Enterohemorrhagic E. coli (EHEC)—Secreted Serine Protease EspP Stimulates Electrogenic Ion Transport in Human Colonoid Monolayers


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
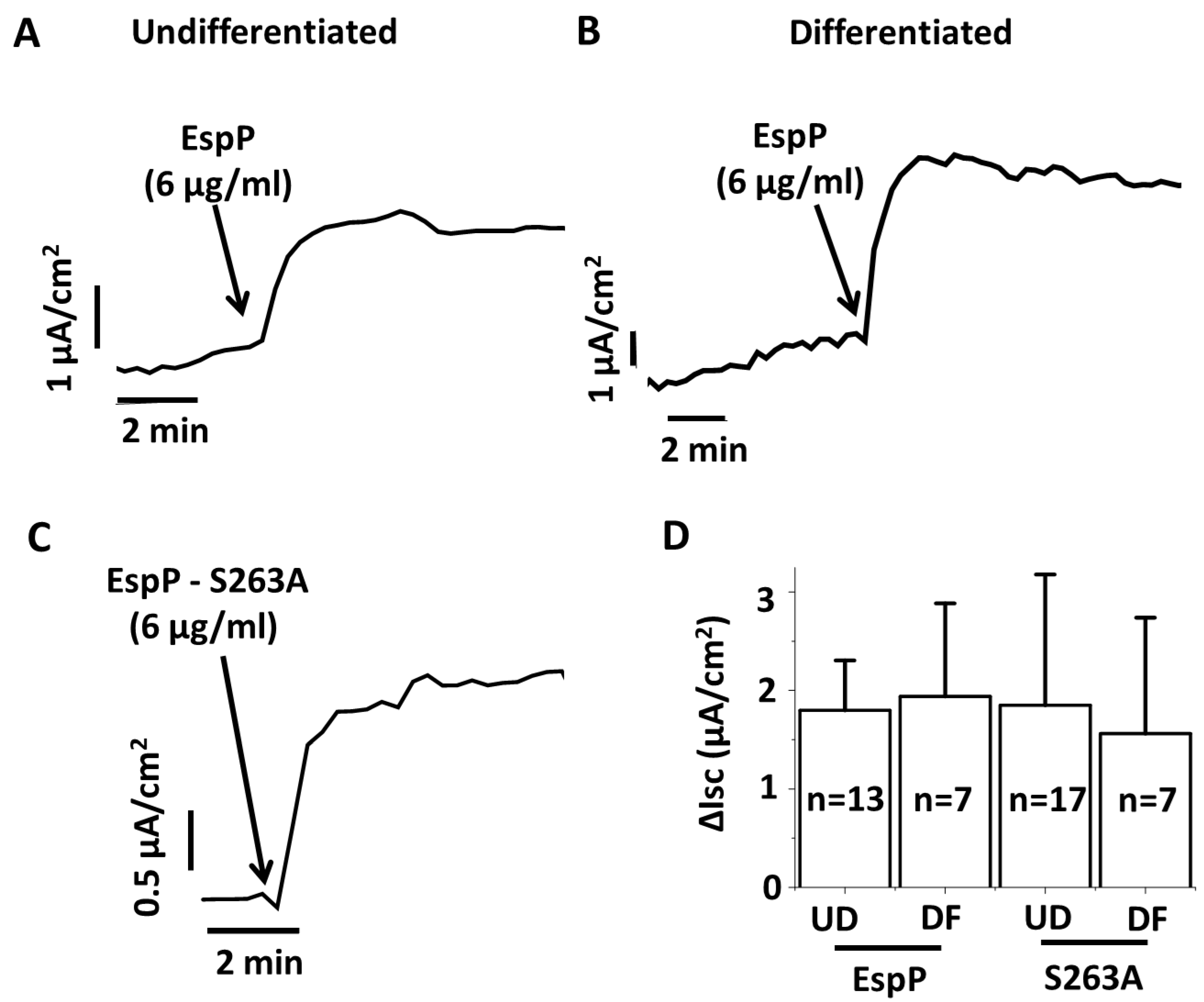
2.1. EspP Demonstrates Enterotoxic Activity
2.2. EspP Enterotoxic Activity is Independent of its Serine Protease Activity
2.3. EspP Specifically Alters Isc in a Concentration-Dependent Manner
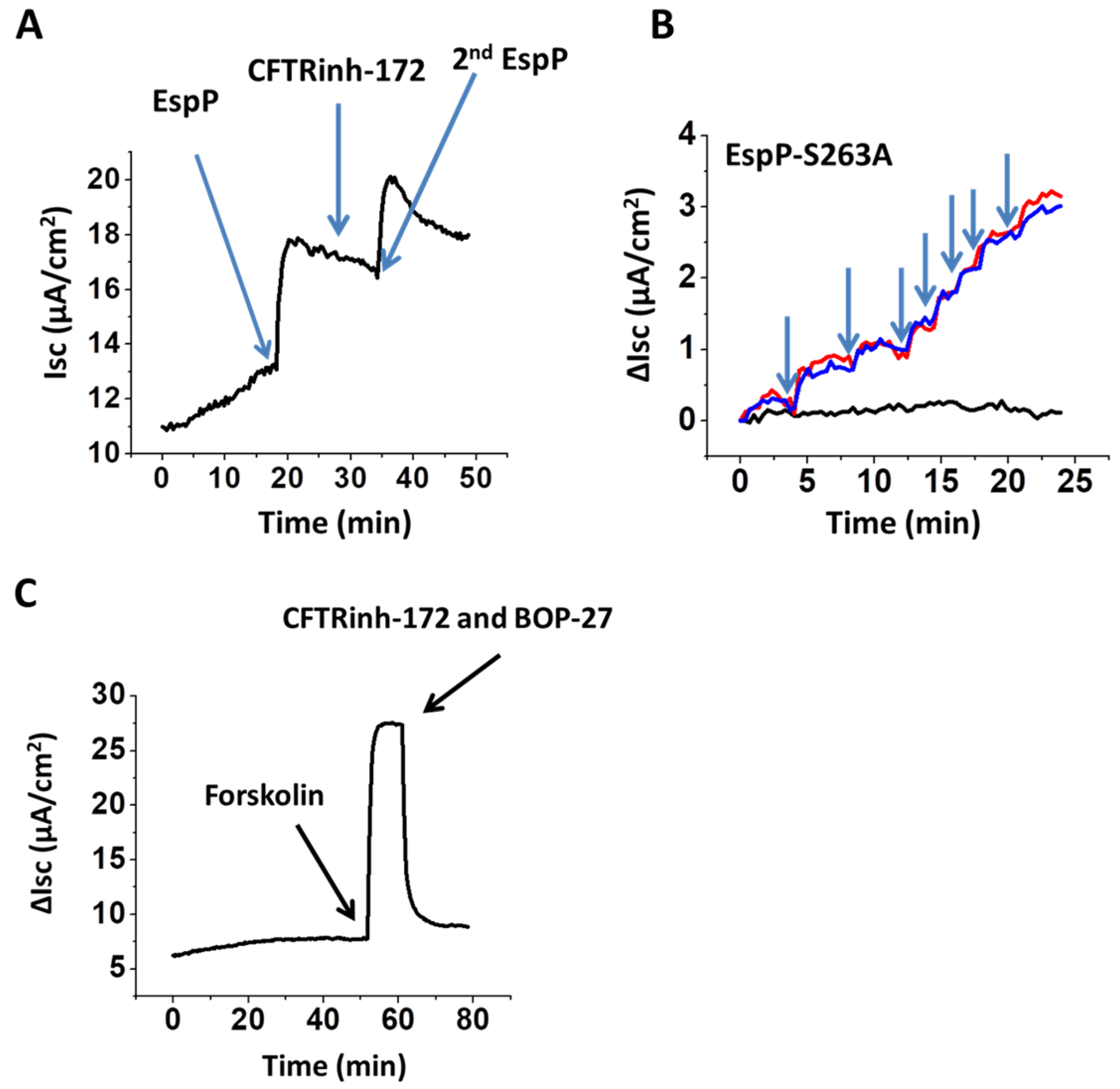
2.4. EspP-Stimulated Current is CFTR-Independent
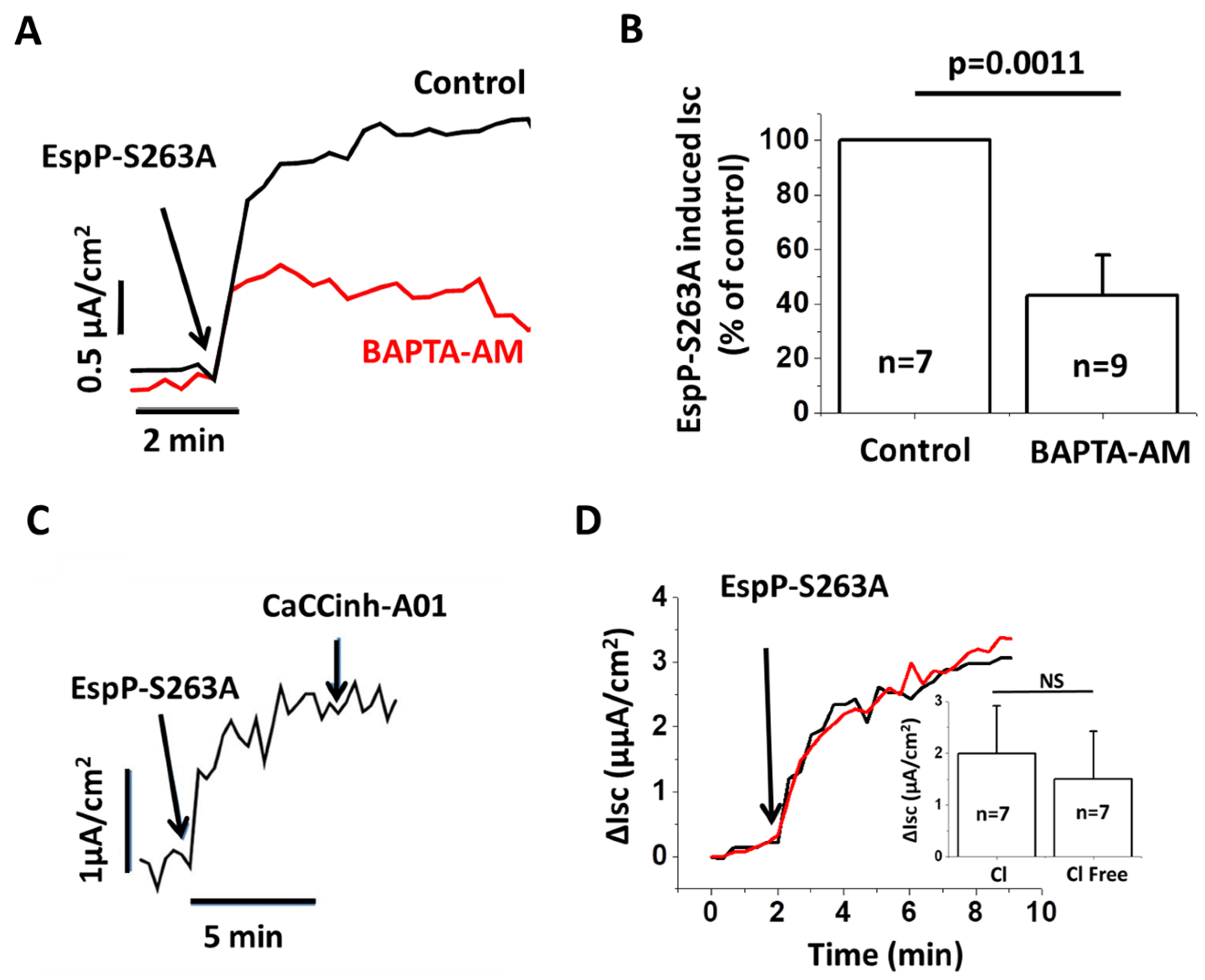
2.5. EspP-Induced Isc is Partially Ca2+ Dependent
2.6. Effects of Class-2 SPATEs on Ion Secretion by Human Intestinal Epithelial Cultures
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents
5.2. Purification of EspP and EspP S263A
5.3. Purification of Pic and SepA
5.4. Human Colonoid and Enteroid Cultures
5.5. Short-Circuit Current Measurements with the Ussing Chamber/Voltage Clamp Technique
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, V.K.; Hodges, K.; Hecht, G. Enteric infection meets intestinal function: How bacterial pathogens cause diarrhoea. Nat. Rev. Microbiol. 2009, 7, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.E.; Elliott, E.J. Interventions for preventing diarrhea-associated hemolytic uremic syndrome: Systemic review. BMC Public Health 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bell, C.; Buret, A.; Robins-Browne, R.; Stiel, D.; O’Loughlin, E. The effect of enterohemorrhagic Escherichia coli 0157:H7 on intestinal structure and solute transport in rabbits. Gastroenterology 1993, 104, 467–474. [Google Scholar] [CrossRef]
- Karmali, M.A.; Griffin, P.M. Emerging public health challenges of Shiga toxin-producing Escherichia coli related to changes in the pathogen, the population, and the environment. Clin. Infect. Dis. 2017, 64, 371–376. [Google Scholar] [PubMed]
- Mellies, J.L.; Navarro-Garcia, F.; Okeke, I.; Frederickson, J.; Nataro, J.P.; Kaper, J.B. espC pathogenicity island of enteropathogenic Escherichia coli encodes an enterotoxin. Infect. Immun. 2001, 69, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Guignot, J.; Segura, A.; Tran Van Nhieu, G. The serine protease EspC from enteropathogenic Escherichia coli regulates pore formation and cytotoxicity mediated by the type III secretion system. PLoS Pathog. 2015, 11, e1005013. [Google Scholar] [CrossRef] [PubMed]
- Dautin, N. Serine protease autotransporters of enterobacteriaceae (SPATEs): Biogenesis and function. Toxins 2010, 2, 1179–1206. [Google Scholar] [CrossRef] [PubMed]
- In, J.; Foulke-Abel, J.; Zachos, N.C.; Hansen, A.-M.; Kaper, J.B.; Bernstein, H.D.; Halushka, M.; Blutt, S.; Estes, M.K.; Donowitz, M.; et al. Enterohemorrhagic Escherichia coli reduce mucus and intermicrovillar bridges in human stem cell-derived colonoids. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 48–62. [Google Scholar] [CrossRef] [PubMed]
- In, J.G.; Foulke-Abel, J.; Estes, M.K.; Zachos, N.C.; Kovbasnjuk, O.; Donowitz, M. Human mini-guts: New insights into intestinal physiology and host-pathogen interactions. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.; Baetz, N.W.; Staab, J.F.; Donowitz, M.; Kovbasnjuk, O.; Pasetti, M.F.; Zachos, N.C. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Sci. Rep. 2017, 7, 45270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut 2015, 64, 911. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.; Sperandio, V. Enterohemorrhagic E. coli (EHEC) pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2, 90. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.R.; Sperandio, V. Shiga toxin in enterohemorrhagic E. coli: Regulation and novel anti-virulence strategies. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef]
- Thiagarajah, J.R.; Donowitz, M.; Verkman, A.S. Secretory diarrhoea: Mechanisms and emerging therapies. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Kopic, S.; Geibel, J.P. Toxin mediated diarrhea in the 21st century: The pathophysiology of intestinal ion transport in the course of ETEC, V. cholerae and rotavirus infection. Toxins 2010, 2, 2132–2157. [Google Scholar] [CrossRef] [PubMed]
- Navaneethan, U.; Giannella, R.A. Mechanisms of infectious diarrhea. Nat. Clin. Pract. Gastroenterol. Hepatol. 2008, 5, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, O.; Peterson, J.H.; Ieva, R.; Bernstein, H.D. Mechanistic link between β barrel assembly and the initiation of autotransporter secretion. Proc. Natl. Acad. Sic. USA 2013, 110, E938–E947. [Google Scholar] [CrossRef] [PubMed]
- In, J.; Lukyanenko, V.; Foulke-Abel, J.; Hubbard, A.L.; Delannoy, M.; Hansen, A.M.; Kaper, J.B.; Boisen, N.; Nataro, J.P.; Zhu, C.; et al. Serine protease Espp from enterohemorrhagic Escherichia coli is sufficient to induce Shiga toxin macropinocytosis in intestinal epithelium. PLoS ONE 2013, 8, e69196. [Google Scholar] [CrossRef] [PubMed]
- Brockmeyer, J.; Bielaszewska, M.; Fruth, A.; Bonn, M.L.; Mellmann, A.; Humpf, H.-U.; Karch, H. Subtypes of the plasmid-encoded serine protease EspP in Shiga toxin-producing Escherichia coli: Distribution, secretion, and proteolytic activity. Appl. Environ. Microbiol. 2007, 73, 6351–6359. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Brockmeyer, J. Prevalence, biogenesis, and functionality of the serine protease autotransporter EspP. Toxins 2012, 5, 25–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, A.; Joerss, H.; Brockmeyer, J. Structural and functional characterization of cleavage and inactivation of human serine protease inhibitors by the bacterial SPATE protease EspPα from enterohemorrhagic E. coli. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajah, J.R.; Song, Y.; Haggie, P.M.; Verkman, A.S. A small molecule CFTR inhibitor produces cystic fibrosis-like submucosal gland fluid secretions in normal airways. FASEB J. 2004, 18, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Cil, O.; Phuan, P.-W.; Gillespie, A.M.; Lee, S.; Tradtrantip, L.; Yin, J.; Tse, M.; Zachos, N.C.; Lin, R.; Donowitz, M.; et al. Benzopyrimido-pyrrolo-oxazine-dione CFTR inhibitor (R)-BPO-27 for antisecretory therapy of diarrheas caused by bacterial enterotoxins. FASEB J. 2017, 31, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Tse, C.-M.; Avula, L.R.; Singh, V.; Foulke-Abel, J.; Jonge, H.R.D.; Donowitz, M. Molecular basis and differentiation-associated alterations of anion secretion in human duodenal enteroid monolayers. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 591–609. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Wiegerinck, C.L.; Jonge, H.R.D.; Bronsveld, I.; Janssens, H.M.; Groot, K.M.D.W.; Brandsma, A.M.; Jong, N.W.M.D.; Bijvelds, M.J.C.; Scholte, B.J.; et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Namkung, W.; Phuan, P.W.; Verkman, A.S. TMEM16A inhibitors reveal TMEM16A as a minor component of calcium-activated chloride channel conductance in airway and intestinal epithelial cells. J. Biol. Chem. 2011, 286, 2365–2374. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.R.; Cappello, R.; Navarrogarcía, F.; Nataro, J.P. Functional comparison of serine protease autotransporters of enterobacteriaceae. Infect. Immun. 2002, 70, 7105–7113. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Perez, F.; Nataro, J.P. Bacterial serine proteases secreted by the autotransporter pathway: Classification, specificity, and role in virulence. Cell. Mol. Life Sci. 2013, 71, 745–770. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Lujan, J.L.; Vijayakumar, V.; Gong, M.; Smith, R.; Santiago, A.E.; Ruiz-Perez, F. Broad spectrum activity of a lectin-like bacterial serine protease family on human leukocytes. PLoS ONE 2014, 9, e107920. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.S.; Sorenson, J.; Harris, J.B.; Robins, W.P.; Charles, R.C.; Jean-charles, R.R.; Bullard, J.; Webster, D.R.; Kasarskis, A.; Peluso, P.; et al. The origin of the haitian cholera outbreak strain. N. Engl. J. Med. 2011, 364, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.; Bouhenia, M.; Alyusfi, R.; Alkohlani, A.; Naji, M.; De, X.R.; Radiguès, X.D.; Abubakar, A.M.; Almoalmi, A.; Seguin, C.; et al. Cholera epidemic in Yemen, 2016–18: An analysis of surveillance data. Lancet Glob. Health 2018, 6, e680–e690. [Google Scholar] [CrossRef]
- Group SWR. Experiences from the Shiga toxin-producing Escherichia coli O104:H4 outbreak in Germany and research needs in the field, Berlin, 28–29 November 2011. Eur. Surveill. 2012, 17. [Google Scholar] [CrossRef]
- Andresen, V.; Löwe, B.; Broicher, W.; Riegel, B.; Fraedrich, K.; Wulffen, M.V.; Gappmayer, K.; Wegscheider, K.; Treszl, A.; Rose, M.; et al. Post-infectious irritable bowel syndrome (PI-IBS) after infection with shiga-like toxin-producing Escherichia coli (STEC) O104:H4: A cohort study with prospective follow-up. United Eur. Gastroenterol. J. 2016, 4, 121. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chem. 2012, 56, 3277–3282. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Christie, D.L. Antimotility agents and E. coli infection. CMAJ 1999, 160, 984–986. [Google Scholar] [PubMed]
- Tarr, P.I.; Watkins, S.L.; Neill, M.A. Risk of hemolytic uremic syndrome from antibiotic treatment of Escherichia coli O157:H7 colitis. JAMA 2002, 288, 3111–3112. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.K.; Mckee, R.; Schnadower, D.; Tarr, P.I. Treatment of Shiga toxin-producing Escherichia coli infections. Infect. Dis Clin. N. Am. 2013, 27, 577–597. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.M.; Griffin, P.M.; Jones, T.F.; Smith, K.E.; Scallan, E. Antimicrobial and antimotility agent use in persons with Shiga toxin–producing Escherichia coli O157 infection in FoodNet sites. Clin. Infect. Dis. 2011, 52, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Zachos, N.C.; Kovbasnjuk, O.; Foulkeabel, J.; In, J.; Blutt, S.E.; Jonge, H.R.D.; Estes, M.K.; Donowitz, M. Human enteroids/colonoids and intestinal organoids functionally recapitulate normal intestinal physiology and pathophysiology. J. Biol. Chem. 2015, 291, 3759–3766. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.Y.; Blutt, S.E.; Crawford, S.E.; Ettayebi, K.; Zeng, X.L.; Saxena, K.; Ramani, S.; Karandikar, U.C.; Zachos, N.C.; Estes, M.K. Human intestinal enteroids: New models to study gastrointestinal virus infections. Methods Mol. Biol. 2017, 1–19. [Google Scholar] [CrossRef]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.J.; Es, J.H.V.; Brink, S.V.D.; Houdt, W.J.V.; Pronk, A.; Gorp, J.V.; Siersema, P.D.; et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.-S.; Iliopoulos, D.; et al. Origins of the e. coli strain causing an outbreak of hemolytic-uremic syndrome in germany. N. Engl. J. Med. 2016, 187, 709–717. [Google Scholar]
- Scheutz, F.; Nielsen, E.M.; Frimodtmøller, J.; Boisen, N.; Morabito, S.; Tozzoli, R.; Nataro, J.P.; Caprioli, A. Characteristics of the enteroaggregative Shiga toxin/verotoxin-producing Escherichia coli O104:H4 strain causing the outbreak of haemolytic uraemic syndrome in Germany, May to June 2011. Eurosurveillance 2011, 16, 46–64. [Google Scholar] [CrossRef]
- Navarro-Garcia, F.; Eslava, C.J.; Lopez-Revilla, R.; Czeczulin, J.; Srinivas, S.; Nataro, J.; Cravioto, A. In vitro effects of a high-molecular-weight heat-labile enterotoxin from enteroaggregative Escherichia coli. Infect. Immun. 1998, 66, 3149–3154. [Google Scholar] [PubMed]
- Munera, D.; Ritchie, J.M.; Hatzios, S.K.; Bronson, R.; Fang, G.; Schadt, E.E.; Davis, B.M.; Waldor, M.K. Autotransporters but not pAA are critical for rabbit colonization by Shiga toxin-producing Escherichia coli O104:H4. Nat. Commun. 2014, 5, 3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.B.; Larocque, R.C.; Qadri, F.; Ryan, E.T.; Calderwood, S.B. Cholera. Lancet 2012, 379, 2466–2476. [Google Scholar] [CrossRef] [Green Version]
- Grisaru, S.; Xie, J.; Samuel, S.; Hartling, L.; Tarr, P.I.; Schnadower, D.; Freedman, S.B.; for the Alberta Provincial Pediatric Enteric Infection Team. Associations between hydration status, intravenous fluid administration, and outcomes of patients infected with Shiga toxin-producing Escherichia coli: A systematic review and meta-analysis. JAMA Pediatr. 2017, 171, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Szabady, R.L.; Peterson, J.H.; Skillman, K.M.; Bernstein, H.D. An unusual signal peptide facilitates late steps in the biogenesis of a bacterial autotransporter. Proc. Natl. Acad. Sic. USA 2005, 102, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Brunder, W.; Schmidt, H.; Karch, H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157:H7 cleaves human coagulation factor V. Mol. Microbiol. 1997, 24, 767–778. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tse, C.M.; In, J.G.; Yin, J.; Donowitz, M.; Doucet, M.; Foulke-Abel, J.; Ruiz-Perez, F.; Nataro, J.P.; Zachos, N.C.; Kaper, J.B.; et al. Enterohemorrhagic E. coli (EHEC)—Secreted Serine Protease EspP Stimulates Electrogenic Ion Transport in Human Colonoid Monolayers. Toxins 2018, 10, 351. https://doi.org/10.3390/toxins10090351
Tse CM, In JG, Yin J, Donowitz M, Doucet M, Foulke-Abel J, Ruiz-Perez F, Nataro JP, Zachos NC, Kaper JB, et al. Enterohemorrhagic E. coli (EHEC)—Secreted Serine Protease EspP Stimulates Electrogenic Ion Transport in Human Colonoid Monolayers. Toxins. 2018; 10(9):351. https://doi.org/10.3390/toxins10090351
Chicago/Turabian StyleTse, C. Ming, Julie G. In, Jianyi Yin, Mark Donowitz, Michele Doucet, Jennifer Foulke-Abel, Fernando Ruiz-Perez, James P. Nataro, Nicholas C. Zachos, James B. Kaper, and et al. 2018. "Enterohemorrhagic E. coli (EHEC)—Secreted Serine Protease EspP Stimulates Electrogenic Ion Transport in Human Colonoid Monolayers" Toxins 10, no. 9: 351. https://doi.org/10.3390/toxins10090351
APA StyleTse, C. M., In, J. G., Yin, J., Donowitz, M., Doucet, M., Foulke-Abel, J., Ruiz-Perez, F., Nataro, J. P., Zachos, N. C., Kaper, J. B., & Kovbasnjuk, O. (2018). Enterohemorrhagic E. coli (EHEC)—Secreted Serine Protease EspP Stimulates Electrogenic Ion Transport in Human Colonoid Monolayers. Toxins, 10(9), 351. https://doi.org/10.3390/toxins10090351