Effect of Vipera ammodytes ammodytes Snake Venom on the Human Cytokine Network

 , , ,
, , , {kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Treatment
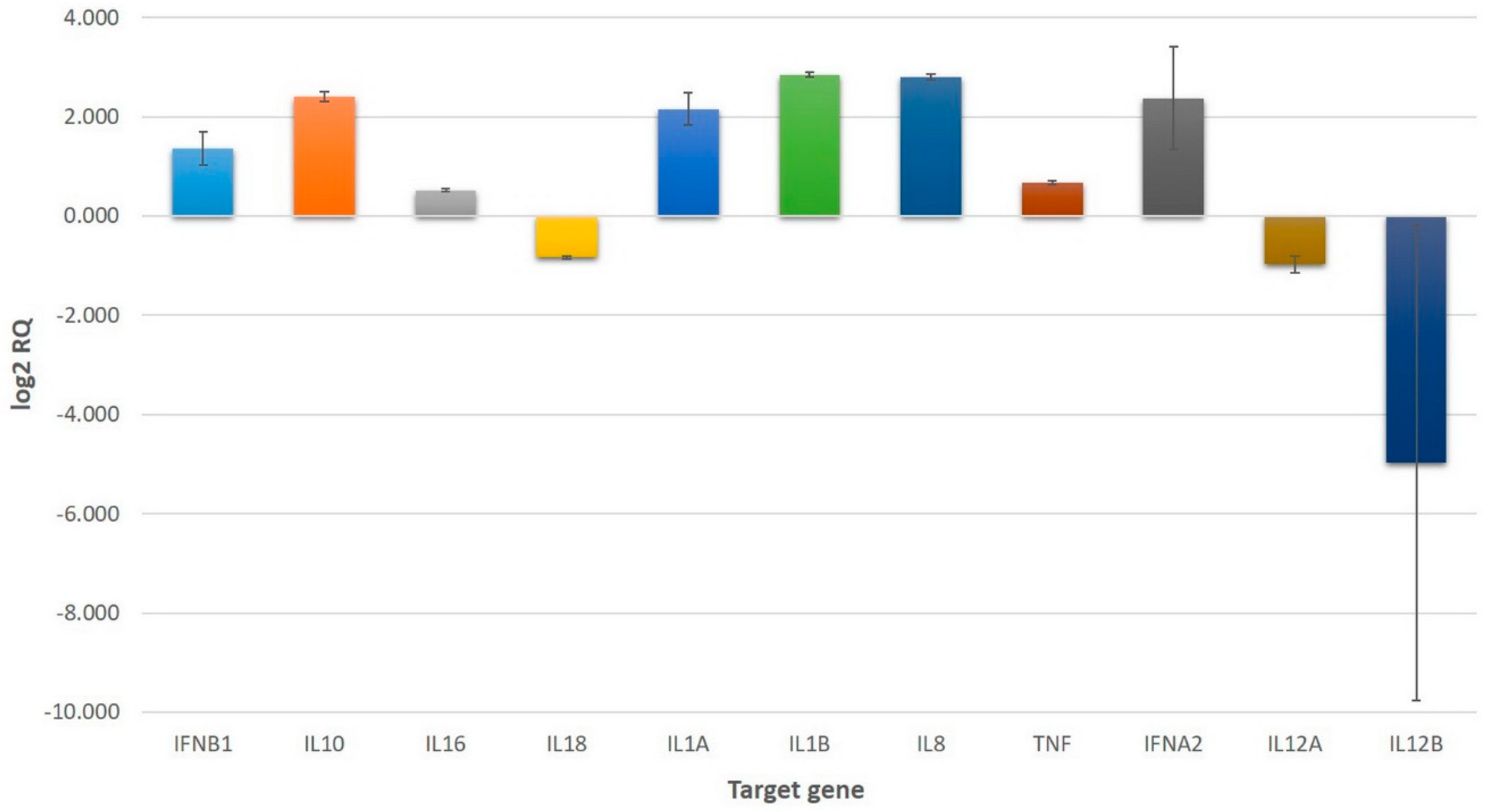
2.2. Gene Expression in U937 Cells Treated with VaaV
2.2.1. Upregulation of Interleukin-Related Genes
2.2.2. Downregulation of Interleukin-Related Genes
2.2.3. Upregulation of Chemokine-Related Genes
2.2.4. Upregulation of Interferon-Related Genes
2.2.5. Upregulation of Tumor Necrosis Factor-Related Genes
2.3. Limitations of the Study
3. Conclusions
4. Materials and Methods
4.1. Snake Venom
4.2. Cell Line
4.3. Treatment of Cells
4.4. RNA Isolation and cDNA Construction
4.5. RT-qPCR
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hakansson, A.; Molin, G. Gut microbiota and inflammation. Nutrients 2011, 3, 637–687. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.M.; Tan, Y.S.; Chua, K.H.; Sabaratnam, V.; Kuppusamy, U.R. Attenuation of Inflammatory Mediators (TNF-α and Nitric Oxide) and Up-Regulation of IL-10 by Wild and Domesticated Basidiocarps of Amauroderma rugosum (Blume & T. Nees) Torrend in LPS-Stimulated RAW264.7 Cells. PLoS ONE 2015, 10, e0139593. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Target Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Van Ginderachter, J.A.; Movahedi, K.; Hassanzadeh Ghassabeh, G.; Meerschaut, S.; Beschin, A.; Raes, G.; De Baetselier, P. Classical and alternative activation of mononuclear phagocytes: Picking the best of both worlds for tumor promotion. Immunobiology 2006, 211, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.F.; Mosser, D.M. A novel phenotype for an activated macrophage: The type 2 activated macrophage. J. Leukoc. Biol. 2002, 72, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Menaldo, D.L.; Bernardes, C.P.; Pereira, J.C.; Silveira, D.S.C.; Mamede, C.C.N.; Stanziola, L.; De Oliveira, F.; Pereira-Crott, L.S.; Faccioli, L.H.; Sampaio, S.V. Effects of two serine proteases from Bothrops pirajai snake venom on the complement system and the inflammatory response. Int. Immunopharmacol. 2013, 15, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.D.F.P.; Fernandes, C.M.; Zuliani, J.P.; Zamuner, S.F. Inflammatory effects of snake venom metalloproteinases. Mem. Inst. Oswaldo Cruz 2005, 100, 181–184. [Google Scholar] [CrossRef]
- Sunitha, K.; Hemshekhar, M.; Thushara, R.M.; Santhosh, M.S.; Sundaram, M.S.; Kemparaju, K.; Girish, K.S. Inflammation and oxidative stress in viper bite: An insight within and beyond. Toxicon 2015, 98, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.F.P.; Landucci, E.C.T.; Antunes, E.; Chacur, M.; Cury, Y. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003, 42, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.K.P.; Camargo, E.A.; Antunes, E. Inflammatory Action of Secretory Phospholipases A2 from Snake Venoms. In Toxins and Drug Discovery; Gopalakrishnakone, P., Cruz, L., Luo, S., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 35–52. ISBN 978-94-007-6451-4. [Google Scholar]
- Wanderley, C.W.S.; Silva, C.M.S.; Wong, D.V.T.; Ximenes, R.M.; Morelo, D.F.C.; Cosker, F.; Aragão, K.S.; Fernandes, C.; Palheta-Júnior, R.C.; Havt, A.; et al. Bothrops jararacussu snake venom-induces a local inflammatory response in a prostanoid- and neutrophil-dependent manner. Toxicon 2014, 90, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Menaldo, D.L.; Bernardes, C.P.; Zoccal, K.F.; Jacob-Ferreira, A.L.; Costa, T.R.; Del Lama, M.P.F.M.; Naal, R.M.Z.G.; Frantz, F.G.; Faccioli, L.H.; Sampaio, S.V. Immune cells and mediators involved in the inflammatory responses induced by a P-I metalloprotease and a phospholipase A2 from Bothrops atrox venom. Mol. Immunol. 2017, 85, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Deka, A.; Sharma, M.; Sharma, M.; Mukhopadhyay, R.; Doley, R. Purification and partial characterization of an anticoagulant PLA2 from the venom of Indian Daboia russelii that induces inflammation through upregulation of proinflammatory mediators. J. Biochem. Mol. Toxicol. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Gambero, A.; Thomazzi, S.M.; Cintra, A.C.O.; Landucci, E.C.T.; De Nucci, G.; Antunes, E. Signalling pathways regulating human neutrophil migration induced by secretory phospholipases A2. Toxicon 2004, 44, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Kanashiro, M.M.; De Escocard, R.C.M.; Petretski, J.H.; Prates, M.V.; Alves, E.W.; Machado, O.L.T.; Da Silva, W.D.; Kipnis, T.L. Biochemical and biological properties of phospholipases A2 from Bothrops atrox snake venom. Biochem. Pharmacol. 2002, 64, 1179–1186. [Google Scholar] [CrossRef]
- Landucci, E.C.T.; Toyama, M.; Marangoni, S.; Oliveira, B.; Cirino, G.; Antunes, E.; De Nucci, G. Effect of crotapotin and heparin on the rat paw oedema induced by different secretory phospholipases A2. Toxicon 2000, 38, 199–208. [Google Scholar] [CrossRef]
- Markland, F.S.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- De Toni, L.G.B.; Menaldo, D.L.; Cintra, A.C.O.; Figueiredo, M.J.; De Souza, A.R.; Maximiano, W.M.A.; Jamur, M.C.; Souza, G.E.P.; Sampaio, S.V. Inflammatory mediators involved in the paw edema and hyperalgesia induced by Batroxase, a metalloproteinase isolated from Bothrops atrox snake venom. Int. Immunopharmacol. 2015, 28, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, C.P.; Menaldo, D.L.; Mamede, C.C.N.; Zoccal, K.F.; Cintra, A.C.O.; Faccioli, L.H.; Stanziola, L.; de Oliveira, F.; Sampaio, S.V. Evaluation of the local inflammatory events induced by BpirMP, a metalloproteinase from Bothrops pirajai venom. Mol. Immunol. 2015, 68, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.M.; Zamuner, S.R.; Zuliani, J.P.; Rucavado, A.; Gutiérrez, J.M.; Teixeira, C.D.F.P. Inflammatory effects of BaP1 a metalloproteinase isolated from Bothrops asper snake venom: Leukocyte recruitment and release of cytokines. Toxicon 2006, 47, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Y.; Clemetson, K.J. Snake venom L-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Pontes, A.S.; da Setúbal, S.D.; Nery, N.M.; Da Silva, F.S.; Da Silva, S.D.; Fernandes, C.F.C.; Stábeli, R.G.; Soares, A.M.; Zuliani, J.P. P38 MAPK is involved in human neutrophil chemotaxis induced by L-amino acid oxidase from Calloselasma rhodosthoma. Toxicon 2016, 119, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.R.T.; Aissa, A.F.; Ribeiro, D.L.; Hernandes, L.C.; Machado, C.S.; Bianchi, M.L.P.; Sampaio, S.V.; Antunes, L.M.G. The toxin BjussuLAAO-II induces oxidative stress and DNA damage, upregulates the inflammatory cytokine genes TNF and IL6, and downregulates the apoptotic-related genes BAX, BCL2 and RELA in human Caco-2 cells. Int. J. Biol. Macromol. 2018, 109, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Menaldo, D.L.; Zoccal, K.F.; Burin, S.M.; Aissa, A.F.; de Castro, F.A.; Faccioli, L.H.; Greggi Antunes, L.M.; Sampaio, S.V. CR-LAAO, an L-amino acid oxidase from Calloselasma rhodostoma venom, as a potential tool for developing novel immunotherapeutic strategies against cancer. Sci. Rep. 2017, 7, 42673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, D.N.; Rypniewski, W.; Perbandt, M.; Jain, M.; Genov, N.; Betzel, C. Crystallization and preliminary X-ray diffraction studies of a toxic phospholipase A2from the venom of Vipera ammodytes meridionalis complexed to a synthetic inhibitor. Biochim. Biophys. Acta Proteins Proteom. 2003, 1650, 1–3. [Google Scholar] [CrossRef]
- Tomovic, L. Systematics of the nose-horned viper (Vipera ammodytes, Linnaeus, 1758). Herpetol. J. 2006, 16, 191–201. [Google Scholar]
- Maretić, T.; Cizelj, I.; Čivljak, R. Venomous snakebites and treatment—On the occassion of the acquisition of new venomous snakes at the Zagreb Zoo and private herpetaria. Infektoloski Glas. 2013, 33, 11–19. [Google Scholar]
- Radonić, V.; Budimir, D.; Bradarić, N.; Luksić, B.; Sapunar, D.; Vilović, K. Envenomation by the horned viper (Vipera ammodytes L.). Mil. Med. 1997, 162, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, D.; Risch, M.; Kardas, A.; Buck, F.; Von Bergen, M.; Betzel, C. Comparative analysis of the venom proteomes of Vipera ammodytes ammodytes and vipera ammodytes meridionalis. J. Proteome Res. 2008, 7, 866–886. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Sajevic, T.; Kovačič, L.; Pungerčar, J.; Lang Balija, M.; Halassy, B.; Trampuš Bakija, A.; Križaj, I. Hemorrhagin VaH4, a covalent heterodimeric P-III metalloproteinase from Vipera ammodytes ammodytes with a potential antitumour activity. Toxicon 2014, 77, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Kurtović, T.; Brgles, M.; Leonardi, A.; Balija, M.L.; Križaj, I.; Allmaier, G.; Marchetti-Deschmann, M.; Halassy, B. Ammodytagin, a heterodimeric metalloproteinase from Vipera ammodytes ammodytes venom with strong hemorrhagic activity. Toxicon 2011, 58, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Fox, J.W.; Trampuš-Bakija, A.; Križaj, I. Ammodytase, a metalloprotease from Vipera ammodytes ammodytes venom, possesses strong fibrinolytic activity. Toxicon 2007, 49, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Chaves, F.; Teixeira, C.F.P.; Gutiérrez, J.M. Role of TNF-α, IL-1β and IL-6 in the local tissue damage induced by Bothrops asper snake venom: An experimental assessment in mice. Toxicon 2005, 45, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, M.S.; Sundaram, M.S.; Sunitha, K.; Kemparaju, K.; Girish, K.S. Viper venom-induced oxidative stress and activation of inflammatory cytokines: A therapeutic approach for overlooked issues of snakebite management. Inflamm. Res. 2013, 62, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Corasolla Carregari, V.; Stuani Floriano, R.; Rodrigues-Simioni, L.; Winck, F.V.; Baldasso, P.A.; Ponce-Soto, L.A.; Marangoni, S. Biochemical, pharmacological, and structural characterization of new basic PLA2 Bbil-TX from Bothriopsis bilineata snake venom. Biomed. Res. Int. 2013, 2013, 612649. [Google Scholar] [CrossRef] [PubMed]
- De Waal Malefyt, R.; Abrams, J.; Bennet, B.; Figdor, C.G.; de Vries, J.E. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.F.; Isbister, G.K.; Shahmy, S.; Mohamed, F.; Abeysinghe, C.; Karunathilake, H.; Ariaratnam, A.; Jacoby-Alner, T.E.; Cotterell, C.L.; Brown, S.G.A. Immune Response to Snake Envenoming and Treatment with Antivenom; Complement Activation, Cytokine Production and Mast Cell Degranulation. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Hernández Cruz, A.; Garcia-Jimenez, S.; Zucatelli Mendonça, R.; Petricevich, V.L. Pro- and anti-inflammatory cytokines release in mice injected with Crotalus durissus terrificus venom. Mediat. Inflamm. 2008, 2008. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L.; Teixeira, C.F.P.; Tambourgi, D.V.; Gutiérrez, J.M. Increments in serum cytokine and nitric oxide levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef]
- McFadden, C.; Morgan, R.; Rahangdale, S.; Green, D.; Yamasaki, H.; Center, D.; Cruikshank, W. Preferential migration of T regulatory cells induced by IL-16. J. Immunol. 2007, 179, 6439–6445. [Google Scholar] [CrossRef] [PubMed]
- Moreira, V.; Dos-Santos, M.C.; Nascimento, N.G.; da Silva, H.B.; Fernandes, C.M.; D’Império Lima, M.R.; Teixeira, C. Local inflammatory events induced by Bothrops atrox snake venom and the release of distinct classes of inflammatory mediators. Toxicon 2012, 60, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Nunes, D.C.O.; Rodrigues, R.S.; Lucena, M.N.; Cologna, C.T.; Oliveira, A.C.S.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Arantes, E.C.; Teixeira, D.N.S.; Ueira-Vieira, C.; et al. Isolation and functional characterization of proinflammatory acidic phospholipase A2 from Bothrops leucurus snake venom. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.L.; Wei, J.F.; Li, T.; Qiao, L.Y.; Liu, Y.L.; Huang, T.; He, S.H. Purification, characterization and potent lung lesion activity of an l-amino acid oxidase from Agkistrodon blomhoffii ussurensis snake venom. Toxicon 2007, 50, 1126–1139. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.M.; Issekutz, T.B.; Mohan, K.; Lee, P.W.K.; Shmulevitz, M.; Marshall, J.S. Human mast cell activation with virus-associated stimuli leads to the selective chemotaxis of natural killer cells by a CXCL8-dependent mechanism. Blood 2008, 111, 5467–5476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Setubal, S.S.; Pontes, A.S.; Nery, N.M.; Bastos, J.S.F.; Castro, O.B.; Pires, W.L.; Zaqueo, K.D.; de Calderon, L.A.; Stábeli, R.G.; Soares, A.M.; et al. Effect of Bothrops bilineata snake venom on neutrophil function. Toxicon 2013, 76, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Meyer, O. Interferons and autoimmune disorders. Jt. Bone Spine 2009, 76, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Hervas-Stubbs, S.; Perez-Gracia, J.L.; Rouzaut, A.; Sanmamed, M.F.; Le Bon, A.; Melero, I. Direct effects of type I interferons on cells of the immune system. Clin. Cancer Res. 2011, 17, 2619–2627. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, V.D.M.; Russo, R.R.; Oliveira Cintra, A.C.; Sartim, M.A.; De Melo Alves-Paiva, R.; Figueiredo, L.T.M.; Sampaio, S.V.; Aquino, V.H. Crotoxin and phospholipases A2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon 2012, 59, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecilio, A.B.; Caldas, S.; De Oliveira, R.A.; Santos, A.S.B.; Richardson, M.; Naumann, G.B.; Schneider, F.S.; Alvarenga, V.G.; Estevão-Costa, M.I.; Fuly, A.L.; et al. Molecular characterization of Lys49 and Asp49 phospholipases A2 from snake venom and their antiviral activities against Dengue virus. Toxins 2013, 5, 1780–1798. [Google Scholar] [CrossRef] [PubMed]
- Ruddle, N.H. Lymphotoxin and TNF: How it all began—A tribute to the travelers. Cytokine Growth Factor Rev. 2014, 25, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.; Namineni, S.; Reisinger, F.; Zöller, J.; Yuan, D.; Heikenwälder, M. Lymphotoxin, NF-κB, and cancer: The dark side of cytokines. Dig. Dis. 2012, 30, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Gupta, S.C.; Kim, J.H. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 2012, 119, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Balogh, P. Developmental Biology of Peripheral Lymphoid Organs; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 978-3-642-14428-8. [Google Scholar]
- Clissa, P.B.; Laing, G.D.; Theakston, R.D.G.; Mota, I.; Taylor, M.J.; Moura-da-Silva, A.M. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on pro-inflammatory cytokines released by murine peritoneal adherent cells. Toxicon 2001, 39, 1567–1573. [Google Scholar] [CrossRef]
- Pontes, A.S.; da Setúbal, S.S.; Xavier, C.V.; Lacouth-Silva, F.; Kayano, A.M.; Pires, W.L.; Nery, N.M.; De Castro, O.B.; Da Silva, S.D.; Calderon, L.A.; et al. Effect of l-amino acid oxidase from Calloselasma rhodosthoma snake venom on human neutrophils. Toxicon 2014, 80, 27–37. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boda, F.; Banfai, K.; Garai, K.; Curticapean, A.; Berta, L.; Sipos, E.; Kvell, K. Effect of Vipera ammodytes ammodytes Snake Venom on the Human Cytokine Network. Toxins 2018, 10, 259. https://doi.org/10.3390/toxins10070259
Boda F, Banfai K, Garai K, Curticapean A, Berta L, Sipos E, Kvell K. Effect of Vipera ammodytes ammodytes Snake Venom on the Human Cytokine Network. Toxins. 2018; 10(7):259. https://doi.org/10.3390/toxins10070259
Chicago/Turabian StyleBoda, Francisc, Krisztina Banfai, Kitti Garai, Augustin Curticapean, Lavinia Berta, Emese Sipos, and Krisztian Kvell. 2018. "Effect of Vipera ammodytes ammodytes Snake Venom on the Human Cytokine Network" Toxins 10, no. 7: 259. https://doi.org/10.3390/toxins10070259
APA StyleBoda, F., Banfai, K., Garai, K., Curticapean, A., Berta, L., Sipos, E., & Kvell, K. (2018). Effect of Vipera ammodytes ammodytes Snake Venom on the Human Cytokine Network. Toxins, 10(7), 259. https://doi.org/10.3390/toxins10070259