Venomous Arachnid Diagnostic Assays, Lessons from Past Attempts

Abstract
:1. Introduction
2. In Vitro Assays for Identification and/or Quantification of Arachnid Venoms
2.1. Spiders
2.2. Scorpions
3. In Vitro Assays for the Establishment of Antivenom Effectiveness
3.1. Spiders
3.2. Scorpions
4. Biotechnological Limitations and Perspectives for Arachnid Venom Detection
5. Unmet Needs for Arachnid Diagnosis and Recommendations to Validate an Efficient Test
6. Materials and Methods
Funding
Conflicts of Interest
References
- Cordeiro, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Arantes, E.C. Arachnids of medical importance in Brazil: Main active compounds present in scorpion and spider venoms and tick saliva. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 24. [Google Scholar] [CrossRef] [PubMed]
- Junghanss, T.; Bodio, M. Medically Important Venomous Animals: Biology, Prevention, First Aid, and Clinical Management. Clin. Infect. Dis. 2006, 43, 1309–1317. [Google Scholar] [PubMed] [Green Version]
- Garb, J.E.; González, A.; Gillespie, R.G. The black widow spider genus Latrodectus (Araneae: Theridiidae): Phylogeny, biogeography, and invasion history. Mol. Phylogenet. Evol. 2004, 31, 1127–1242. [Google Scholar] [CrossRef] [PubMed]
- Gremski, L.H.; Trevisan-Silva, D.; Ferrer, V.P.; Matsubara, F.H.; Meissner, G.O.; Wille, A.C.M.; Vuitika, L.; Dias-Lopes, C.; Ullah, A.; de Moraes, F.R.; et al. Recent advances in the understanding of brown spider venoms: From the biology of spiders to the molecular mechanisms of toxins. Toxicon 2014, 83, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Vetter, R.S.; Isbister, G.K. Medical aspects of spider bites. Annu. Rev. Entomol. 2008, 53, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Isbister, G.K.; Fan, H.W. Spider bite. Lancet 2011, 378, 2039–2047. [Google Scholar] [CrossRef]
- Lucas, S.M. The history of venomous spider identification, venom extraction methods and antivenom production: A long journey at the Butantan Institute, São Paulo, Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Bawaskar, H.S.; Bawaskar, P.H. Scorpion sting. J. Assoc. Phys. India 2012, 60, 46–55. [Google Scholar] [CrossRef]
- Isbister, G.K.; Bawaskar, H.S. Scorpion Envenomation. N. Engl. J. Med. 2014, 371, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.V.; Theodorou, A.A.; Berg, R.A.; Mallie, J.; Chávez-Méndez, A.; García-Ubbelohde, W.; Hardiman, S.; Alagón, A. Antivenom for Critically Ill Children with Neurotoxicity from Scorpion Stings. N. Engl. J. Med. 2009, 360, 2090–2098. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P. Emerging options for the management of scorpion stings. Drug Des. Dev. Ther. 2012, 6, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Gaver-Wainwright, M.M.; Zack, R.S.; Foradori, M.J.; Lavine, L.C. Misdiagnosis of Spider Bites: Bacterial Associates, Mechanical Pathogen Transfer, and Hemolytic Potential of Venom From the Hobo Spider, Tegenaria agrestis (Araneae: Agelenidae). J. Med. Entomol. 2011, 48, 382–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowry, J.B.; Spyker, D.A.; Cantilena, L.R.; McMillan, N.; Ford, M. 2013 Annual Report of the American Association of Poison Control Centers’ National Poison Data System (NPDS): 31st Annual Report. Clin. Toxicol. 2014, 52, 1032–1283. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.G.; Vetter, R.S. An approach to spider bites. Erroneous attribution of dermonecrotic lesions to brown recluse or hobo spider bites in Canada. Can. Fam. Phys. 2004, 50, 1098–1101. [Google Scholar]
- Vetter, R.S. Arachnids misidentified as brown recluse spiders by medical personnel and other authorities in North America. Toxicon 2009, 54, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Vetter, R.S.; Cushing, P.E.; Crawford, R.L.; Royce, L.A. Diagnoses of brown recluse spider bites (loxoscelism) greatly outnumber actual verifications of the spider in four western American states. Toxicon 2003, 42, 413–418. [Google Scholar] [CrossRef]
- Swanson, D.L.; Vetter, R.S. Bites of brown recluse spiders and suspected necrotic arachnidism. N. Engl. J. Med. 2005, 352, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Rezende, N.A.D.E.; Chavez-Olortegui, C.; Amaral, C.F.S. Is the severity of Tityus serrulatus scorpion envenoming related to plasma venom concentrations? Toxicon 1996, 34, 820–823. [Google Scholar] [CrossRef]
- Rezende, N.A.; de Dias, M.B.; Campolina, D.; Chavez-Olortegui, C.; Amaral, C.F.S. Standardization of an enzyme linked immunosorbent assay (ELISA) for detecting circulating toxic venom antigens in patients stung by the scorpion Tityus serrulatus. Rev. Inst. Med. Trop. São Paulo 1995, 37, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi-triki, D.; Ferquel, E.; Robbe-vincent, A.; Bon, C.; Choumet, V.; Laraba-djebari, F. Epidemiological data, clinical admission gradation and biological quantification by ELISA of scorpion envenomations in Algeria: Effect of immunotherapy. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 240–250. [Google Scholar] [CrossRef]
- Krifi, M.N.; Kharrat, H.; Zghal, K.; Abdouli, M.; Abroug, F.; Bouchoucha, S.; Dellagi, K.; El Ayeb, M. Development of an ELISA for the detection of scorpion venoms in sera of humans envenomed by Androctonus australis garzonii (AAG) and Buthus occitanus tunetanus (BOT): Correlation with clinical severity of envenoming in Tunisia. Toxicon 1998, 36, 887–900. [Google Scholar] [CrossRef]
- McGlasson, D.L.; Green, J.A.; Stoecker, W.V.; Babcock, J.L.; Calcara, D.A. Duration of Loxosceles reclusa venom detection by ELISA from swabs. Clin. Lab. Sci. 2009, 22, 216–222. [Google Scholar] [PubMed]
- Barrett, S.M.; Romine-Jenkins, M.; Blick, K.E. Passive hemagglutination inhibition test for diagnosis of brown recluse spider bite envenomation. Clin. Chem. 1993, 39, 2104–2107. [Google Scholar] [CrossRef]
- Stoecker, W.V.; Green, J.A.; Gomez, H.F. Diagnosis of loxoscelism in a child confirmed with an enzyme-linked immunosorbent assay and noninvasive tissue sampling. J. Am. Acad. Dermatol. 2006, 55, 888–890. [Google Scholar] [CrossRef] [PubMed]
- Keklikci, U.; Akdeniz, S.; Sakalar, Y.B.; Cakmak, S.S.; Unlu, K. Loxosceles reclusa bite to the eyelid. Eur. J. Ophthalmol. 2008, 18, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Pauli, I.; Minozzo, J.C.; Henrique, D.A.; Silva, P.; Chaim, O.M.; Veiga, S.S. Analysis of therapeutic benefits of antivenin at different time intervals after experimental envenomation in rabbits by venom of the brown spider (Loxosceles intermedia). Toxicon 2009, 53, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Gomez, H.F.; Greenfield, D.M.; Miller, M.J.; Warren, J.S. Direct correlation between diffusion of Loxosceles reclusa venom and extent of dermal inflammation. Acad. Emerg. Med. 2001, 8, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Krywko, D.M.; Gomez, H.F. Detection of Loxosceles species venom in dermal lesions: A comparison of 4 venom recovery methods. Ann. Emerg. Med. 2002, 39, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.J.; Gomez, H.F.; Snider, R.J.; Stephens, E.L.; Czop, R.M.; Warren, J.S. Detection of Loxosceles venom in lesional hair shafts and skin: Application of a specific immunoassay to identify dermonecrotic arachnidism. Am. J. Emerg. Med. 2000, 18, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Olórtegui, C.; Zanetti, V.C.; Ferreira, A.P.; Minozzo, J.C.; Mangili, O.C.; Gubert, I.C. ELISA for the detection of venom antigens in experimental and clinical envenoming by Loxosceles intermedia spiders. Toxicon 1998, 36, 563–569. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Cardoso, J.L.C.; Eickstedt, V.R.D.; Mota, I. IgG antibodies to Loxosceles sp. spider venom in human envenoming. Toxicon 1992, 30, 1117–1121. [Google Scholar] [CrossRef]
- Lucas, S. Spiders in Brazil. Toxicon 1988, 26, 759–772. [Google Scholar] [CrossRef]
- Miller, M.; Leary, M.A.O.; Isbister, G.K. Towards rationalisation of antivenom use in funnel-web spider envenoming: Enzyme immunoassays for venom concentrations. Clin. Toxicol. 2016, 54, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Olórtegui, C.; Bohórquez, K.; Alvarenga, L.M.; Kalapothakis, E.; Campolina, D.; Maria, W.S.; Diniz, C.R. Sandwich-ELISA detection of venom antigens in envenoming by Phoneutria nigriventer spider. Toxicon 2001, 39, 909–911. [Google Scholar] [CrossRef]
- Bucaretchi, F.; Mello, S.M.; Vieira, R.J.; Mamoni, R.L.; Blotta, M.H.; Antunes, E.; Hyslop, S. Systemic envenomation caused by the wandering spider Phoneutria nigriventer, with quantification of circulating venom. Clin. Toxicol. 2008, 46, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Olórtegui, C.; Fonseca, S.C.; Campolina, D.; Amaral, C.F.; Diniz, C.R. ELISA for the detection of toxic antigens in experimental and clinical envenoming by Tityus serrulatus scorpion venom. Toxicon 1994, 32, 1649–1656. [Google Scholar] [CrossRef]
- Ghalim, N.; El-hafny, B.; Sebti, F.; Heikel, J.; Lazar, N.; Moustanir, R.; Benslimane, A. Scorpion envenomation and serotherapy in Morocco. Am. J. Trop. Med. 2000, 62, 277–283. [Google Scholar] [CrossRef]
- D’Suze, G.; Moncada, S.; Gonzales, C.; Sevcik, C.; Aguilar, V.; Alagon, A. Relationship between plasmatic levels of various cytokines, tumour necrosis factor, enzymes, glucose and venom concentration following Tityus scorpion sting. Toxicon 2003, 41, 367–375. [Google Scholar]
- Osnaya-Romero, N.; Acosta-Saavedra, L.C.; Goytia-Acevedo, R.; Lares-Asseff, I.; Basurto-Celaya, G.; Perez-Guille, G.; Possani, L.D.; Calderón-Aranda, E.S. Serum level of scorpion toxins, electrolytes and electrocardiogram alterations in Mexican children envenomed by scorpion sting. Toxicon 2016, 122, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Chase, P.; Boyer-Hassen, L.; Mcnally, J.; Vazquez, H.L.; Theodorou, A.A.; Walter, F.G.; Alagon, A. Serum levels and urine detection of Centruroides sculpturatus venom in significantly envenomated patients. Clin. Toxicol. 2009, 47, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Krifi, M.N.; Savin, S.; Debray, M.; Bon, C.; El Ayeb, M.; Choumet, V. Pharmacokinetic studies of scorpion venom before and after antivenom immunotherapy. Toxicon 2005, 45, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Khoobdel, M.; Nikbakhtboroujeni, G.; Zahraeisalehi, T.; Khosravi, M.; Sasani, F.; Bokaei, S.; Koochakzadeh, A.; Zamani-Ahmadmahmudi, M.; Akbari, A. Diagnosis of Mesobuthus eupeus envenomation by skin test: Reverse passive Arthus reaction. Toxicon 2014, 77, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Krifi, M.N.; Amri, F.; Kharrat, H.; El Ayeb, M. Evaluation of antivenom therapy in children severely envenomed by Androctonus australis garzonii (Aag) and Buthus occitanus tunetanus (Bot) scorpions. Toxicon 1999, 37, 1627–1634. [Google Scholar] [CrossRef]
- Toro, A.F.; Malta, M.B.; Soares, S.L.; Rocha, G.C.; da Lira, M.; De Oliveira, T.A.; Takehara, H.A.; Lopes-Ferreira, M.; Santoro, M.L.; Guidolin, R.; et al. Role of IgG (T) and IgGa isotypes obtained from arachnidic antivenom to neutralize toxic activities of Loxosceles gaucho, Phoneutria nigriventer and Tityus serrulatus venoms. Toxicon 2006, 48, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Isbister, G.K.; Gray, M.R.; Balit, C.R.; Raven, R.J.; Stokes, B.J.; Porges, K.; Tankel, A.S.; Turner, E.; White, J.; Fisher, M.M. Funnel-web spider bite: A Systematic Review of Recorded Clinical Cases. Med. J. Aust. 2005, 182, 407–411. [Google Scholar] [PubMed]
- Krifi, M.N.; Miled, K.; Abderrazek, M.; El Ayeb, M. Effects of antivenom on Buthus occitanus tunetanus (Bot) scorpion venom pharmacokinetics: Towards an optimization of antivenom immunotherapy in a rabbit model. Toxicon 2001, 39, 1317–1326. [Google Scholar] [CrossRef]
- Revelo, M.P.; Bambirra, E.A.; Ferreira, A.P.; Diniz, C.R.; Chavez-Olortegui, C. Body distribution of Tityus serrulatus scorpion venom in mice and effects of scorpion antivenom. Toxicon 1996, 34, 1119–1125. [Google Scholar] [CrossRef]
- Rezende, N.A.; Amaral, C.F.; Freire-maia, L. Immunotherapy for scorpion envenoming in Brazil. Toxicon 1998, 36, 1507–1513. [Google Scholar] [CrossRef]
- Venancio, E.J.; Portaro, F.C.V.; Kuniyoshi, A.K.; Carvalho, D.C.; Pidde-Queiroz, G.; Tambourgi, D.V. Enzymatic properties of venoms from Brazilian scorpions of Tityus genus and the neutralisation potential of therapeutical antivenoms. Toxicon 2013, 69, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Santana, G.C.; Freire, A.C.T.; Ferreira, A.P.L.; Chavez-Olortegui, C.; Diniz, C.R.; Freire-Maia, L. Pharmacokinetics of Tityus serrulatus scorpion venom determined by enzyme-linked immunosorbent assay in the rat. Toxicon 1996, 34, 1063–1066. [Google Scholar] [CrossRef]
- Ismail, M. The scorpion envenomation syndrome. Toxicon 1995, 33, 825–858. [Google Scholar] [CrossRef]
- Michnick, S.W.; Sidhu, S.S. Submitting antibodies to binding arbitration. Nat. Chem. Biol. 2008, 4, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.M.; Avila, R.A.M.; De Amim, P.R.; Martins, M.S. Molecular characterization of a neutralizing murine monoclonal antibody against Tityus serrulatus scorpion venom. Toxicon 2005, 46, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.M.; Martins, M.S.; Moura, J.F.; Oliveira, C.; Mangili, O.C.; Granier, C.; Chávez-Olórtegui, C. Production of monoclonal antibodies capable of neutralizing dermonecrotic activity of Loxosceles intermedia spider venom and their use in a specific immunometric assay. Toxicon 2003, 42, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Clot-faybesse, O.; Juin, M.; Roche, H.; Devaux, C. Monoclonal antibodies against the Androctonus australis hector scorpion neurotoxin I: Characterisation and use for venom neutralisation. FEBS Lett. 1999, 458, 313–318. [Google Scholar] [CrossRef]
- Devaux, C.; Clot-faybesse, O.; Juin, M.; Mabrouk, K.; Sabatier, J.-M.; Rochat, H. Monoclonal antibodies neutralizing the toxin II from Androctonus australis hector scorpion venom: Usefulness of a synthetic, non-toxic analog. FEBS Lett. 1997, 412, 456–460. [Google Scholar] [CrossRef]
- Dias-Lopes, C.; Felicori, L.; Rubrecht, L.; Cobo, S.; Molina, L.; Nguyen, C.; Galéa, P.; Granier, C.; Molina, F.; Chávez-Olortegui, C. Generation and molecular characterization of a monoclonal antibody reactive with conserved epitope in sphingomyelinases D from Loxosceles spider venoms. Vaccine 2014, 32, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, P.; Fernandes, I.; Barbaro, K.C. Neutralization of dermonecrotic and lethal activities and differences among 32–35 kDa toxins of medically important Loxosceles spider venoms in Brazil revealed by monoclonal antibodies. Toxicon 2001, 39, 1333–1342. [Google Scholar] [CrossRef]
- Jiacomini, I.; Silva, S.K.; Aubrey, N.; Muzard, J.; Chavez-olortegui, C.; Moura, J.; De Moura, J.; Billiald, P.; Alvarenga, L.M. Immunodetection of the “brown” spider (Loxosceles intermedia) dermonecrotoxin with an scFv-alkaline phosphatase fusion protein. Immunol. Lett. 2016, 173, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Zoccal, K.F.; Roncolato, E.C.; Bertolini, T.B.; Campos, L.B.; Cologna, C.T.; Faccioli, L.H.; Arantes, E.C.; Barbosa, J.E. Serrumab: A human monoclonal antibody that counters the biochemical and immunological effects of Tityus serrulatus venom. J. Immunotoxicol. 2012, 9, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauridsen, L.H.; Shamaileh, H.A.; Edwards, S.L.; Taran, E.; Veedu, R.N. Rapid one-step selection method for generating nucleic acid aptamers: Development of a DNA Aptamer against α-bungarotoxin. PLoS ONE 2012, 7, e41702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Zheng, Y.; Wang, X.; Tan, X.; Zhang, T.; Xin, W.; Wang, J.; Huang, Y.; Fan, Q.; Wang, J. Recognition of Bungarus multicinctus Venom by a DNA Aptamer against β-Bungarotoxin. PLoS ONE 2014, 9, e105404. [Google Scholar] [CrossRef] [PubMed]
- Sapag, A.; Salinas-luypaert, C.; Constenla-muñoz, C. First report of in vitro selection of RNA aptamers targeted to recombinant Loxosceles laeta spider toxins. Biol. Res. 2014, 47, 2. [Google Scholar] [PubMed]
- Zhang, S.; D’Angeli, A.G.; Brennan, J.P.; Huo, Q. Predicting detection limits of enzyme-linked immunosorbent assay (ELISA) and bioanalytical techniques in general. Analyst 2014, 139, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Drain, P.K.; Hyle, E.P.; Noubary, F.; Freedberg, K.A.; Wilson, D.; Bishai, W.R.; Rodriguez, W.; Bassett, I.V. Diagnostic point-of-care tests in resource-limited settings. Lancet Infect. Dis. 2014, 14, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Tambourgi, D.V.; De, F.; Fernandes Pedrosa, M.; van den Berg, C.W.; Goncalves-de-Andrade, R.M.; Ferracini, M.; Paixao-Cavalcante, D.; Morgan, B.P.; Rushmere, N.K. Molecular cloning, expression, function and immunoreactivities of members of a gene family of sphingomyelinases from Loxosceles venom glands. Mol. Immunol. 2004, 41, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Tighe, P.J.; Ryder, R.R.; Todd, I.; Fairclough, L.C. ELISA in the multiplex era: Potentials and pitfalls. Proteom.-Clin. Appl. 2015, 9, 406–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vial, S.; Wenger, J. Single-step homogeneous immunoassay for detecting prostate-specific antigen using dual-color light scattering of metal nanoparticles. Analyst 2017, 142, 3484–3491. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, J.A.; Matas, L.; Manterola, J.M.; Blavia, R.; Sopena, N.; Belda, F.J.; Padilla, E.; Giménez, M.; Sabrià, M.; Morera, J.; et al. Comparison of Radioimmunoassay and Enzyme Immunoassay Kits for Detection of Legionella pneumophila Serogroup 1 Antigen in Both Concentrated and Nonconcentrated Urine Samples. J. Clin. Microbiol. 1997, 35, 1627–1629. [Google Scholar] [PubMed]
- Suntrarachun, S.; Pakmanee, N.; Tirawatnapong, T.; Chanhome, L.; Sitprija, V. Development of a polymerase chain reaction to distinguish monocellate cobra (Naja khouthia) bites from other common Thai snake species, using both venom extracts and bite-site swabs. Toxicon 2001, 39, 1087–1090. [Google Scholar] [CrossRef]
- Sano, T.; Smith, C.; Cantor, C. Immuno-PCR: Very sensitive antigen detection by means of specific antibody-DNA conjugates. Science 1992, 258, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Fehlbaum-Beurdeley, P.; Sol, O.; Désiré, L.; Touchon, J.; Dantoine, T.; Vercelletto, M.; Gabelle, A.; Jarrige, A.C.; Haddad, R.; Lemarié, J.C.; et al. Validation of AclarusDxTM, a Blood-Based Transcriptomic Signature for the Diagnosis of Alzheimer’s Disease. J. Alzheimers Dis. 2012, 32, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Zadel, M.; Maver, A.; Kovanda, A.; Peterlin, B. Transcriptomic Biomarkers for Huntington’ s Disease: Are gene expression signatures in whole blood reliable biomarkers? OMICS 2018, 22, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Darboe, F.; Mbandi, S.K.; Thompson, E.G.; Fisher, M.; Rodo, M.; Rooyen, M.; van Rooyen, M.; Filander, E.; Bilek, N.; Mabwe, S.; et al. Diagnostic performance of an optimized transcriptomic signature of risk of tuberculosis in cryopreserved peripheral blood mononuclear cells. Tuberculosis 2018, 108, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Bochukova, E.G.; Lawler, K.; Croizier, S.; Keogh, J.M.; Patel, N.; Strohbehn, G.; Lo, K.K.; Humphrey, J.; Hokken-Koelega, A.; Damen, L.; et al. A Transcriptomic Signature of the Hypothalamic Response to Fasting and BDNF Deficiency in Prader-Willi Syndrome. Cell Rep. 2018, 22, 3401–3408. [Google Scholar] [CrossRef] [PubMed]
- Nikolayeva, I.; Bost, P.; Casademont, I.; Duong, V.; Koeth, F.; Prot, M.; Czerwinska, U.; Ly, S.; Bleakley, K.; Cantaert, T.; et al. A blood RNA signature detecting severe disease in young dengue patients at hospital arrival. J. Infect. Dis. 2018, 217, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Corzo, G.; Espino-Solis, G.P. Selected scorpion toxin exposures induce cytokine release in human peripheral blood mononuclear cells. Toxicon 2017, 127, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Gehrie, E.A.; Nian, H.; Young, P.P. Brown Recluse spider bite mediated hemolysis: Clinical features, a possible role for complement inhibitor therapy, and reduced RBC surface glycophorin A as a potential biomarker of venom exposure. PLoS ONE 2013, 8, e76558. [Google Scholar] [CrossRef] [PubMed]
- Dantas, R.T.; Sampaio, T.L.; Lima, D.B.; Menezes, B.; Canuto, J.A.; Toyama, M.H.; Evangelista, M.; Martins, C. Evaluation of KIM-1 as an early biomarker of snakebite-induced AKI in mice. Toxicon 2018, 151, 24–28. [Google Scholar] [CrossRef] [PubMed]
- McCall, L.; Morton, J.T.; Bernatchez, J.A.; Jair, L.; Knight, R.; Dorrestein, P.C.; McKerrow, J.H. Mass Spectrometry-Based Chemical Cartography of a Cardiac Parasitic Infection. Anal. Chem. 2017, 89, 10414–10421. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Su, H.; Wu, W.; Wu, D.; Hou, M.; Kuo, C.; Shiea, J. Biomarker Characterization by MALDI–TOF/MS. Adv. Clin. Chem. 2015, 2015. 69, 209–254. [Google Scholar]
- Pham, T.V.; Piersma, S.R.; Oudgenoeg, G.; Jimenez, C.R. Label-free mass spectrometry- based proteomics for biomarker discovery and validation. Expert Rev. 2012, 12, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Celis, F.; Cid-Barrio, L.; Ruiz Encinar, J.; Sanz-Medel, A.; Calvete, J.J. Absolute venomics: Absolute quantification of intact venom proteins through elemental mass spectrometry. J. Proteom. 2017, 164, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Nedelkov, D. Mass spectrometry-based protein assays for in vitro diagnostic testing. Expert Rev. 2012, 12, 235–239. [Google Scholar]
- Mabey, D.; Peeling, R.W.; Ustianowski, A.; Perkins, M.D. Diagnostics for the developing world. Nat. Rev. Microbiol. 2004, 2, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P. Epidemiology of envenomations by terrestrial venomous animals in Brazil based on case reporting: From obvious facts to contingencies. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Gan, S.D.; Patel, K.R. Enzyme immunoassay and enzyme-linked immunosorbent assay. J. Inve. Dermatol. 2013, 133, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Bossuyt, P.M.; Reitsma, J.B.; Bruns, D.E.; Gatsonis, C.A.; Glasziou, P.P.; Irwig, L.; Lijmer, J.G.; Moher, D.; Rennie, D.; De Vet, H.C.; et al. STARD 2015: An updated list of essential items for reporting diagnostic accuracy studies. Clin. Chem. 2015, 61, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B. Clinical Performance Evaluation of Molecular Diagnostic Tests. J. Mol. Diagn. 2016, 18, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Frisoni, G.B.; Boccardi, M.; Barkhof, F.; Blennow, K.; Cappa, S.; Chiotis, K.; Démonet, J.F.; Garibotto, V.; Giannakopoulos, P.; Gietl, A.; et al. Strategic roadmap for an early diagnosis of Alzheimer’s disease based on biomarkers. Lancet Neurol. 2017, 16, 661–676. [Google Scholar] [CrossRef]
- Pepe, M.S.; Etzioni, R.; Feng, Z.; Potter, J.D.; Thompson, M.L.; Thornquist, M.; Winget, M.; Yasui, Y. Phases of Biomarker Development for Early Detection of Cancer. J. Natl. Cancer Inst. 2001, 93, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banoo, S.; Bell, D.; Bossuyt, P.; Herring, A.; Mabey, D.; Poole, F.; Smith, P.G.; Sriram, N.; Wongsrichanalai, C.; Linke, R.; et al. Evaluation of diagnostic tests for infectious diseases: General principles. Nat. Rev. Microbiol. 2010, 8, 17–29. [Google Scholar] [CrossRef]
- Bossuyt, P.M.M.; Reitsma, J.B.; Linnet, K.; Moons, K.G.M. Beyond Diagnostic Accuracy: The Clinical Utility of Diagnostic Tests. Clin. Chem. 2012, 58, 1636–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante di Ruffano, L.; Hyde, C.J.; Mccaffery, K.J.; Bossuyt, P.M.M.; Deeks, J.J. Assessing the value of diagnostic tests: A framework for designing and evaluating trials. BMJ 2012, 344, e686. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Parker, S.K.; Todd, J.K.; Dominguez, S.R. Implementation of Rapid Molecular Infectious Disease Diagnostics: The Role of Diagnostic and Antimicrobial Stewardship. J. Clin. Microbiol. 2017, 55, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Siontis, K.C.; Siontis, G.C.M.; Contopoulos-ioannidis, D.G.; Ioannidis, J.P.A. Diagnostic tests often fail to lead to changes in patient outcomes. J. Clin. Epidemiol. 2014, 67, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Van den Bruel, A.; Cleemput, I.; Aertgeerts, B.; Ramaekers, D.; Buntinx, F. The evaluation of diagnostic tests: Evidence on technical and diagnostic accuracy, impact on patient outcome and cost-effectiveness is needed. J. Clin. Epidemiol. 2007, 60, 1116–1122. [Google Scholar] [CrossRef] [PubMed]



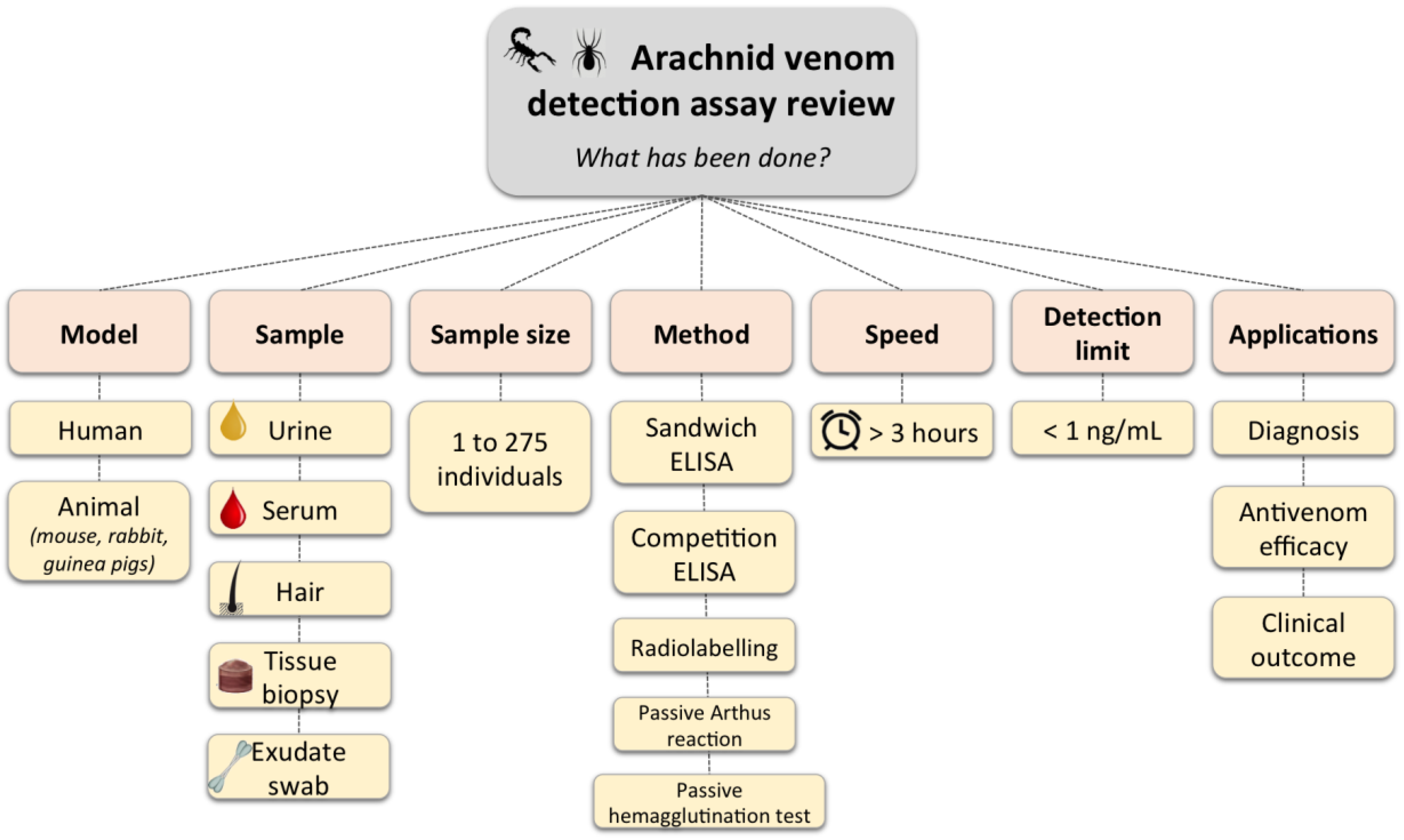{kind=link}
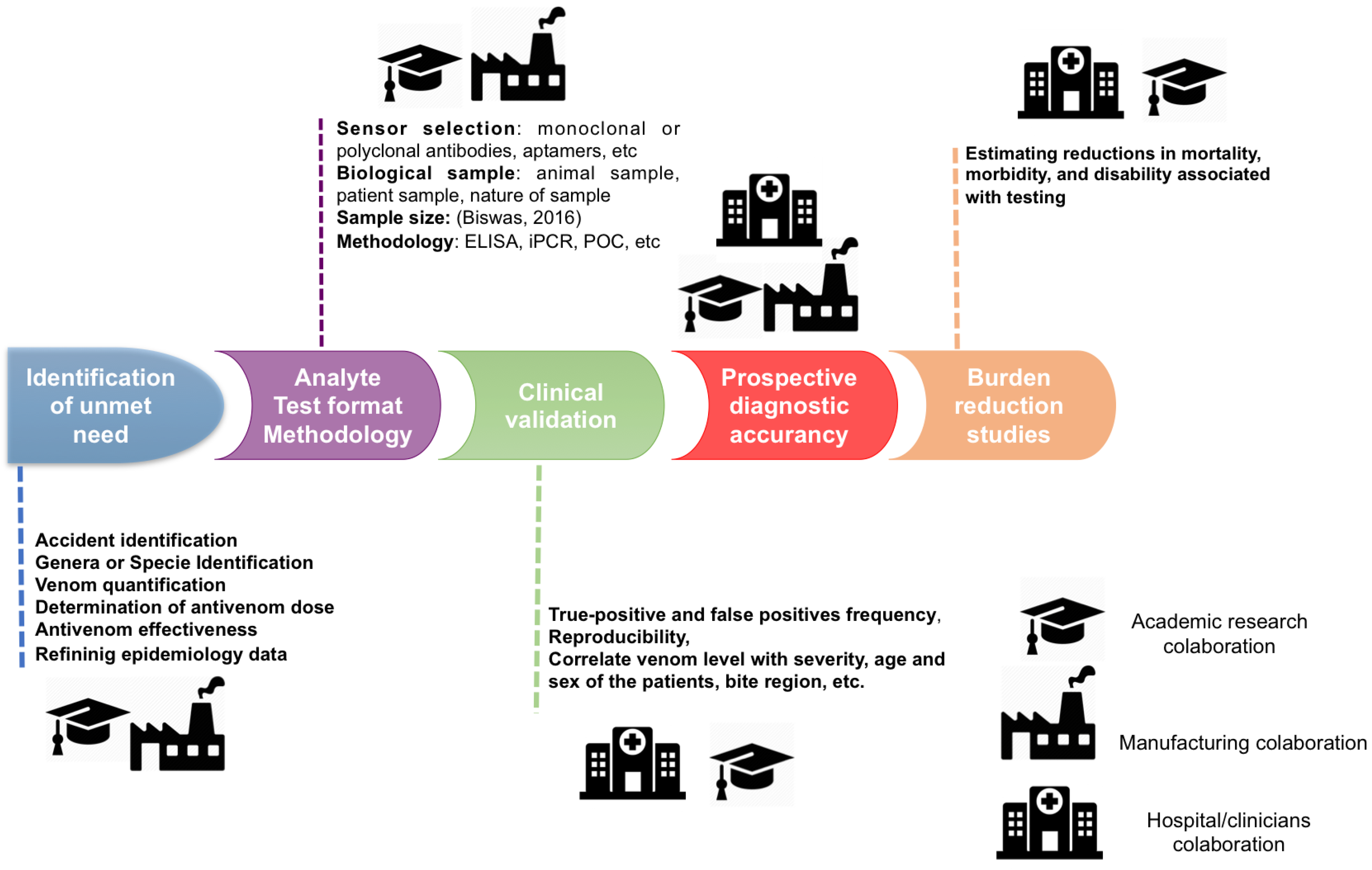{kind=link}
| Target | Sample | Model | Technology | Amount Injected/Route | Time after Injection | Amount Detected | Detection Limit | Reference |
|---|---|---|---|---|---|---|---|---|
| IgG anti-Loxosceles venom | Serum | Human (n = 20) | Indirect ELISA | Bite | 9 to 120 days | Detectable in 4 patients | ND | [31] |
| Loxosceles venom | Skin exudate | Guinea pigs (n=26) | Passive Hemagglutination Inhibition Test | 24 μg/i.d | Up to 3 days | Detectable | ND | [23] |
| Serum | Mouse (n = 10) | Sandwich ELISA | 2.5 μg/s.c | 0.25 h | Not detectable | <0.1 ng | [30] | |
| 0.50 h | 60 ng/mL | |||||||
| 1.0 h | 45 ng/mL | |||||||
| 2.0 h | 40 ng/mL | |||||||
| 4.0 h | 10 ng/mL | |||||||
| 8.0 h | 5 ng/mL | |||||||
| 12.0 h, 1, 2 and 3 days | Not detectable | |||||||
| Biopsy | Human (n = 1) | Competitive ELISA | Bite | 4 days | 3350 pg/2-mm biopsy | ND | [29] | |
| Hair | Competitive ELISA (Strep-Biot) | 1113 pg/100 μL | ||||||
| Biopsy | Rabbits (n = 3) | Sandwich ELISA | 3 μg/i.d | 1 day | 4280 pg/4-mm biopsy | <0.1 ng | [27] | |
| Biopsy | Rabbits (n = 3) | Sandwich ELISA | 3 μg/i.d | 7 days | 205 pg/4-mm biopsy | <0.1 ng | [28] | |
| Serum | 1, 2, 3, 4 and 7 days | Not detectable | ||||||
| Hair | 1 day | 216 pg/100 μL | ||||||
| 2 days | 192 pg/100 μL | |||||||
| 3 days | 172 pg/100 μL | |||||||
| 4 days | 148 pg/100 μL | |||||||
| 7 days | 80 pg/100 μL | |||||||
| Loxosceles venom | Skin exudate | Human (n = 1) | Sandwich ELISA | Bite | 3 days | 34 pg/100 μL | 24 pg | [24] |
| Serum | Not detectable | |||||||
| Skin exudate | Human (n = 1) | Sandwich ELISA | Bite | 13 days | Detectable | <0.1 ng | [25] | |
| Skin exudate | Rabbits (n = 3) | Sandwich ELISA | 4–5 μg/s.c | 7, 10, 14 and 21 days | ~5 pg | ND | [22] | |
| Biopsy | 1 and 3 days | Not detectable | ||||||
| Phoneutria venom | Serum | Mice (n = 5) | Sandwich ELISA | 5 μg/s.c | 0.50 h | 25 ng/mL | <2 ng | [34] |
| Human (n = 2) | Bite | ND | 11–26 ng/mL |
| Target | Sample | Model | Technology | Amount Injected/Route | Time after Injection | Amount Detected (Normalized) | Detection Limit | Reference |
|---|---|---|---|---|---|---|---|---|
| Tityus venom | Serum | Human (n = 40) | Sandwich ELISA | Sting | ND | * | 0.1 ng/mL | [36] |
| Serum | Mice (n = 10) | Sandwich ELISA | 1 µg/s.c | 0.5 h | * | 0.1 ng/mL | ||
| Serum | Human (n = 56) | Sandwich ELISA | Sting | ND | * | 4.8 ng/mL | [19] | |
| Serum | Human (n = 19) | Sandwich ELISA | Sting | 1.5 h | 2.14–50 ng/mL | ND | [18] | |
| Serum | Human (n = 205) | Sandwich ELISA (Strep-Biot) | Sting | 0.5–6.0 h | 0.09–202 ng/mL | 0.09 ng/mL | [38] | |
| Androctonus and Buthus venom | Serum | Human (n = 180) | Sandwich ELISA | Sting | 5 to 4.8 h | GI-0.9 to 4.2 ng/mL | 0.9 ng/mL | [21] |
| GII-3 to 16 ng/mL | ||||||||
| GIII-13 to 38 ng/mL | ||||||||
| Centruroides venom | Serum | Human (n = 3) | Sandwich ELISA | Sting | 50 min–5.2 h | 8.2–29.7 ng/mL | 1 ng/mL | [40] |
| Urine | ~490 min–8.2 h | 9.0 ng/mL | ||||||
| Mesobuthus venom | Skin exudate | Mice (n = 6) | Reverse passive Arthus reaction (RPA) | 100 μg/s.c | 45 min | Detectable in ≈ 84.4% | ND | [42] |
| Antigen | Antivenom | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Target | Model | Amount Injected/Route | Time after Injection | Amount Detected | Antivenom | Amount Injected/Route | Time after Venom for Antivenom Injection | Amount Detected (Time after Antivenom Injection) | Detection Limit | Reference |
| Loxosceles venom | Mice (n = 4) | 10 µg i.d | 0.25 h | 20 ng/mL | Anti-arachnidic (BUTANTAN) | 0.2 mL/animal i.v | Immediately after venom | Not detectable in all time points (0.25 h, 0.5 h, 1.0 h, 4.0 h and 24 h) | 2 ng/mL | [44] |
| 0.5 h | 28 ng/mL | |||||||||
| 1.0 h | 18 ng/mL | |||||||||
| 4.0 h | 7 ng/mL | |||||||||
| 24.0 h | 0 ng/mL | |||||||||
| Phoneutria venom | Mice (n = 4) | 10 µg i.d | 0.25 h | 65 ng/mL | Anti-arachnidic (BUTANTAN) | 0.2 mL/animal i.v | Immediately after venom | 1 ng/mL (0.25 h) | 2 ng/mL | [44] |
| 0.5 h | 50 ng/mL | 1 ng/mL (0.5 h) | ||||||||
| 1.0 h | 48 ng/mL | 1 ng/mL (1.0 h) | ||||||||
| 4.0 h | 17 ng/mL | Not detectable (4.0 h) | ||||||||
| 24.0 h | 0 ng/mL | Not detectable (24 h) | ||||||||
| Human (n = 1) | Bite | 4.0 h | 47.5 ng/mL | Anti-arachnidic (BUTANTAN) | 5 vials i.v | 4.0 h | Not detectable in all time points (1.0 h, 6.0 h, 24 h and 48 h) | 17.1 ng/mL | [35] | |
| Atrax and Hadronyche venom ** | Human (n = 9) | Bite | ND | 0.4–35 ng/mL | Anti-funnel-web spider (CSL) | 2–12 vials i.v | 1.0–5.0 h | Not detectable to 1 ng/mL (various time points) | 0.2 ng/mL | [33] |
| Antigen | Antivenom | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Target | Model | Amount Injected/Route | Time after Injection | Amount Detected | Antivenom | Amount Injected/Route | Time after Venom for Antivenom Injection | Amount Detected (Time after Antivenom Injection) | Detection Limit | Reference |
| Tityus venom | Mice (n = 28) | 10 μg/s.c | 0.25 h | 760 ng/mL | Anti-scorpionic (FUNED) | 10 μL/i.v | Immediately after venom | 41 ng/mL (0.25 h) | ND | [47] |
| 0.5 h | 780 ng/mL | 50 ng/mL (0.5 h) | ||||||||
| 1.0 h | 400 ng/mL | 100 ng/mL (1 h) | ||||||||
| 2.0 h | 360 ng/mL | 100 ng/mL (2 h) | ||||||||
| 4.0 h | 50 ng/mL | 40 ng/mL (4 h) | ||||||||
| 8.0 h | 0 ng/mL | Not detectable (8 h) | ||||||||
| 1.0 h | Not detectable in all time points | |||||||||
| Human (n = 18) | Sting | ND | ≈15.07 ng/mL | Anti-scorpionic (FUNED) | 5–30 mL/i.v | ND | Not detectable (1.0 h) | 0.1 ng/mL | [48] | |
| Mice (n = 4) | 10 µg/i.d | 0.25 h | 34 ng/mL | Anti-arachnidic (BUTANTAN) | 0.2 mL/animal i.v | Immediately after venom | Not detectable in all time points | 2 ng/mL | [44] | |
| 0.5 h | 25 ng/mL | |||||||||
| 1.0 h | 28 ng/mL | |||||||||
| 4.0 h | 18 ng/mL | |||||||||
| 24.0 h | 0 ng/mL | |||||||||
| Androctonus venom | Human (n = 40) | Sting | Up to 2.0 h | GI (n = 31): 0.96 ± 0.36 ng/mL | Horse F(ab′)2 anti-AahFG50 | 10 mL/i.m | ND | 0.63 ± 0.19 ng/mL (1.0 h) | 0.5 ng/mL | [20] |
| 0.49 ± 0.15 ng/mL (3.0 h) | ||||||||||
| GII (n = 9): 2.44 ± 1.1 ng/mL | 1.11 ± 0.57 ng/mL (1.0 h) | |||||||||
| 0.63 ± 0.21 ng/mL (3.0 h) | ||||||||||
| Rabbit (n = 3) # | 100 μg/kg **** s.c | 0–13.0 h | *** | Anti-BotFG50 purified from Bivalent anti-Aag and anti-Bot (PIT) | 80, 8, 4 or 0.8 mg/i.m or i.v | 1.0 h | *** | ND | [41] | |
| Androctonus and Buthus venom | Human (n = 147) | Sting | ND | ≈23 ng/mL (n = 12) | Bivalent anti-Aag and anti-Bot (PIT) | No antivenom | 6 ± 1 ng/mL (6.0 h) | 0.9 ng/mL | [43] | |
| ≈24 ng/mL (n = 28) | 1 × 2 mL/kg (10–30 mL)/i.m | At hospital admission | 4 ± 2 ng/mL (6.0 h) | |||||||
| *** (n = 34) | 1 × 2 mL/kg (10–30 mL)/i.v | 3 ± 1 ng/mL (1.0 h) | ||||||||
| ≈13 ng/mL (n = 16) | 2 × 2 mL /kg (5–15 mL)/i.m | 1st at hospital admission 2nd 1 h later | Not detectable (6.0 h after 2nd injection) | |||||||
| *** (n = 42) | 2 × 2 mL/kg (5–15 mL)/i.m/i.v | Not detectable (0.5 h after 2nd injection) | ||||||||
| *** (n = 15) | 2 × 2 mL /kg (5–15 mL)/i.v | Not detectable (0.5 h after 2nd injection) | ||||||||
| Human (n = 275) | Sting | 5 min–16.0 h (1.27 ± 1.6 h) | ≈18.72 ng/mL (n = 96) | Horse anti-A.mauretanicus mauretanicus | No antivenom | 22.20 ± 27.04 ng/mL (3.0 h) | 0.78 ng/mL | [37] | ||
| ≈12.10 ng/mL (n = 48) | 2–5 mL | 9.86 ± 14.86 ng/mL (1.0 h) | ||||||||
| ND | 8.81 ± 15.63 ng/mL (3.0 h) | |||||||||
| ≈19.35 ng/mL (n = 131) | 10 mL | 6.64 ± 11.12 ng/mL (1.0 h) | ||||||||
| 4.98 ± 15.25 ng/mL (3.0 h) | ||||||||||
| Buthus venom | Rabbit (n = 3–6) | 100 μg/kg s.c | 5 min–4 8h | *** | Anti-BotFG50 purified from Bivalent anti-Aag and anti-Bot (PIT) | 30, 12 and 6 mg/kg in 1 mL i.m or i.v | 0.5–3.0 h | *** | ND | [46] |
| Centruroides venom | Human (n = 44) | Sting | ND | 0–87.4 ng/mL | Alacramyn® (Bioclon, Mexico) | ND | ND | Not detectable to 4.7 ng/mL (0.5 h) | 0.1 ng/mL | [39] |
| Genera | Arachnid Genera Identification from Venom Detected in Human Biological Samples | Diagnostic Test for Clinical Use | Venom Quantification or/and Correlation with Patient Clinical Outcome | Determination of Antivenom Dose for Human Treatment | Determination of Antivenom Treatment Effectiveness in Patients | Refining of Epidemiology Data | Sample Size and References |
|---|---|---|---|---|---|---|---|
| Loxosceles | ✔✔ | ✖ | ✖ | ✖ | ✖ | ✖ | n = 30 [30]; n = 1 [24] |
| Latrodectus | ✖ | ✖ | ✖ | ✖ | ✖ | ✖ | |
| Phoneutria | ✔ | ✖ | ✔ | ✖ | ✖ | ✖ | n = 2 [34]; n = 1 [35] |
| Atrax/Hadronyche | ✔ | ✖ | ✔ | ✔ | ✔ | ✖ | n = 9 [33] |
| Leiurus | ✖ | ✖ | ✖ | ✖ | ✖ | ✖ | |
| Androctonus | ✔✔✔ | ✖ | ✔✔✔ | ✔✔✔ | ✔✔✔ | ✖ | n = 180 [21]; n = 147 [43]; n = 275 [37] |
| Buthus | ✔✔✔ | ✖ | ✔✔✔ | ✔✔✔ | ✔✔✔ | ✖ | n =180 [21]; n = 147 [43]; n = 275 [37] |
| Tityus | ✔✔✔ | ✖ | ✔✔✔ | ✖ | ✔✔ | ✖ | n = 205 [38]; n = 18 [51] n = 40 [36]; n = 56 [19]; n = 19 [18] |
| Centruroides | ✔✔ | ✖ | ✔✔ | ✖ | ✔✔ | ✖ | n = 3 [40]; n = 44 [39] |
| Mesobuthus | ✖ | ✖ | ✖ | ✖ | ✖ | ✖ | |
| Parabuthus | ✖ | ✖ | ✖ | ✖ | ✖ | ✖ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias-Lopes, C.; Paiva, A.L.; Guerra-Duarte, C.; Molina, F.; Felicori, L. Venomous Arachnid Diagnostic Assays, Lessons from Past Attempts. Toxins 2018, 10, 365. https://doi.org/10.3390/toxins10090365
Dias-Lopes C, Paiva AL, Guerra-Duarte C, Molina F, Felicori L. Venomous Arachnid Diagnostic Assays, Lessons from Past Attempts. Toxins. 2018; 10(9):365. https://doi.org/10.3390/toxins10090365
Chicago/Turabian StyleDias-Lopes, Camila, Ana Luiza Paiva, Clara Guerra-Duarte, Franck Molina, and Liza Felicori. 2018. "Venomous Arachnid Diagnostic Assays, Lessons from Past Attempts" Toxins 10, no. 9: 365. https://doi.org/10.3390/toxins10090365
APA StyleDias-Lopes, C., Paiva, A. L., Guerra-Duarte, C., Molina, F., & Felicori, L. (2018). Venomous Arachnid Diagnostic Assays, Lessons from Past Attempts. Toxins, 10(9), 365. https://doi.org/10.3390/toxins10090365