OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Actinobacterial Strains for the Prevention and the Detoxification of OTA
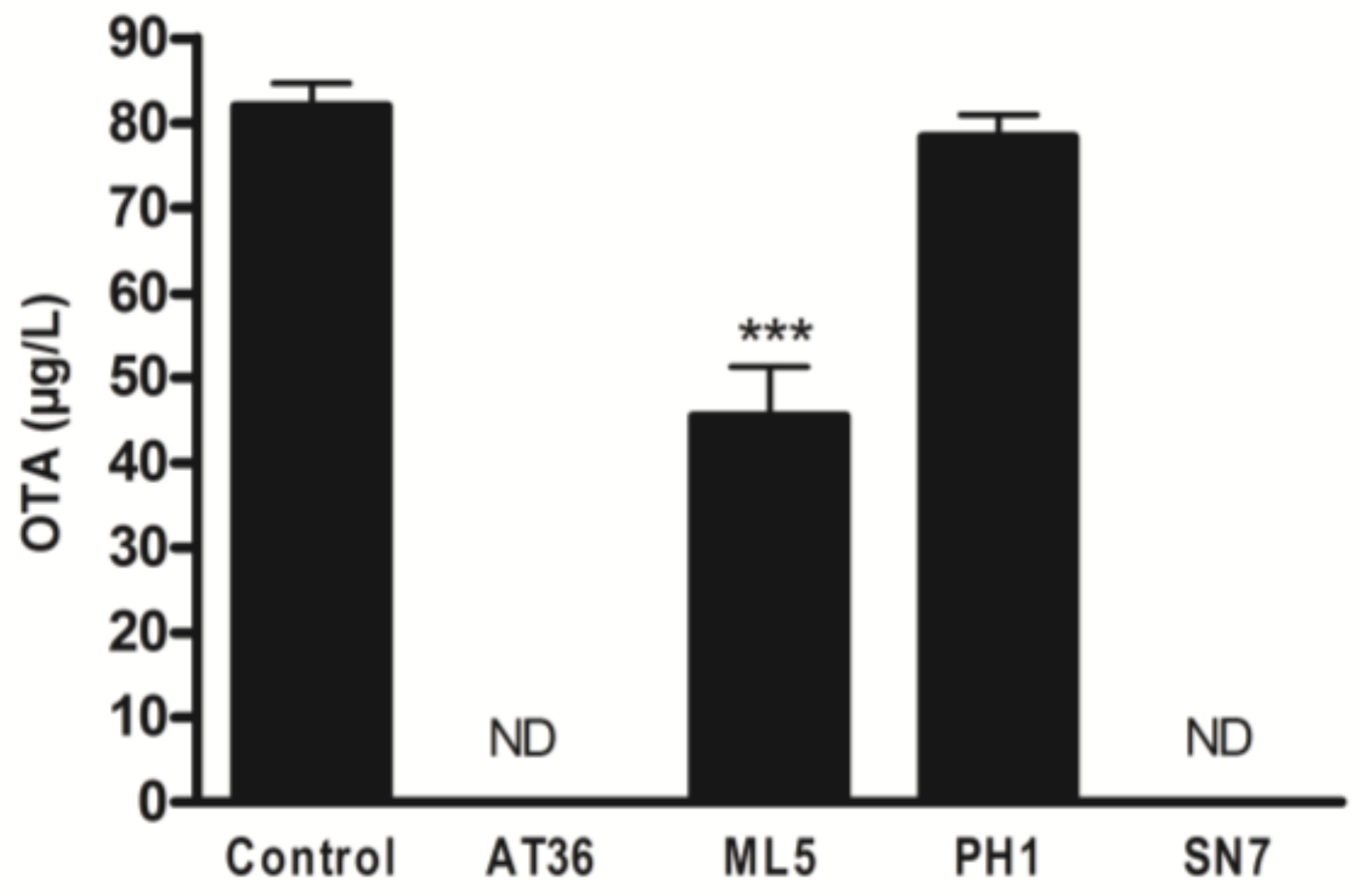
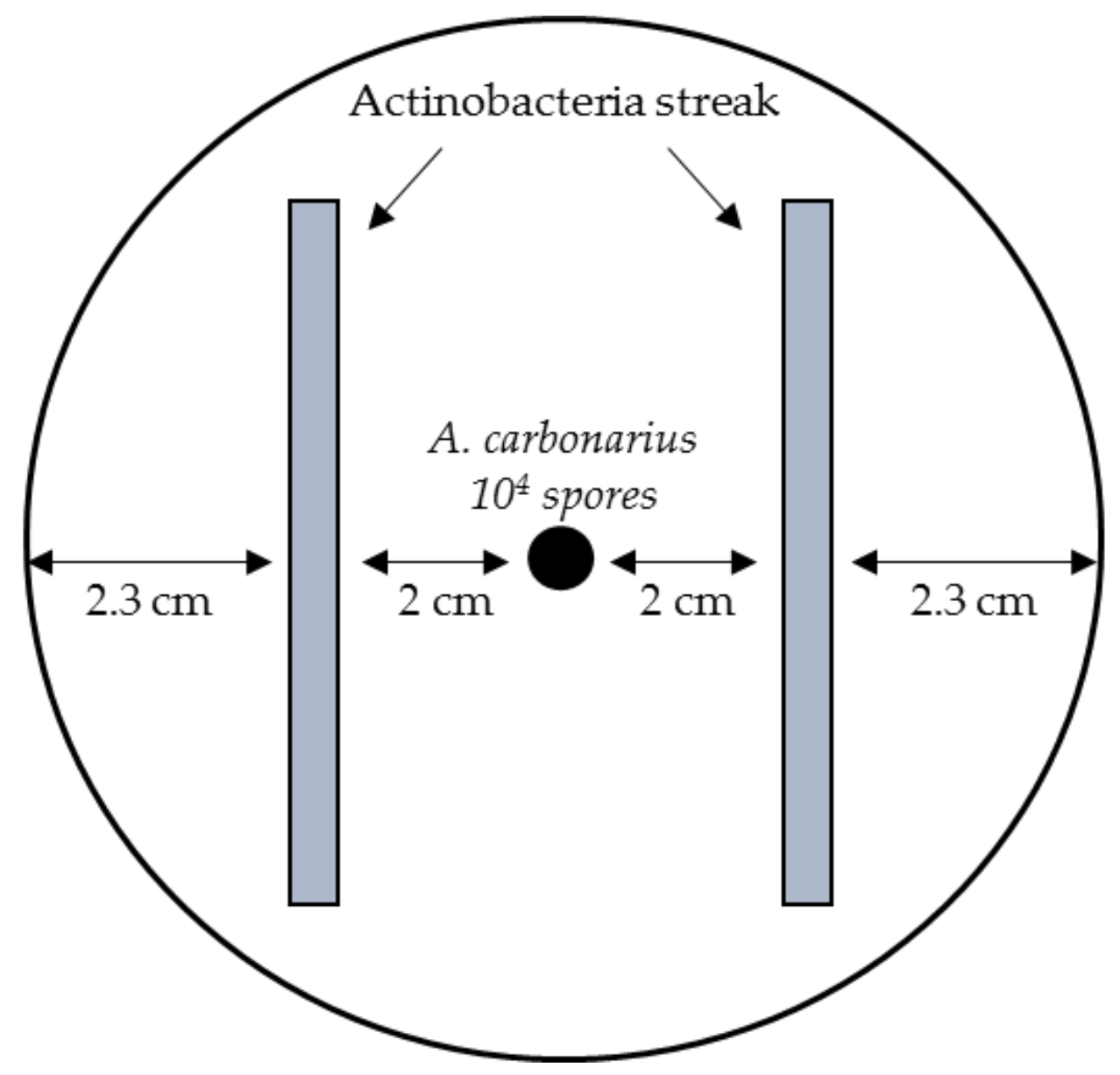
2.1.1. Screening of the Ability of Different Actinobacterial Strains to Reduce OTA Concentration
2.1.2. Transcriptomic Regulation of OTA Production by Actinobacteria in Solid Co-Culture with A. carbonarius
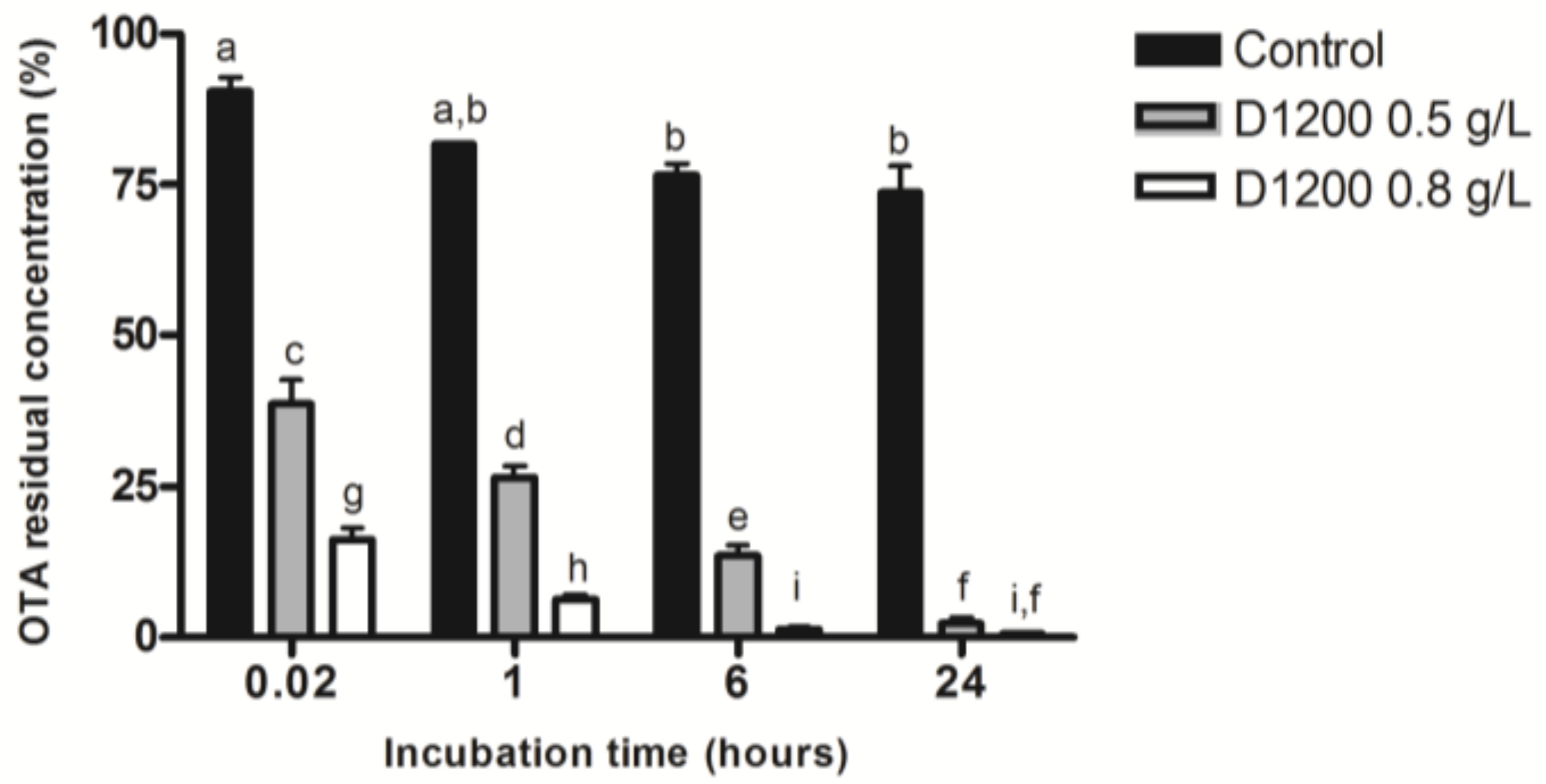
2.1.3. OTA Degradation and Adsorption Assays
2.2. Removal of OTA via Adsorption on Activated Carbon Fibers (ACFs)
2.2.1. Textural Characterization of ACFs
2.2.2. OTA Adsorption Depends on ACFs
2.2.3. Effect of ACFs Concentration on OTA Adsorption in AWM
2.2.4. Effect of OTA Concentration on Its Adsorption by ACFs in AWM
2.2.5. OTA Adsorption by ACFs in Red Grape Juice and Red Wine
3. Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Strains, Media, and Culture Conditions
4.3. Co-Culture of Actinobacterial Strains and A. Carbonarius
4.4. OTA Degradation Assay by Actinobacteria
4.5. OTA Adsorption Assay on Actinobacteria Spores
4.6. Analysis of Relative Expression of the OTA Cluster Genes by Qrt-PCR
4.7. Activated Carbon Fibers (ACFs)
4.8. ACFs Characterization
4.9. Liquid Matrices for OTA Adsorption by Acfs
4.10. OTA Adsorption on Acfs in AWM, Red Grape Juice, and Red Wine
4.11. OTA Quantification by HPLC/FLD/DAD
4.12. Statistical Analysis
Author Contributions
Conflicts of Interest
References
- Ostry, V.; Malir, F.; Ruprich, J. Producers and important dietary sources of ochratoxin A and citrinin. Toxins 2013, 5, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- IARC. Ochratoxin A. IARC Monogr. Eval. Carcinog. Risk Chem. to Hum. 1993, 56, 489–521. [Google Scholar]
- European Commission (EC). Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Commun. 2006, L364, 5–24. [Google Scholar] [CrossRef]
- Bellver Soto, J.; Fernández-Franzón, M.; Ruiz, M.J.; Juan-García, A. Presence of ochratoxin A (OTA) mycotoxin in alcoholic drinks from southern european countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M.; Venâncio, A.; Lima, N.; Guilloux-Bénatier, M.; Rousseaux, S. Predominant mycotoxins, mycotoxigenic fungi and climate change related to wine. Food Res. Int. 2017, 103, 478. [Google Scholar] [CrossRef] [PubMed]
- Şen, L.; Ocak, İ.; Nas, S.; Şevik, R. Effects of different drying treatments on fungal population and ochratoxin A occurrence in sultana type grapes. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.; Patiño, B. Wine Contamination with Ochratoxins: A Review. Beverages 2018, 4, 6. [Google Scholar] [CrossRef]
- Barberis, M.G.; Merlera, G.G.; Reynoso, M.M.; Chulze, S.N.; Torres, A.M. Factors affecting distribution and abundance of Aspergillus section Nigri in vineyard soils from grapevine growing regions of Argentina. J. Sci. Food Agric. 2014, 94, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; El Khoury, R.; Medina, Á.; Lippi, Y.; Naylies, C.; Atoui, A.; El Khoury, A.; Oswald, I.P.; Bailly, J.D.; Puel, O. Deciphering the anti-aflatoxinogenic properties of eugenol using a large-scale q-PCR approach. Toxins 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuda, S.; Yoshinari, T.; Furukawa, T.; Jermnak, U.; Takagi, K.; Iimura, K.; Yamamoto, T.; Suzuki, M. Search for aflatoxin and trichothecene production inhibitors and analysis of their modes of action. Biosci. Biotechnol. Biochem. 2016, 80, 43–54. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, R.; Caceres, I.; Puel, O.; Bailly, S.; Atoui, A.; Oswald, I.P.; El Khoury, A.; Bailly, J.D. Identification of the anti-aflatoxinogenic activity of Micromeria graeca and elucidation of its molecular mechanism in Aspergillus flavus. Toxins 2017, 9, 87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Apaliya, M.T.; Mahunu, G.K.; Chen, L.; Li, W. Control of ochratoxin A-producing fungi in grape berry by microbial antagonists: A review. Trends Food Sci. Technol. 2016, 51, 88–97. [Google Scholar] [CrossRef]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin a detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, R.; Mathieu, F.; Atoui, A.; Kawtharani, H.; El Khoury, A.; Afif, C.; Maroun, R.G.; El Khoury, A. Ability of soil isolated actinobacterial strains to prevent, bind and biodegrade ochratoxin A. Toxins 2017, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Verheecke, C.; Liboz, T.; Anson, P.; Zhu, Y.; Mathieu, F. Streptomyces-Aspergillus flavus interactions: Impact on aflatoxin B accumulation. Food Addit. Contam. Part A. Chem. Anal. Control Expo. Risk Assess. 2015, 32, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Verheecke, C.; Liboz, T.; Anson, P.; Diaz, R.; Mathieu, F. Reduction of aflatoxin production by Aspergillus flavus and Aspergillus parasiticus in interaction with Streptomyces. Microbiology 2015, 161, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Niu, Y.; Ma, T.; Xu, P.; Huang, W.; Zhan, J. Determination, content analysis and removal efficiency of fining agents on ochratoxin A in Chinese wines. Food Control 2017, 73, 382–392. [Google Scholar] [CrossRef]
- Balcerek, M.; Pielech-Przybylska, K.; Patelski, P.; Dziekońska-Kubczak, U.; Jusel, T. Treatment with activated carbon and other adsorbents as an effective method for the removal of volatile compounds in agricultural distillates. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2017, 34, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Castellari, M.; Versari, A.; Fabiani, A.; Parpinello, G.P.; Galassi, S. Removal of ochratoxin A in red wines by means of adsorption treatments with commercial fining agents. J. Agric. Food Chem. 2001, 49, 3917–3921. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villaran, M.C.; De Armentia, L.I.; Elejalde, E. Ochratoxin a removal from red wine by several oenological fining agents: Bentonite, egg albumin, allergen-free adsorbents, chitin and chitosan. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar] [CrossRef]
- Espejo, F.J.; Armada, S. Effect of activated carbon on ochratoxin A reduction in “pedro Ximenez” sweet wine made from off-vine dried grapes. Eur. Food Res. Technol. 2009, 229, 255–262. [Google Scholar] [CrossRef]
- Olivares-Marín, M.; Del Prete, V.; Garcia-Moruno, E.; Fernández-González, C.; Macías-García, A.; Gómez-Serrano, V. The development of an activated carbon from cherry stones and its use in the removal of ochratoxin A from red wine. Food Control 2009, 20, 298–303. [Google Scholar] [CrossRef]
- Le Cloirec, P.; Brasquet, C.; Subrenat, E. The adsorption onto fibrous activated carbon applications to water and air treatments. ACS Div. Fuel Chem. 1996, 41, 379–381. [Google Scholar] [CrossRef]
- Pelekani, C.; Snoeyink, V. Competitive adsorption in natural water: Role of activated carbon pore size. Water Res. 1999, 33, 1209–1219. [Google Scholar] [CrossRef]
- Knudsen, G.R.; Dandurand, L.-M.C. Ecological Complexity and the Success of Fungal Biological Control Agents. Adv. Agric. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Park, D.; Kim, K.; Lim, S.M.; Yu, N.H.; Kim, S.; Kim, H.Y.; Jung, K.S.; Jang, J.Y.; Park, J.C.; et al. Characterization of Bacillus amyloliquefaciens DA12 showing potent antifungal activity against mycotoxigenic Fusarium species. Plant Pathol. J. 2017, 33, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Kuhn, Y.; Schmidt-Heydt, M.; Geisen, R.; Chulze, S.N. Effect of Kluyveromyces thermotolerans on polyketide synthase gene expression and ochratoxin accumulation by Penicillium and Aspergillus. World Mycotoxin J. 2013, 6, 291–297. [Google Scholar] [CrossRef]
- Pan, D.; Mionetto, A.; Tiscornia, S.; Bettucci, L. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum and deoxynivalenol production in wheat. Mycotoxin Res. 2015, 31, 137–143. [Google Scholar] [CrossRef] [PubMed]
- de Melo Pereira, G.V.; Beux, M.; Pagnoncelli, M.G.B.; Soccol, V.T.; Rodrigues, C.; Soccol, C.R. Isolation, selection and evaluation of antagonistic yeasts and lactic acid bacteria against ochratoxigenic fungus Aspergillus westerdijkiae on coffee beans. Lett. Appl. Microbiol. 2016, 62, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a Halogenase Involved in the Biosynthesis of Ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microbiol. 2016, 82, 5631–5641. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Shams-Ghahfarokhi, M.; Kalantari, S.; Razzaghi-Abyaneh, M. Terrestrial Bacteria from Agricultural Soils: Versatile Weapons against Aflatoxigenic Fungi Terrestrial Bacteria from Agricultural Soils: Versatile. In Aflatoxins—Recent Advances and Future Prospects; InTech: London, UK, 2013; pp. 23–39. ISBN 9789535109044. [Google Scholar]
- Tsitsigiannis, D.I.; Dimakopoulou, M.; Antoniou, P.P.; Tjamos, E.C. Biological control strategies of mycotoxigenic fungi and associated mycotoxins in Mediterranean basin crops. Phytopathol. Mediterr. 2012, 51, 158–174. [Google Scholar] [CrossRef]
- Cubaiu, L.; Abbas, H.; Dobson, A.D.W.; Budroni, M.; Migheli, Q. A Saccharomyces cerevisiae wine strain inhibits growth and decreases ochratoxin A biosynthesis by Aspergillus carbonarius and Aspergillus ochraceus. Toxins 2012, 4, 1468–1481. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Shan, S.; Liu, Q.; Wang, X.; Yu, F. Biocontrol of Aspergillus flavus on peanut kernels by use of a strain of marine Bacillus megaterium. Int. J. Food Microbiol. 2010, 139, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi-Abyaneh, M.; Shams-Ghahfarokhi, M.; Chang, P.-K. Aflatoxins: Mechanisms of Inhibition by Antagonistic Plants and Microorganisms. In Aflatoxins—Biochemistry and Molecular Biology; InTech: London, UK, 2011. [Google Scholar]
- Yoshinari, T.; Noda, Y.; Yoda, K.; Sezaki, H.; Nagasawa, H.; Sakuda, S. Inhibitory activity of blasticidin A, a strong aflatoxin production inhibitor, on protein synthesis of yeast: Selective inhibition of aflatoxin production by protein synthesis inhibitors. J. Antibiot. 2010, 63, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, T.; Sugita-Konishi, Y.; Ohnishi, T.; Terajima, J. Inhibitory activities of blasticidin S derivatives on aflatoxin production by Aspergillus flavus. Toxins 2017, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, T.; Akiyama, T.; Nakamura, K.; Kondo, T.; Takahashi, Y.; Muraoka, Y.; Nonomura, Y.; Nagasawa, H.; Sakuda, S. Dioctatin A is a strong inhibitor of aflatoxin production by Aspergillus parasiticus. Microbiology 2007, 153, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Paterson, R.R.; Venancio, A. Biodegradation of ochratoxin a for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlovsky, P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat. Toxins 1999, 7, 1–23. [Google Scholar] [CrossRef]
- De Curtis, F.; de Felice, D.V.; Ianiri, G.; De Cicco, V.; Castoria, R. Environmental factors affect the activity of biocontrol agents against ochratoxigenic Aspergillus carbonarius on wine grape. Int. J. Food Microbiol. 2012, 159, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, V.; Rodriguez, H.; Carrascosa, A.V.; de las Rivas, B.; Garcia-Moruno, E.; Muñoz, R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar] [CrossRef] [PubMed]
- Bejaoul, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Assaf, J.C.; Atoui, A.; El Khoury, A.; Chokr, A.; Louka, N. A comparative study of procedures for binding of aflatoxin M1 to Lactobacillus rhamnosus GG. Braz. J. Microbiol. 2017, 49, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Patharajan, S.; Reddy, K.R.N.; Karthikeyan, V.; Spadaro, D.; Lore, A.; Gullino, M.L.; Garibaldi, A. Potential of yeast antagonists on invitro biodegradation of ochratoxin A. Food Control 2011, 22, 290–296. [Google Scholar] [CrossRef]
- Fallou, H. Adsorption Sur des Tissus de Carbone Activé de Micropolluants Emergents à L’état de Traces Dans les Eaux: Traitements Multi-Echelles et Modélisation. Ph.D. Thesis, Ecole Nationale Supérieure de Chimie, Rennes, France, 2015. [Google Scholar]
- Giraudet, S.; Le Cloirec, P. Activated carbon filters for filtration–adsorption. In Activated Carbon Fiber and Textiles; Chen, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 211–243. [Google Scholar]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Kurtbay, H.M.; Bekçi, Z.; Merdivan, M.; Yurdakoç, K. Reduction of ochratoxin a levels in red wine by bentonite, modified bentonites, and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Gambuti, A.; Strollo, D.; Genovese, A.; Ugliano, M.; Ritieni, A.; Moio, L. Influence of enological practices on ochratoxin A concentration in wine. Am. J. Enol. Vitic. 2005, 56, 155–162. [Google Scholar]
- Al Mardini, F.; Legube, B. Effect of the adsorbate (Bromacil) equilibrium concentration in water on its adsorption on powdered activated carbon. Part 2: Kinetic parameters. J. Hazard. Mater. 2009, 170, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Fallou, H.; Cimetière, N.; Giraudet, S.; Wolbert, D.; Le Cloirec, P. Adsorption of pharmaceuticals onto activated carbon fiber cloths—Modeling and extrapolation of adsorption isotherms at very low concentrations. J. Environ. Manag. 2016, 166, 544–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Castilla, C.; López-Ramón, M.V.; Pastrana-Martínez, L.M.; Álvarez-Merino, M.A.; Fontecha-Cámara, M.A. Competitive adsorption of the herbicide fluroxypyr and tannic acid from distilled and tap water on activated carbons and their thermal desorption. Adsorption 2012, 18, 173–179. [Google Scholar] [CrossRef]
- Andriantsiferana, C.; Julcour-Lebigue, C.; Creanga-Manole, C.; Delmas, H.; Wilhelm, A.-M.; Ph, D. Competitive Adsorption of p-Hydroxybenzoic Acid and Phenol on Activated Carbon: Experimental Study and Modeling. J. Environ. Eng. 2013, 139, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, E.F.; Andriantsiferana, C.; Wilhelm, A.M.; Delmas, H. Competitive adsorption of phenolic compounds from aqueous solution using sludge-based activated carbon. Environ. Technol. 2011, 32, 1325–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunauer, S.; Emmett, P.H.; Teller, E. Adsorption of Gases in Multimolecular Layers. J. Am. Chem. Soc. 1938, 60, 309–319. [Google Scholar] [CrossRef]
- Horvath, G.; Kawazoe, K. Method for the calculation of effectiv pore size distribution in molecular sieve carbon. J. Chem. Eng. Jpn. 1983, 16, 470–475. [Google Scholar] [CrossRef]
- Barrett, E.P.; Joyner, L.G.; Halenda, P.P. The Determination of Pore Volume and Area Distributions in Porous Substances. I. Computations from Nitrogen Isotherms. J. Am. Chem. Soc. 1951, 73, 373–380. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | OTA µg/L ± SEM | OTA Recovery (%) | ||
|---|---|---|---|---|
| Filtrate | Water wash | Methanol wash | ||
| Control | 97.1 ± 3 | 0.8 ± 0.4 | 3.5 ± 0.1 | 101.4 ± 3.8 |
| AT36 | 105.7 ±2 | 2.1 ± 0.6 | 1.6 ± 0.3 | 109.4 ± 2.9 |
| ML5 | 111.3 ± 5 | 0.4 ± 0.4 | 0.1 ± 0.1 | 111.8 ± 5.7 |
| PH1 | 106.6 ± 1 | 1 ± 0.7 | 0.4 ± 0.3 | 108 ± 1.5 |
| SN7 | 107.4 ± 1 | 1.7 ± 0.5 | 0.2 ± 0.2 | 109.3 ± 2.2 |
| Sample | AC10 | AC15 | AC20 | D1200 | D800 |
|---|---|---|---|---|---|
| Supplier | KYNOL | DACARB | |||
| Reference | ACC-5092-10 | ACC-5092-15 | ACC-5092-20 | TIS-KIP-1200 | MOU-CS-800 |
| Precursor | Novoïd fiber | Phenolic resin | Polyurethane foam impregnated with activated carbon | ||
| BET surface area (m2/g) | 940 | 1758 | 2032 | 1428 | 40 |
| Mesoporous volume (cm3/g) | 0.037 | 0.074 | 0.105 | 0.050 | 0.049 |
| Microporous volume (cm3/g) | 0.358 (90% volume) | 0.681 (90% volume) | 0.780 (88% volume) | 0.542 (92% volume) | 0.008 (14% volume) |
| Size pore diameter distribution (nm) | 0.4–0.8 0.8–1.5 | 0.5–0.8 0.8–2.2 | 0.5–0.8 0.8–2.5 | 0.5–0.8 0.8–2 | - |
| Initial OTA Concentration | ACFs Type | OTA Residual Concentration (%) | ||
|---|---|---|---|---|
| Incubation Time | ||||
| 0.02 h | 1 h | 6 h | ||
| 2 μg/L | D1200 | 39.9 ± 1.3 | 19.0 ± 0.4 | 18.3 ± 1.1 |
| AC15 | 36.4 ± 1.3 | 24.8 ± 1.0 | 24.2 ± 1.2 | |
| AC20 | 31.4 ± 1.1 | 22.7 ± 0.5 | 18.5 ± 0.7 | |
| 50 μg/L | D1200 | 17.0 ± 0.1 | 4.8 ± 0.2 | 0.6 ± 0.07 |
| AC15 | 16.3 ± 0.61 | 6.5 ± 0.6 | 3.3 ± 0.23 | |
| AC20 | 20.0 ± 0.5 | 3.8 ± 0.5 | 0.9 ± 0.2 | |
| 200 μg/L | D1200 | 12.0 ± 0.38 | 7.4 ± 0.3 | 2.2 ± 0.4 |
| AC15 | 22.7 ± 0.1 | 12.4 ± 0.3 | 3.6 ± 0.7 | |
| AC20 | 18.0 ± 0.3 | 15.2 ± 0.6 | 14.6 ± 0.5 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Khoury, R.; Choque, E.; El Khoury, A.; Snini, S.P.; Cairns, R.; Andriantsiferana, C.; Mathieu, F. OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results. Toxins 2018, 10, 137. https://doi.org/10.3390/toxins10040137
El Khoury R, Choque E, El Khoury A, Snini SP, Cairns R, Andriantsiferana C, Mathieu F. OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results. Toxins. 2018; 10(4):137. https://doi.org/10.3390/toxins10040137
Chicago/Turabian StyleEl Khoury, Rhoda, Elodie Choque, Anthony El Khoury, Selma P. Snini, Robbie Cairns, Caroline Andriantsiferana, and Florence Mathieu. 2018. "OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results" Toxins 10, no. 4: 137. https://doi.org/10.3390/toxins10040137
APA StyleEl Khoury, R., Choque, E., El Khoury, A., Snini, S. P., Cairns, R., Andriantsiferana, C., & Mathieu, F. (2018). OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results. Toxins, 10(4), 137. https://doi.org/10.3390/toxins10040137