dever, E.P. Temporal patterns of species composition of siliceous phytoplankton flux in the Santa Barbara Basin. J. Plankton Res. 2008, 30, 283–297. [Google Scholar] [CrossRef]
- Lundholm, N.; Clarke, a.; Ellegaard, M. A 100-year record of changing Pseudo-nitzschia species in a sill-fjord in Denmark related to nitrogen loading and temperature. Harmful Algae 2010, 9, 449–457. [Google Scholar] [CrossRef]
- Sekula-wood, E.; Benitez-nelson, C.R.; Bennett, M.A.; Thunell, R. Magnitude and composition of sinking particulate phosphorus fluxes in Santa Barbara Basin, California. Global Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef] [Green Version]
- Rines, J.E.B.; Donaghay, P.L.; Dekshenieks, M.M.; Sullivan, J.M.; Twardowski, M.S. Thin layers and camouflage: Hidden Pseudo-nitzschia spp. (Bacillariophyceae) populations in a fjord in the San Juan Islands, Washington, USA. Mar. Ecol. Prog. Ser. 2002, 225, 123–137. [Google Scholar] [CrossRef]
- Buesseler, K.O.; Antia, A.N.; Chen, M.; Fowler, S.W.; Gardner, W.D.; Gustafsson, O.; Harada, K.; Michaels, A.F.; van der Loeff, M.R.; Sarin, M.; et al. An assessment of the use of sediment traps for estimating upper ocean particle fluxes. J. Mar. Res. 2007, 65, 345–416. [Google Scholar] [CrossRef] [Green Version]
- Miesner, A.K.; Lundholm, N.; Krock, B.; Nielsen, T.G. The effect of Pseudo-nitzschia seriata on grazing and fecundity of Calanus finmarchicus and Calanus glacialis. J. Plankton Res. 2016, 38, 564–574. [Google Scholar] [CrossRef]
- Drapeau, D.T.; Dam, H.G.; Grenier, G. An Improved Flocculator Design for Use in Particle Aggregation Experiments. Limnol. Oceanogr. 1994, 39, 723–729. [Google Scholar] [CrossRef]
- Engel, A. The role of transparent exopolymer particles (TEP) in the increase in apparent particle stickiness (alpha) during the decline of a diatom bloom. J. Plankton Res. 2000, 22, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Alldredge, A.L.; Logan, B.E. The role of particulate carbohydrate exudates in the flocculation of diatom blooms. Deep. Res. Part I 1994, 41, 335–357. [Google Scholar] [CrossRef]
- Callahan, M. Gov. Brown Seeks Disaster Declaration over Dungeness Crab Closure. Available online: https://huffman.house.gov/media-center/in-the-news/gov-brown-seeks-disaster-declaration-over-dungeness-crab-closure (accessed on 17 November 2018).
- Hasle, G.R. The Inverted-Microscope Method. In Phytoplankton Manual; Sournia, A., Ed.; UNESCO: Paris, France, 1978; pp. 88–96. [Google Scholar]
- Hallegraeff, G.M.; Anderson, A.D.M. Intergovernmental Oceanographic Commission Manuals and Guides, 1995. Available online: http://unesdoc.unesco.org/images/0012/001220/122021eo.pdf (accessed on 17 November 2018).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Parameter | N | Min. | Max. | Median | Mean | Std. Dev. |
|---|---|---|---|---|---|---|---|
| Pier Stations | Pseudo-nitzschia (cells L−1) | 465 | BD | 8.03 × 105 | 7.15 × 103 | 4.08 × 104 | 8.97 × 104 |
| Narrow (cells L−1) | 463 | BD | 7.86 × 105 | 1.86 × 103 | 2.23 × 104 | 7.57 × 104 | |
| Wide (cells L−1) | 463 | BD | 3.77 × 105 | 928 | 1.86 × 104 | 4.74 × 104 | |
| Pseudo-nitzschia pDA (ng L−1) | 193 | BD | 9.33 × 103 | 94 | 409 | 1.04 × 103 | |
| Pseudo-nitzschia cDA (pg cell−1) | 182 | BD | 833 | 3.76 | 17.6 | 65.2 | |
| Narrow DA (pg cell−1) | 144 | BD | 2.14 × 103 | 4.97 | 92.7 | 274 | |
| Wide DA (pg cell−1) | 149 | BD | 833 | 10.77 | 26.5 | 72.7 | |
| Surface | Pseudo-nitzschia (cells L−1) | 223 | BD | 3.56 × 106 | 6.66 × 104 | 2.45 × 105 | 4.94 × 105 |
| Stations (1–7) | Pseudo-nitzschia % of Total | 223 | BD | 91.5% | 16.5% | 25.7% | 25.3% |
| Total Microphytoplankton (cells L−1) | 223 | 5.64 × 103 | 6.75 × 106 | 4.16 × 105 | 8.00 × 105 | 1.04 × 106 | |
| Chl a (ng L−1) | 251 | 83.4 | 2.76 × 104 | 1.87 × 103 | 2.90 × 103 | 3.31 × 103 | |
| tDA (ng L−1) | 235 | BD | 2.23 × 105 | 762 | 7.82 × 103 | 2.22 × 104 | |
| pDA (ng L−1) | 224 | BD | 1.10 × 105 | 257 | 3.65 × 103 | 1.07 × 104 | |
| dDA (ng L−1) | 233 | BD | 1.13 × 105 | 276 | 4.38 × 103 | 1.21 × 104 | |
| cDA (pg cell−1) | 203 | BD | 1.44 × 103 | 1.86 | 37 | 124 | |
| Station 4 | Pseudo-nitzschia (cells L−1) | 178 | BD | 3.83 × 106 | 2.08 × 104 | 1.48 × 105 | 105 |
| (0–150 m) | Pseudo-nitzschia % of Total | 177 | BD | 98.1% | 10.1% | 21.5% | 26.0% |
| Total Microphytoplankton (cells L−1) | 177 | 3.60 × 103 | 5.11 × 106 | 2.14 × 105 | 5.07 × 105 | 8.37 × 105 | |
| Chl a (ng L−1) | 187 | 20 | 1.18 × 104 | 1.35 × 103 | 2.22 × 103 | 2.38 × 103 | |
| tDA (ng L−1) | 243 | BD | 4.27 × 104 | 71.46 | 2.69 × 103 | 6.81 × 103 | |
| pDA (ng L−1) | 234 | BD | 2.18 × 104 | 19.32 | 1.17 × 103 | 3.12 × 103 | |
| dDA (ng L−1) | 242 | BD | 2.61 × 104 | 1.04 | 1.58 × 103 | 4.02 × 103 | |
| cDA(pg cell−1) | 159 | BD | 323 | 1.13 | 18.3 | 45.6 | |
| 147 m Trap | Pseudo-nitzschia (cells g sed.−1). | 31 | 1.17 × 103 | 1.03 × 108 | 1.74 × 105 | 1.21 × 107 | 2.26 × 107 |
| Pseudo-nitzschia flux (cells m−2 d−1) | 31 | 523 | 6.00 × 107 | 9.49 × 104 | 5.37 × 106 | 1.31 × 107 | |
| DA concentration (µg g sed.−1) | 46 | BD | 79.9 | 11.6 | 8.4 | 17.3 | |
| DA flux (µg m−2 d−1) | 46 | BD | 70.8 | 0.7 | 5.6 | 13.0 | |
| 509 m Trap | Pseudo-nitzschia (cells g sed.−1). | 22 | 8.22 × 103 | 9.54 × 106 | 4.31 × 105 | 2.04 × 106 | 2.64 × 106 |
| Pseudo-nitzschia flux (cells m−2 d−1) | 22 | 7.59 × 103 | 1.48 × 107 | 8.74 × 105 | 3.07 × 106 | 4.13 × 106 | |
| DA concentration (µg g sed.−1) | 28 | BD | 58.4 | 1.4 | 8.0 | 15.0 | |
| DA flux (µg m−2 d−1) | 28 | BD | 139.6 | 2.6 | 15.2 | 30.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umhau, B.P.; Benitez-Nelson, C.R.; Anderson, C.R.; McCabe, K.; Burrell, C. A Time Series of Water Column Distributions and Sinking Particle Flux of Pseudo-Nitzschia and Domoic Acid in the Santa Barbara Basin, California. Toxins 2018, 10, 480. https://doi.org/10.3390/toxins10110480
Umhau BP, Benitez-Nelson CR, Anderson CR, McCabe K, Burrell C. A Time Series of Water Column Distributions and Sinking Particle Flux of Pseudo-Nitzschia and Domoic Acid in the Santa Barbara Basin, California. Toxins. 2018; 10(11):480. https://doi.org/10.3390/toxins10110480
Chicago/Turabian StyleUmhau, Blaire P., Claudia R. Benitez-Nelson, Clarissa R. Anderson, Kelly McCabe, and Christopher Burrell. 2018. "A Time Series of Water Column Distributions and Sinking Particle Flux of Pseudo-Nitzschia and Domoic Acid in the Santa Barbara Basin, California" Toxins 10, no. 11: 480. https://doi.org/10.3390/toxins10110480
APA StyleUmhau, B. P., Benitez-Nelson, C. R., Anderson, C. R., McCabe, K., & Burrell, C. (2018). A Time Series of Water Column Distributions and Sinking Particle Flux of Pseudo-Nitzschia and Domoic Acid in the Santa Barbara Basin, California. Toxins, 10(11), 480. https://doi.org/10.3390/toxins10110480