Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving




Abstract
:1. Introduction
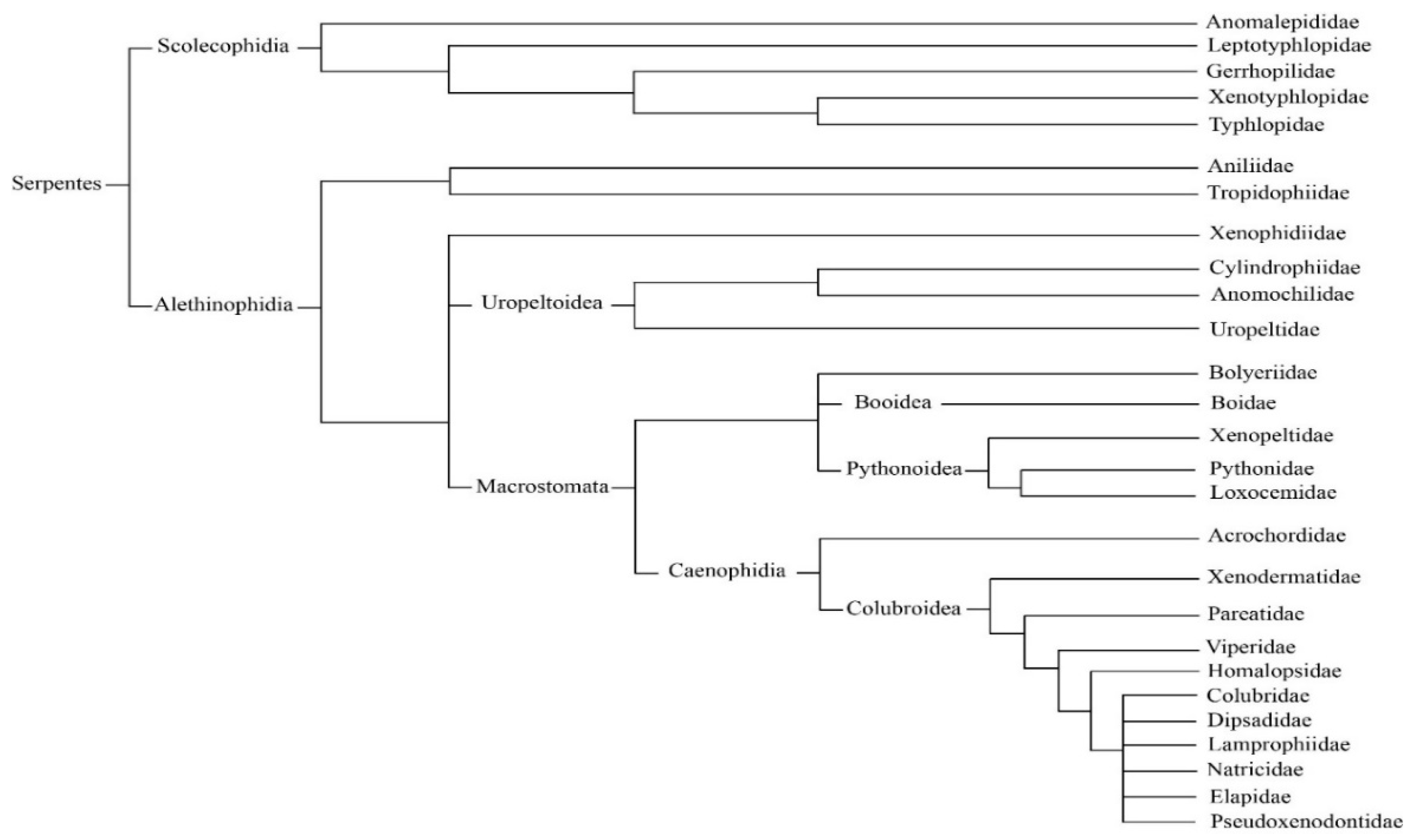
2. Classification and Distribution of Venomous Snakes
3. Composition of Snake Venoms
3.1. Enzymatic Proteins from Snake Venoms
3.1.1. Proteolytic Enzymes
3.1.2. Arginine Ester Hydrolases
3.1.3. Hyaluronidases
3.1.4. Phospholipase A2 (PLA2)
3.1.5. Acetylcholinesterase (AChE)
3.1.6. Nucleases (RNase, DNase, and Phosphodiesterase)
3.1.7. L-amino-Acid Oxidase (LAAO)
3.2. Non-Enzymatic Proteins from Snake Venoms
4. Pathophysiological and Pharmacological Actions of Snake Venoms
4.1. Neurotoxicity
4.1.1. Presynaptic Neurotoxins
4.1.2. Postsynaptic Neurotoxins
4.1.3. Dendrotoxins
4.2. Hemotoxicity
4.2.1. Pro-Coagulant Activities
4.2.2. Anticoagulant Activities
4.2.3. Fibrinolytic Factors
4.3. Cytotoxicity
4.4. Myotoxicity
4.5. Antimicrobial
5. Snake Venoms for Drug Discovery
5.1. FDA-Approved Drugs Derived from Snake Venom Proteins
5.2. Toxin-Derived Drugs from Snake Venom Proteins in Clinical Trials and Stages of Development
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jimenez-Porras, J.M. Pharmacology of peptides and proteins in snake venoms. Annu. Rev. Pharm. 1968, 8, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional Toxins in Snake Venoms and Therapeutic Implications: From Pain to Hemorrhage and Necrosis. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.A., 2nd; Figge, H.L. Treatment of snakebite poisoning. Am. J. Hosp. Pharm. 1991, 48, 2190–2196. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO: Geneva, Switzerland, 2018; pp. 1–192. Available online: https://www.who.int/bloodproducts/snake_antivenoms/snakeantivenomguide/en/ (accessed on 2 July 2019).
- Hardy, M.C.; Cochrane, J.; Allavena, R.E. Venomous and poisonous Australian animals of veterinary importance: A rich source of novel therapeutics. Biomed. Res. Int. 2014, 2014, 671041. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.L.; Rowan, E.G.; Albericio, F.; Stabeli, R.G.; Calderon, L.A.; Soares, A.M. Animal toxins and their advantages in biotechnology and pharmacology. Biomed. Res. Int. 2014, 2014, 951561. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Tytgat, J. Toxins in Drug Discovery and Pharmacology. Toxins 2018, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Why do we study animal toxins? Dongwuxue Yanjiu 2015, 36, 183–222. [Google Scholar] [CrossRef]
- Beraud, E.; Chandy, K.G. Therapeutic potential of peptide toxins that target ion channels. Inflamm. Allergy Drug Targets 2011, 10, 322–342. [Google Scholar] [CrossRef]
- Hawgood, B.J. Abbe Felice Fontana (1730–1805): Founder of modern toxinology. Toxicon 1995, 33, 591–601. [Google Scholar] [CrossRef]
- Waheed, H.; Moin, S.F.; Choudhary, M.I. Snake Venom: From Deadly Toxins to Life-saving Therapeutics. Curr. Med. Chem. 2017, 24, 1874–1891. [Google Scholar] [CrossRef]
- Warrell, D.A. 75 - Venomous and Poisonous Animals. In Manson’s Tropical Infectious Diseases, 23rd ed.; Farrar, J., Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; W.B. Saunders: London, UK, 2014; pp. 1096–1127.e1093. [Google Scholar] [CrossRef]
- He, Q.Y.; He, Q.Z.; Deng, X.C.; Yao, L.; Meng, E.; Liu, Z.H.; Liang, S.P. ATDB: A uni-database platform for animal toxins. Nucleic Acids Res. 2008, 36, D293–D297. [Google Scholar] [CrossRef] [PubMed]
- White, J. Venomous animals: Clinical toxinology. EXS 2010, 100, 233–291. [Google Scholar] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D. Venomous and Poisonous Animals: A Handbook for Biologists, Toxicologists and Toxinologists, Physicians and Pharmacists; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Smith, M.A. The Classification of Snakes in accordance with their Dentition and the Evolution of the Poison Fang: Section of Tropical Diseases and Parasitology. Proc. R. Soc. Med. 1934, 27, 1081–1083. [Google Scholar] [PubMed]
- Pyron, R.A.; Burbrink, F.T.; Colli, G.R.; de Oca, A.N.; Vitt, L.J.; Kuczynski, C.A.; Wiens, J.J. The phylogeny of advanced snakes (Colubroidea), with discovery of a new subfamily and comparison of support methods for likelihood trees. Mol. Phylogenet Evol. 2011, 58, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Tu, A.T. Overview of snake venom chemistry. Adv. Exp. Med. Biol. 1996, 391, 37–62. [Google Scholar]
- Gopalakrishnakone, P.; Inagaki, H. Snake Venoms; Springer: Berlin, Germany, 2017. [Google Scholar]
- Kumar, V.; Rejent, T.A.; Elliott, W.B. Anticholinesterase activity of elapid venoms. Toxicon 1973, 11, 131–138. [Google Scholar] [CrossRef]
- Chippaux, J.P.; Williams, V.; White, J. Snake venom variability: Methods of study, results and interpretation. Toxicon 1991, 29, 1279–1303. [Google Scholar] [CrossRef]
- Tu, A.T. Venoms: Chemistry and Molecular Biology; Jhon Wiley & Sons Inc.: New York, NY, USA, 1977. [Google Scholar]
- Alape-Giron, A.; Sanz, L.; Escolano, J.; Flores-Diaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancehead pitviper Bothrops asper: Geographic, individual, and ontogenetic variations. J. Proteome Res. 2008, 7, 3556–3571. [Google Scholar] [CrossRef]
- Saldarriaga, M.M.; Otero, R.; Nunez, V.; Toro, M.F.; Diaz, A.; Gutierrez, J.M. Ontogenetic variability of Bothrops atrox and Bothrops asper snake venoms from Colombia. Toxicon 2003, 42, 405–411. [Google Scholar] [CrossRef]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.; Serrano, S.M. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Sasa, M. Diet and snake venom evolution: Can local selection alone explain intraspecific venom variation? Toxicon 1999, 37, 249–252, author reply 253–260. [Google Scholar]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim. Et Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Bottrall, J.L.; Madaras, F.; Biven, C.D.; Venning, M.G.; Mirtschin, P.J. Proteolytic activity of Elapid and Viperid Snake venoms and its implication to digestion. J. Venom. Res. 2010, 1, 18–28. [Google Scholar] [PubMed]
- Preciado, L.M.; Pereañez, J.A. Low molecular mass natural and synthetic inhibitors of snake venom metalloproteinases. Toxin Rev. 2018, 37, 19–26. [Google Scholar] [CrossRef]
- Birrell, G.W.; Earl, S.T.; Wallis, T.P.; Masci, P.P.; de Jersey, J.; Gorman, J.J.; Lavin, M.F. The diversity of bioactive proteins in Australian snake venoms. Mol. Cell Proteom. 2007, 6, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Kress, L.F. Inactivation of human plasma serine proteinase inhibitors (serpins) by limited proteolysis of the reactive site loop with snake venom and bacterial metalloproteinases. J. Cell. Biochem. 1986, 32, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. The intriguing world of prothrombin activators from snake venom. Toxicon 2005, 45, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-J.; Shih, C.-H.; Huang, T.-F. Primary structure and antiplatelet mechanism of a snake venom metalloproteinase, acurhagin, from Agkistrodon acutus venom. Biochimie 2005, 87, 1065–1077. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Markland, F.S., Jr.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Padrón, O.; Castro-Guillén, J.L.; García-Arredondo, J.A.; Cruz-Pérez, M.S.; Díaz-Peña, L.F.; Saldaña, C.; Blanco-Labra, A.; García-Gasca, T. Snake Venom Hemotoxic Enzymes: Biochemical Comparison between Crotalus Species from Central Mexico. Molecules 2019, 24, 1489. [Google Scholar] [CrossRef] [PubMed]
- Zaqueo, K.D.; Kayano, A.M.; Domingos, T.F.S.; Moura, L.A.; Fuly, A.L.; da Silva, S.L.; Acosta, G.; Oliveira, E.; Albericio, F.; Zanchi, F.B.; et al. BbrzSP-32, the first serine protease isolated from Bothrops brazili venom: Purification and characterization. Comp. Biochem. Physiol. Part. A: Mol. Integr. Physiol. 2016, 195, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.M.; Yu, H.; Liu, Z.Z.; Pei, J.Z.; Yang, Y.E.; Yan, S.X.; Zhang, C.; Zhao, W.L.; Wang, Z.Z.; Wang, Y.M.; et al. Serine protease isoforms in Gloydius intermedius venom: Full sequences, molecular phylogeny and evolutionary implications. J. Proteom. 2017, 164, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Zaqueo, K.D.; Kayano, A.M.; Simões-Silva, R.; Moreira-Dill, L.S.; Fernandes, C.F.; Fuly, A.L.; Maltarollo, V.G.; Honório, K.M.; da Silva, S.L.; Acosta, G.; et al. Isolation and Biochemical Characterization of a New Thrombin-Like Serine Protease from Bothrops pirajai Snake Venom. BioMed Res. Int. 2014, 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Masood, R.; Ali, I.; Ullah, K.; Ali, H.; Akbar, H.; Betzel, C. Thrombin-like enzymes from snake venom: Structural characterization and mechanism of action. Int. J. Biol. Macromol. 2018, 114, 788–811. [Google Scholar] [CrossRef]
- Nolan, C.; Hall, L.S.; Barlow, G.H. Ancrod, the coagulating enzyme from Malayan pit viper (Agkistrodon rhodostoma) venom. Methods Enzym. 1976, 45, 205–213. [Google Scholar]
- Markland, F.S.; Damus, P.S. Purification and properties of a thrombin-like enzyme from the venom of Crotalus adamanteus (Eastern diamondback rattlesnake). J. Biol. Chem. 1971, 246, 6460–6473. [Google Scholar]
- Liu, S.; Sun, M.Z.; Sun, C.; Zhao, B.; Greenaway, F.T.; Zheng, Q. A novel serine protease from the snake venom of Agkistrodon blomhoffii ussurensis. Toxicon 2008, 52, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Hahn, B.S.; Yang, K.Y.; Park, E.M.; Chang, I.M.; Kim, Y.S. Purification and molecular cloning of calobin, a thrombin-like enzyme from Agkistrodon caliginosus (Korean viper). J. Biochem. 1996, 119, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Oyama, E.; Takahashi, H. Purification and characterization of a thrombin-like enzyme, elegaxobin, from the venom of Trimeresurus elegans (Sakishima-habu). Toxicon 2000, 38, 1087–1100. [Google Scholar] [CrossRef]
- Sant’Ana, C.D.; Ticli, F.K.; Oliveira, L.L.; Giglio, J.R.; Rechia, C.G.V.; Fuly, A.L.; Selistre de Araújo, H.S.; Franco, J.J.; Stabeli, R.G.; Soares, A.M.; et al. BjussuSP-I: A new thrombin-like enzyme isolated from Bothrops jararacussu snake venom. Comp. Biochem. Physiol. Part. A: Mol. Integr. Physiol. 2008, 151, 443–454. [Google Scholar] [CrossRef]
- Serrano, S.M.T. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Oshima, G.; Sato-Ohmori, T.; Suzuki, T. Proteinase, arginineester hydrolase and a kinin releasing enzyme in snake venoms. Toxicon 1969, 7, 229–233. [Google Scholar] [CrossRef]
- Andriao-Escarso, S.H.; Soares, A.M.; Rodrigues, V.M.; Mancin, A.C.; Reis, M.L.; Ballejo, G.; Giglio, J.R. Isolation and characterization of an arginine ester hydrolase from Bothrops jararacussu venom which induces contractions of the isolated rat uterus. Biochem. Mol. Biol. Int. 1999, 47, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Sim, S.; Gnanathasan, C.; Fung, S.; Ponnudurai, G.; Pailoor, J.; Tan, N. Enzymatic and toxinological activities of Hypnale hypnale (hump-nosed pit viper) venom and its fractionation by ion exchange high performance liquid chromatography. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 473–485. [Google Scholar] [CrossRef]
- Bala, E.; Hazarika, R.; Singh, P.; Yasir, M.; Shrivastava, R. A biological overview of Hyaluronidase: A venom enzyme and its inhibition with plants materials. Mater. Today: Proc. 2018, 5, 6406–6412. [Google Scholar] [CrossRef]
- Bordon, K.C.; Perino, M.G.; Giglio, J.R.; Arantes, E.C. Isolation, enzymatic characterization and antiedematogenic activity of the first reported rattlesnake hyaluronidase from Crotalus durissus terrificus venom. Biochimie 2012, 94, 2740–2748. [Google Scholar] [CrossRef]
- Bhavya, J.; Vineetha, M.S.; Sundaram, P.M.; Veena, S.M.; Dhananjaya, B.L.; More, S.S. Low-molecular weight hyaluronidase from the venom of Bungarus caeruleus (Indian common krait) snake: Isolation and partial characterization. J. Liq. Chromatogr. Relat. Technol. 2016, 39, 203–208. [Google Scholar] [CrossRef]
- Van Deenen, L.L.M.; De Haas, G.H.; Heemskerk, C.T. Hydrolysis of synthetic mixed-acid phosphatides by phospholipase A from human pancreas. Biochim. Biophys. Acta (BBA) -Spec. Sect. Enzymol. Subj. 1963, 67, 295–304. [Google Scholar] [CrossRef]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. of Lipids 2006, 1761, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Terra, A.L.C.; Moreira-Dill, L.S.; Simões-Silva, R.; Monteiro, J.R.N.; Cavalcante, W.L.G.; Gallacci, M.; Barros, N.B.; Nicolete, R.; Teles, C.B.G.; Medeiros, P.S.M.; et al. Biological characterization of the Amazon coral Micrurus spixii snake venom: Isolation of a new neurotoxic phospholipase A2. Toxicon 2015, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cedro, R.C.A.; Menaldo, D.L.; Costa, T.R.; Zoccal, K.F.; Sartim, M.A.; Santos-Filho, N.A.; Faccioli, L.H.; Sampaio, S.V. Cytotoxic and inflammatory potential of a phospholipase A2 from Bothrops jararaca snake venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 33. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Scheib, H.; van der Weerd, L.; Young, B.; McNaughtan, J.; Ramjan, S.F.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an arsenal: Structural and functional diversification of the venom system in the advanced snakes (Caenophidia). Mol. Cell Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [PubMed]
- Vanzolini, K.L.; Ainsworth, S.; Bruyneel, B.; Herzig, V.; Seraus, M.G.L.; Somsen, G.W.; Casewell, N.R.; Cass, Q.B.; Kool, J. Rapid ligand fishing for identification of acetylcholinesterase-binding peptides in snake venom reveals new properties of dendrotoxins. Toxicon 2018, 152, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cousin, X.; Creminon, C.; Grassi, J.; Meflah, K.; Cornu, G.; Saliou, B.; Bon, S.; Massoulie, J.; Bon, C. Acetylcholinesterase from Bungarus venom: A monomeric species. FEBS Lett. 1996, 387, 196–200. [Google Scholar] [CrossRef]
- Frobert, Y.; Créminon, C.; Cousin, X.; Rémy, M.-H.; Chatel, J.-M.; Bon, S.; Bon, C.; Grassi, J. Acetylcholinesterases from Elapidae snake venoms: Biochemical, immunological and enzymatic characterization. Biochim. Et Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1997, 1339, 253–267. [Google Scholar] [CrossRef]
- Sales, P.B.V.; Santoro, M.L. Nucleotidase and DNase activities in Brazilian snake venoms. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2008, 147, 85–95. [Google Scholar] [CrossRef]
- Dhananjaya, B.L.; CJ, D.S. An overview on nucleases (DNase, RNase, and phosphodiesterase) in snake venoms. Biochem. (Mosc) 2010, 75, 1–6. [Google Scholar] [CrossRef] [PubMed]
- De Roodt, A.R.; Litwin, S.; Angel, S.O. Hydrolysis of DNA by 17 snake venoms. Comp. Biochem. Physiol. C Toxicol. Pharm. 2003, 135, 469–479. [Google Scholar] [CrossRef]
- Sittenfeld, A.; Raventós, H.; Cruz, R.; Gutiérrez, J.M. DNase activity in Costa Rican crotaline snake venoms: Quantification of activity and identification of electrophoretic variants. Toxicon 1991, 29, 1213–1224. [Google Scholar] [CrossRef]
- Trummal, K.; Tonismagi, K.; Aaspollu, A.; Siigur, J.; Siigur, E. Vipera lebetina venom nucleases. Toxicon 2016, 119, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Taborda, A.R.; Taborda, L.C.; Williams, J.N., Jr.; Elvehjem, C.A. A study of the ribonuclease activity of snake venoms. J. Biol. Chem. 1952, 194, 227–233. [Google Scholar] [PubMed]
- Mahalakshmi, Y.V.; Jagannadham, M.V.; Pandit, M.W. Ribonuclease from cobra snake venom: Purification by affinity chromatography and further characterization. Iubmb Life 2000, 49, 309–316. [Google Scholar] [CrossRef]
- Halim, H.Y.; Shaban, E.A.; Hagag, M.M.; Daoud, E.W.; El-Asmar, M.F. Purification and characterization of phosphodiesterase (exonuclease) from Cerastes cerastes (Egyptian sand viper) venom. Toxicon 1987, 25, 1199–1207. [Google Scholar] [CrossRef]
- Trummal, K.; Aaspollu, A.; Tonismagi, K.; Samel, M.; Subbi, J.; Siigur, J.; Siigur, E. Phosphodiesterase from Vipera lebetina venom-structure and characterization. Biochimie 2014, 106, 48–55. [Google Scholar] [CrossRef]
- Mamillapalli, R.; Haimovitz, R.; Ohad, M.; Shinitzky, M. Enhancement and inhibition of snake venom phosphodiesterase activity by lysophospholipids. FEBS Lett. 1998, 436, 256–258. [Google Scholar] [CrossRef]
- More, S.; Kiran, K.; Veena, S.; Gadag, J. Purification of an L-amino acid oxidase from Bungarus caeruleus (Indian krait) venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 60–76. [Google Scholar] [CrossRef]
- Abdelkafi-Koubaa, Z.; Morjen, M.; Srairi-Abid, N.; El Ayeb, M.; Marrakchi, N. [Snake Venom L-Amino Acid Oxidases potential biomedical applications]. Arch. Inst. Pasteur Tunis 2014, 91, 15–32. [Google Scholar] [PubMed]
- Izidoro, L.F.; Sobrinho, J.C.; Mendes, M.M.; Costa, T.R.; Grabner, A.N.; Rodrigues, V.M.; da Silva, S.L.; Zanchi, F.B.; Zuliani, J.P.; Fernandes, C.F.; et al. Snake venom L-amino acid oxidases: Trends in pharmacology and biochemistry. Biomed. Res. Int. 2014, 2014, 196754. [Google Scholar] [CrossRef] [PubMed]
- Costal-Oliveira, F.; Stransky, S.; Guerra-Duarte, C.; Naves de Souza, D.L.; Vivas-Ruiz, D.E.; Yarlequé, A.; Sanchez, E.F.; Chávez-Olórtegui, C.; Braga, V.M.M. L-amino acid oxidase from Bothrops atrox snake venom triggers autophagy, apoptosis and necrosis in normal human keratinocytes. Sci. Rep. 2019, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal protein toxins: Origins and therapeutic applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed]
- McCleary, R.J.R.; Kini, R.M. Non-enzymatic proteins from snake venoms: A gold mine of pharmacological tools and drug leads. Toxicon 2013, 62, 56–74. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Molecular moulds with multiple missions: Functional sites in three-finger toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Kostiza, T.; Meier, J. Nerve growth factors from snake venoms: Chemical properties, mode of action and biological significance. Toxicon 1996, 34, 787–806. [Google Scholar] [CrossRef]
- Higuchi, S.; Murayama, N.; Saguchi, K.; Ohi, H.; Fujita, Y.; Camargo, A.C.M.; Ogawa, T.; Deshimaru, M.; Ohno, M. Bradykinin-potentiating peptides and C-type natriuretic peptides from snake venom. Immunopharmacology 1999, 44, 129–135. [Google Scholar] [CrossRef]
- Kordiš, D.; Gubenšek, F. Adaptive evolution of animal toxin multigene families. Gene 2000, 261, 43–52. [Google Scholar] [CrossRef]
- Abd El-Aziz, T.M.; Al Khoury, S.; Jaquillard, L.; Triquigneaux, M.; Martinez, G.; Bourgoin-Voillard, S.; Seve, M.; Arnoult, C.; Beroud, R.; De Waard, M. Actiflagelin, a new sperm activator isolated from Walterinnesia aegyptia venom using phenotypic screening. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 2. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Mackessy, S.P.; Dutta, S. Characterization of a Kunitz-type protease inhibitor peptide (Rusvikunin) purified from Daboia russelii russelii venom. Int. J. Biol. Macromol. 2014, 67, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Imredy, J.P.; MacKinnon, R. Energetic and structural interactions between delta-dendrotoxin and a voltage-gated potassium channel. J. Mol. Biol. 2000, 296, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Titani, K.; Usami, Y.; Suzuki, M.; Oyama, R.; Matsui, T.; Fukui, H.; Sugimoto, M.; Ruggeri, Z.M. Isolation and chemical characterization of two structurally and functionally distinct forms of botrocetin, the platelet coagglutinin isolated from the venom of Bothrops jararaca. Biochemistry 1991, 30, 1957–1964. [Google Scholar] [CrossRef] [PubMed]
- Hamako, J.; Matsui, T.; Suzuki, M.; Ito, M.; Makita, K.; Fujimura, Y.; Ozeki, Y.; Titani, K. Purification and characterization of bitiscetin, a novel von Willebrand factor modulator protein from Bitis arietans snake venom. Biochem. Biophys. Res. Commun. 1996, 226, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Wijeyewickrema, L.C.; Gardiner, E.E.; Gladigau, E.L.; Berndt, M.C.; Andrews, R.K. Nerve growth factor inhibits metalloproteinase-disintegrins and blocks ectodomain shedding of platelet glycoprotein VI. J. Biol. Chem. 2010, 285, 11793–11799. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Itoh, T.; Anuruddhe, B.M.; Bandaranayake, I.K.; Shirani Ranasinghe, J.G.; Athauda, S.B.; Moriyama, A. Molecular diversity in venom proteins of the Russell’s viper (Daboia russellii russellii) and the Indian cobra (Naja naja) in Sri Lanka. Biomed. Res. 2010, 31, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ianzer, D.; Konno, K.; Marques-Porto, R.; Vieira Portaro, F.C.; Stocklin, R.; Martins de Camargo, A.C.; Pimenta, D.C. Identification of five new bradykinin potentiating peptides (BPPs) from Bothrops jararaca crude venom by using electrospray ionization tandem mass spectrometry after a two-step liquid chromatography. Peptides 2004, 25, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Politi, V.; De Luca, G.; Di Stazio, G.; Schinina, E.; Bossa, F. A new peptide from Crotalus atrox snake venom. Peptides 1985, 6 (Suppl. 3), 343–346. [Google Scholar] [CrossRef]
- Da Silva, S.L.; Dias-Junior, C.A.; Baldasso, P.A.; Damico, D.C.; Carvalho, B.M.; Garanto, A.; Acosta, G.; Oliveira, E.; Albericio, F.; Soares, A.M.; et al. Vascular effects and electrolyte homeostasis of the natriuretic peptide isolated from Crotalus oreganus abyssus (North American Grand Canyon rattlesnake) venom. Peptides 2012, 36, 206–212. [Google Scholar] [CrossRef]
- Evangelista, J.S.; Martins, A.M.; Nascimento, N.R.; Sousa, C.M.; Alves, R.S.; Toyama, D.O.; Toyama, M.H.; Evangelista, J.J.; Menezes, D.B.; Fonteles, M.C.; et al. Renal and vascular effects of the natriuretic peptide isolated from Crotalus durissus cascavella venom. Toxicon 2008, 52, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Ranawaka, U.K.; Lalloo, D.G.; de Silva, H.J. Neurotoxicity in snakebite-The limits of our knowledge. Plos Negl. Trop. Dis. 2013, 7, e2302. [Google Scholar] [CrossRef]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Antivenom for Neuromuscular Paralysis Resulting from Snake Envenoming. Toxins 2017, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, U.; Dissanayake, S. Neurological manifestations of snake bite in Sri Lanka. J. Postgrad. Med. 2002, 48, 275–278, discussion 278–279. [Google Scholar] [PubMed]
- Warrell, D.A.; Looareesuwan, S.; White, N.J.; Theakston, R.D.; Warrell, M.J.; Kosakarn, W.; Reid, H.A. Severe neurotoxic envenoming by the Malayan krait Bungarus candidus (Linnaeus): Response to antivenom and anticholinesterase. Br. Med. J. (Clin. Res. Ed.) 1983, 286, 678–680. [Google Scholar] [CrossRef] [PubMed]
- Weinelt, W.; Sattler, R.W.; Mebs, D. Persistent paresis of the facialis muscle after European adder (Vipera berus) bite on the forehead. Toxicon 2002, 40, 1627–1629. [Google Scholar] [CrossRef]
- Malina, T.; Krecsak, L.; Warrell, D.A. Neurotoxicity and hypertension following European adder (Vipera berus berus) bites in Hungary: Case report and review. Qjm: Mon. J. Assoc. Physicians 2008, 101, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, W.C.; Wickramaratna, J.C. In vitro neuromuscular activity of snake venoms. Clin. Exp. Pharmacol. Physiol. 2002, 29, 807–814. [Google Scholar] [CrossRef]
- Harvey, A.L. Presynaptic effects of toxins. Int. Rev. Neurobiol. 1990, 32, 201–239. [Google Scholar]
- Rowan, E.G.; Pemberton, K.E.; Harvey, A.L. On the blockade of acetylcholine release at mouse motor nerve terminals by beta-bungarotoxin and crotoxin. Br. J. Pharm. 1990, 100, 301–304. [Google Scholar] [CrossRef]
- Strong, P.N.; Goerke, J.; Oberg, S.G.; Kelly, R.B. beta-Bungarotoxin, a pre-synaptic toxin with enzymatic activity. Proc. Natl. Acad. Sci. USA 1976, 73, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.; Eaker, D.; Ryden, L. Purification of a presynaptic neurotoxin from the venom of the australian tiger snake Notechis scutatus scutatus. Toxicon 1972, 10, 405–413. [Google Scholar] [CrossRef]
- Su, M.J.; Chang, C.C. Presynaptic effects of snake venom toxins which have phospholipase A2 activity (beta-bungarotoxin, taipoxin, crotoxin). Toxicon 1984, 22, 631–640. [Google Scholar] [CrossRef]
- Su, M.J.; Coulter, A.R.; Sutherland, S.K.; Chang, C.C. The presynaptic neuromuscular blocking effect and phospholipase A2 activity of textilotoxin, a potent toxin isolated from the venom of the Australian brown snake, Pseudonaja textilis. Toxicon 1983, 21, 143–151. [Google Scholar] [CrossRef]
- Tsetlin, V. Snake venom α-neurotoxins and other ‘three-finger’ proteins. Eur. J. Biochem. 1999, 264, 281–286. [Google Scholar] [CrossRef]
- Lewis, R.L.; Gutmann, L. Snake venoms and the neuromuscular junction. Semin Neurol 2004, 24, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.; Armugam, A.; Jeyaseelan, K. Postsynaptic short-chain neurotoxins from Pseudonaja textilis. Eur. J. Biochem. 1999, 265, 982–989. [Google Scholar] [CrossRef]
- Lumsden, N.G.; Banerjee, Y.; Kini, R.M.; Kuruppu, S.; Hodgson, W.C. Isolation and characterization of rufoxin, a novel protein exhibiting neurotoxicity from venom of the psammophiine, Rhamphiophis oxyrhynchus (Rufous beaked snake). Neuropharmacology 2007, 52, 1065–1070. [Google Scholar] [CrossRef]
- Lumsden, N.G.; Fry, B.G.; Ventura, S.; Kini, R.M.; Hodgson, W.C. Pharmacological characterisation of a neurotoxin from the venom of Boiga dendrophila (mangrove catsnake). Toxicon 2005, 45, 329–334. [Google Scholar] [CrossRef]
- Dufton, M.J.; Harvey, A.L. The long and the short of snake toxins. Trends Pharmacol. Sci. 1989, 10, 258–259. [Google Scholar] [CrossRef]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Karatt-Vellatt, A.; Masters, E.W.; Arias, A.S.; Pus, U.; Knudsen, C.; Oscoz, S.; Slavny, P.; Griffiths, D.T.; Luther, A.M.; et al. In vivo neutralization of dendrotoxin-mediated neurotoxicity of black mamba venom by oligoclonal human IgG antibodies. Nat. Commun. 2018, 9, 3928. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Recent studies on dendrotoxins and potassium ion channels. Gen. Pharmacol: Vasc. Syst. 1997, 28, 7–12. [Google Scholar] [CrossRef]
- Benishin, C.G.; Sorensen, R.G.; Brown, W.E.; Krueger, B.K.; Blaustein, M.P. Four polypeptide components of green mamba venom selectively block certain potassium channels in rat brain synaptosomes. Mol. Pharm. 1988, 34, 152–159. [Google Scholar]
- Harvey, A.L. Twenty years of dendrotoxins. Toxicon 2001, 39, 15–26. [Google Scholar] [CrossRef]
- Grissmer, S.; Nguyen, A.N.; Aiyar, J.; Hanson, D.C.; Mather, R.J.; Gutman, G.A.; Karmilowicz, M.J.; Auperin, D.D.; Chandy, K.G. Pharmacological characterization of five cloned voltage-gated K+ channels, types Kv1.1, 1.2, 1.3, 1.5, and 3.1, stably expressed in mammalian cell lines. Mol. Pharm. 1994, 45, 1227–1234. [Google Scholar]
- Harvey, A.L.; Robertson, B. Dendrotoxins: Structure-Activity Relationships and Effects on Potassium Ion Channels. Curr. Med. Chem. 2004, 11, 3065–3072. [Google Scholar] [CrossRef]
- Hutton, R.A.; Warrell, D.A. Action of snake venom components on the haemostatic system. Blood Rev. 1993, 7, 176–189. [Google Scholar] [CrossRef]
- Ouyang, C.; Teng, C.-M.; Huang, T.-F. Characterization of snake venom components acting on blood coagulation and platelet function. Toxicon 1992, 30, 945–966. [Google Scholar] [CrossRef]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic snake venoms: Their functional activity, impact on snakebite victims and pharmaceutical promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef]
- Fontana, F.; Skinner, J. Treatise on the Venom of the Viper, on the American Poisons, and on the Cherry Laurel, and Some Other Vegetable poisons: To Which Are Annexed, Observations on the Primitive Structure of the Animal Body, Different Experiments on the Reproduction of the Nerves, and a Description of a New Canal of the Eye; Printed for John Cuthell: London, UK, 1795; Volume v.1. [Google Scholar]
- Lu, Q.; Clemetson, J.M.; Clemetson, K.J. Snake venoms and hemostasis. J. Thromb. Haemost. 2005, 3, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Anticoagulant proteins from snake venoms: Structure, function and mechanism. Biochem. J. 2006, 397, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Marsh, N.; Williams, V. Practical applications of snake venom toxins in haemostasis. Toxicon 2005, 45, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K. The Pro-Coagulant Fibrinogenolytic Serine Protease Isoenzymes Purified from Daboia russelii russelii Venom Coagulate the Blood through Factor V Activation: Role of Glycosylation on Enzymatic Activity. PLoS ONE 2014, 9, e86823. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.; Bon, C. Blood coagulation induced by the venom of Bothrops atrox. 2. Identification, purification, and properties of two factor X activators. Biochemistry 1987, 26, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-H.; Zhang, Y.; Wang, W.-Y.; Xiong, Y.-L.; Gao, R. Isolation and properties of a blood coagulation factor X activator from the venom of king cobra (Ophiophagus hannah). Toxicon 1995, 33, 1263–1276. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiong, Y.-L.; Bon, C. An activator of blood coagulation factor X from the venom of Bungarus fasciatus. Toxicon 1995, 33, 1277–1288. [Google Scholar] [CrossRef]
- Siigur, E.; Tonismagi, K.; Trummal, K.; Samel, M.; Vija, H.; Subbi, J.; Siigur, J. Factor X activator from Vipera lebetina snake venom, molecular characterization and substrate specificity. Biochim. Biophys. Acta 2001, 1568, 90–98. [Google Scholar] [CrossRef]
- Tans, G.; Rosing, J. Snake venom activators of factor X: An overview. Haemostasis 2001, 31, 225–233. [Google Scholar] [CrossRef]
- Bittenbinder, M.A.; Zdenek, C.N.; Op den Brouw, B.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib). Toxins 2018, 10, 516. [Google Scholar] [CrossRef]
- Kini, R.M. Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon 2005, 45, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.D.; Walker, F.J. Purification of a protein C activator from the venom of the southern copperhead snake (Agkistrodon contortrix contortrix). Biochemistry 1986, 25, 4175–4179. [Google Scholar] [CrossRef] [PubMed]
- Bakker, H.M.; Tans, G.; Yukelson, L.Y.; Janssen-Claessen, T.W.; Bertina, R.M.; Hemker, H.C.; Rosing, J. Protein C activation by an activator purified from the venom of Agkistrodon halys halys. Blood Coagul. Fibrinolysis 1993, 4, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Gempeler-Messina, P.M.; Volz, K.; Buhler, B.; Muller, C. Protein C activators from snake venoms and their diagnostic use. Haemostasis 2001, 31, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, C.; Teng, C.M. Purification and properties of the anticoagulant principle of Agkistrodon acutus venom. Biochim. Biophys. Acta 1972, 278, 155–162. [Google Scholar] [CrossRef]
- Ouyang, C.; Yang, F.Y. Purification and properties of the anticoagulant principle of Trimeresurus gramineus venom. Biochim. Biophys. Acta 1975, 386, 479–492. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tsai, I.H. Functional and sequence characterization of coagulation factor IX/factor X-binding protein from the venom of Echis carinatus leucogaster. Biochemistry 1996, 35, 5264–5271. [Google Scholar] [CrossRef]
- Atoda, H.; Yoshida, N.; Ishikawa, M.; Morita, T. Binding properties of the coagulation factor IX/factor X-binding protein isolated from the venom of Trimeresurus flavoviridis. Eur. J. Biochem. 1994, 224, 703–708. [Google Scholar] [CrossRef]
- Zingali, R.B.; Jandrot-Perrus, M.; Guillin, M.C.; Bon, C. Bothrojaracin, a new thrombin inhibitor isolated from Bothrops jararaca venom: Characterization and mechanism of thrombin inhibition. Biochemistry 1993, 32, 10794–10802. [Google Scholar] [CrossRef]
- Markland, F.S., Jr. Snake venom fibrinogenolytic and fibrinolytic enzymes: An updated inventory. Registry of Exogenous Hemostatic Factors of the Scientific and Standardization Committee of the International Society on Thrombosis and Haemostasis. Thromb. Haemost. 1998, 79, 668–674. [Google Scholar] [CrossRef]
- Siigur, E.; Siigur, J. Purification and characterization of lebetase, a fibrinolytic enzyme from Vipera lebetina (snake) venom. Biochim Biophys Acta 1991, 1074, 223–229. [Google Scholar] [CrossRef]
- Maruyama, M.; Sugiki, M.; Yoshida, E.; Shimaya, K.; Mihara, H. Broad substrate specificity of snake venom fibrinolytic enzymes: Possible role in haemorrhage. Toxicon 1992, 30, 1387–1397. [Google Scholar] [CrossRef]
- Maruyama, M.; Tanigawa, M.; Sugiki, M.; Yoshida, E.; Mihara, H. Purification and characterization of low molecular weight fibrinolytic/hemorrhagic enzymes from snake (Bothrops jararaca) venom. Enzym. Protein 1993, 47, 124–135. [Google Scholar]
- Kerkkamp, H.; Bagowski, C.; Kool, J.; van Soolingen, B.; Vonk, F.J.; Vlecken, D. Whole snake venoms: Cytotoxic, anti-metastatic and antiangiogenic properties. Toxicon 2018, 150, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, D.; Sarkar, A. Cytotoxic Effects of Snake Venoms. In Snake Venoms; Inagaki, H., Vogel, C.-W., Mukherjee, A.K., Rahmy, T.R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 327–342. [Google Scholar] [CrossRef]
- Chaim-Matyas, A.; Ovadia, M. Cytotoxic activity of various snake venoms on melanoma, B16F10 and chondrosarcoma. Life Sci. 1987, 40, 1601–1607. [Google Scholar] [CrossRef]
- Damico, D.C.S.; Nascimento, J.M.; Lomonte, B.; Ponce-Soto, L.A.; Joazeiro, P.P.; Novello, J.C.; Marangoni, S.; Collares-Buzato, C.B. Cytotoxicity of Lachesis muta muta snake (bushmaster) venom and its purified basic phospholipase A2 (LmTX-I) in cultured cells. Toxicon 2007, 49, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Brahmbhatt, K.; Bhatt, H.; Parmar, U. Therapeutic potential of snake venom in cancer therapy: Current perspectives. Asian Pac. J. Trop Biomed. 2013, 3, 156–162. [Google Scholar] [CrossRef]
- Shanbhag, V.K.L. Applications of snake venoms in treatment of cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 275–276. [Google Scholar] [CrossRef] [Green Version]
- Vonk, F.J.; Jackson, K.; Doley, R.; Madaras, F.; Mirtschin, P.J.; Vidal, N. Snake venom: From fieldwork to the clinic: Recent insights into snake biology, together with new technology allowing high-throughput screening of venom, bring new hope for drug discovery. Bioessays 2011, 33, 269–279. [Google Scholar] [CrossRef]
- Koh, D.C.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef]
- Stransky, S.; Costal-Oliveira, F.; Lopes-de-Souza, L.; Guerra-Duarte, C.; Chávez-Olórtegui, C.; Braga, V.M.M. In vitro assessment of cytotoxic activities of Lachesis muta muta snake venom. PLoS Negl. Trop. Dis. 2018, 12, e0006427. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.a.; Ownby, C.L. Skeletal muscle degeneration induced by venom phospholipases A2: Insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D.; Ownby, C.L. Myotoxic components of snake venoms: Their biochemical and biological activities. Pharmacol. Ther. 1990, 48, 223–236. [Google Scholar] [CrossRef]
- Silva, A.; Johnston, C.; Kuruppu, S.; Kneisz, D.; Maduwage, K.; Kleifeld, O.; Smith, A.I.; Siribaddana, S.; Buckley, N.A.; Hodgson, W.C.; et al. Clinical and Pharmacological Investigation of Myotoxicity in Sri Lankan Russell’s Viper (Daboia russelii) Envenoming. PLoS Negl. Trop. Dis. 2016, 10, e0005172. [Google Scholar] [CrossRef] [PubMed]
- Lin Shiau, S.Y.; Huang, M.C.; Lee, C.Y. Mechanism of action of cobra cardiotoxin in the skeletal muscle. J. Pharm. Exp. 1976, 196, 758–770. [Google Scholar]
- Cameron, D.L.; Tu, A.T. Chemical and functional homology of myotoxin a from prairie rattlesnake venom and crotamine from South American rattlesnake venom. Biochim. Biophys. Acta 1978, 532, 147–154. [Google Scholar] [CrossRef]
- Ferreira, B.L.; Santos, D.O.; Santos, A.L.d.; Rodrigues, C.R.; de Freitas, C.C.; Cabral, L.M.; Castro, H.C. Comparative Analysis of Viperidae Venoms Antibacterial Profile: A Short Communication for Proteomics. Evid. -Based Complement. Altern. Med. 2011, 2011, 4. [Google Scholar] [CrossRef]
- De Oliveira Junior, N.G.; e Silva Cardoso, M.H.; Franco, O.L. Snake venoms: Attractive antimicrobial proteinaceous compounds for therapeutic purposes. Cell Mol. Life Sci. 2013, 70, 4645–4658. [Google Scholar] [CrossRef]
- Charvat, R.A.; Strobel, R.M.; Pasternak, M.A.; Klass, S.M.; Rheubert, J.L. Analysis of snake venom composition and antimicrobial activity. Toxicon 2018, 150, 151–167. [Google Scholar] [CrossRef]
- Almeida, J.R.; Palacios, A.L.V.; Patiño, R.S.P.; Mendes, B.; Teixeira, C.A.S.; Gomes, P.; da Silva, S.L. Harnessing snake venom phospholipases A2 to novel approaches for overcoming antibiotic resistance. Drug Dev. Res. 2019, 80, 68–85. [Google Scholar] [CrossRef]
- Perumal Samy, R.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H.K. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.G.; Toyama, M.H.; Novello, J.C.; Beriam, L.O.; Marangoni, S. Structural and functional characterization of basic PLA2 isolated from Crotalus durissus terrificus venom. J. Protein Chem. 2002, 21, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Samy, R.P.; Gopalakrishnakone, P.; Stiles, B.G.; Girish, K.S.; Swamy, S.N.; Hemshekhar, M.; Tan, K.S.; Rowan, E.G.; Sethi, G.; Chow, V.T. Snake venom phospholipases A(2): A novel tool against bacterial diseases. Curr. Med. Chem. 2012, 19, 6150–6162. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Dantas, R.; Menezes, R.; Toyama, M.; Filho, E.; Oliveira, M.; Nogueira, N.; Oliveira, M.; Monteiro, H.; Martins, A. Antimicrobial activity of an L-amino acid oxidase isolated from Bothrops leucurus snake venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 614–622. [Google Scholar] [CrossRef]
- Izidoro, L.F.; Ribeiro, M.C.; Souza, G.R.; Sant’Ana, C.D.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Goulart, L.R.; Beleboni, R.O.; Nomizo, A.; Sampaio, S.V.; et al. Biochemical and functional characterization of an L-amino acid oxidase isolated from Bothrops pirajai snake venom. Bioorganic Med. Chem. 2006, 14, 7034–7043. [Google Scholar] [CrossRef]
- Samy, R.P.; Stiles, B.G.; Gopalakrishnakone, P.; Chow, V.T. Antimicrobial proteins from snake venoms: Direct bacterial damage and activation of innate immunity against Staphylococcus aureus skin infection. Curr. Med. Chem 2011, 18, 5104–5113. [Google Scholar] [CrossRef]
- Petricevich, V.L.; Mendonca, R.Z. Inhibitory potential of Crotalus durissus terrificus venom on measles virus growth. Toxicon 2003, 42, 143–153. [Google Scholar] [CrossRef]
- Muller, V.D.; Soares, R.O.; dos Santos, N.N., Jr.; Trabuco, A.C.; Cintra, A.C.; Figueiredo, L.T.; Caliri, A.; Sampaio, S.V.; Aquino, V.H. Phospholipase A2 isolated from the venom of Crotalus durissus terrificus inactivates dengue virus and other enveloped viruses by disrupting the viral envelope. PLoS ONE 2014, 9, e112351. [Google Scholar] [CrossRef]
- Borkow, G.; Ovadia, M. Selective lysis of virus-infected cells by cobra snake cytotoxins: A sendai virus, human erythrocytes, and cytotoxin model. Biochem. Biophys. Res. Commun. 1999, 264, 63–68. [Google Scholar] [CrossRef]
- Fenard, D.; Lambeau, G.; Valentin, E.; Lefebvre, J.C.; Lazdunski, M.; Doglio, A. Secreted phospholipases A(2), a new class of HIV inhibitors that block virus entry into host cells. J. Clin. Investig. 1999, 104, 611–618. [Google Scholar] [CrossRef]
- Alrajhi, A.A.; Almohaizeie, A. Snake venom preparation for drug-resistant human immunodeficiency virus. Ann. Saudi Med. 2008, 28, 292–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.J.; Wang, J.H.; Lee, W.H.; Wang, Q.; Liu, H.; Zheng, Y.T.; Zhang, Y. Molecular characterization of Trimeresurus stejnegeri venom L-amino acid oxidase with potential anti-HIV activity. Biochem. Biophys. Res. Commun. 2003, 309, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, J.F.; Pereira, C.M.; Bittar, C.; Batista, M.N.; Campos, G.R.F.; da Silva, S.; Cintra, A.C.O.; Zothner, C.; Harris, M.; Sampaio, S.V.; et al. Multiple effects of toxins isolated from Crotalus durissus terrificus on the hepatitis C virus life cycle. PLoS ONE 2017, 12, e0187857. [Google Scholar] [CrossRef] [PubMed]
- Magaldi, S.; Giron, M.E.; Aguilar, I.; Rodriguez-Acosta, A. Antifungal activity of Crotalus durissus cumanensis venom. Mycoses 2002, 45, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Yamane, E.S.; Bizerra, F.C.; Oliveira, E.B.; Moreira, J.T.; Rajabi, M.; Nunes, G.L.; de Souza, A.O.; da Silva, I.D.; Yamane, T.; Karpel, R.L.; et al. Unraveling the antifungal activity of a South American rattlesnake toxin crotamine. Biochimie 2013, 95, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Gomez, R.; Zerrouk, H.; Sebti, F.; Loyens, M.; Benslimane, A.; Ouaissi, M.A. Growth inhibition of Trypanosoma cruzi and Leishmania donovani infantum by different snake venoms: Preliminary identification of proteins from Cerastes cerastes venom which interact with the parasites. Toxicon 1994, 32, 875–882. [Google Scholar] [CrossRef]
- Ciscotto, P.; Machado de Avila, R.A.; Coelho, E.A.; Oliveira, J.; Diniz, C.G.; Farias, L.M.; de Carvalho, M.A.; Maria, W.S.; Sanchez, E.F.; Borges, A.; et al. Antigenic, microbicidal and antiparasitic properties of an l-amino acid oxidase isolated from Bothrops jararaca snake venom. Toxicon 2009, 53, 330–341. [Google Scholar] [CrossRef]
- Goncalves, A.R.; Soares, M.J.; de Souza, W.; DaMatta, R.A.; Alves, E.W. Ultrastructural alterations and growth inhibition of Trypanosoma cruzi and Leishmania major induced by Bothrops jararaca venom. Parasitol. Res. 2002, 88, 598–602. [Google Scholar] [CrossRef]
- Franca, S.C.; Kashima, S.; Roberto, P.G.; Marins, M.; Ticli, F.K.; Pereira, J.O.; Astolfi-Filho, S.; Stabeli, R.G.; Magro, A.J.; Fontes, M.R.; et al. Molecular approaches for structural characterization of Bothrops L-amino acid oxidases with antiprotozoal activity: cDNA cloning, comparative sequence analysis, and molecular modeling. Biochem. Biophys. Res. Commun. 2007, 355, 302–306. [Google Scholar] [CrossRef]
- Costa Torres, A.F.; Dantas, R.T.; Toyama, M.H.; Filho, E.D.; Zara, F.J.; Rodrigues de Queiroz, M.G.; Pinto Nogueira, N.A.; Rosa de Oliveira, M.; de Oliveira Toyama, D.; Monteiro, H.S.A.; et al. Antibacterial and antiparasitic effects of Bothrops marajoensis venom and its fractions: Phospholipase A2 and l-amino acid oxidase. Toxicon 2010, 55, 795–804. [Google Scholar] [CrossRef]
- Gomes, A. Snake Venom—An Anti Arthritis Natural Product. Al Ameen J. Med. Sci. 2010, 3, 179. [Google Scholar]
- Crow, J.M. Venomous drugs: Captopril. New Sci. 2012, 214, 35. [Google Scholar] [CrossRef]
- Peng, H.; Carretero, O.A.; Vuljaj, N.; Liao, T.D.; Motivala, A.; Peterson, E.L.; Rhaleb, N.E. Angiotensin-converting enzyme inhibitors: A new mechanism of action. Circulation 2005, 112, 2436–2445. [Google Scholar] [CrossRef] [PubMed]
- Stepensky, D. Pharmacokinetics of Toxin-Derived Peptide Drugs. Toxins 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.G.; Vane, J.R. The discovery of captopril. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 788–789. [Google Scholar] [CrossRef]
- Koh, C.Y.; Kini, R.M. From snake venom toxins to therapeutics--cardiovascular examples. Toxicon 2012, 59, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Lazarovici, P.; Marcinkiewicz, C.; Lelkes, P.I. From Snake Venom’s Disintegrins and C-Type Lectins to Anti-Platelet Drugs. Toxins 2019, 11, 303. [Google Scholar] [CrossRef]
- Egbertson, M.S.; Chang, C.T.; Duggan, M.E.; Gould, R.J.; Halczenko, W.; Hartman, G.D.; Laswell, W.L.; Lynch, J.J., Jr.; Lynch, R.J.; Manno, P.D.; et al. Non-peptide fibrinogen receptor antagonists. 2. Optimization of a tyrosine template as a mimic for Arg-Gly-Asp. J. Med. Chem. 1994, 37, 2537–2551. [Google Scholar] [CrossRef]
- Chen, Y.; Pitzenberger, S.M.; Garsky, V.M.; Lumma, P.K.; Sanyal, G.; Baum, J. Proton NMR assignments and secondary structure of the snake venom protein echistatin. Biochemistry 1991, 30, 11625–11636. [Google Scholar] [CrossRef]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S. Trigramin. A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein IIb-IIIa complex. J. Biol Chem. 1987, 262, 16157–16163. [Google Scholar]
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. Echistatin. A potent platelet aggregation inhibitor from the venom of the viper, Echis carinatus. J. Biol. Chem. 1988, 263, 19827–19832. [Google Scholar]
- Curran, M.P.; Keating, G.M. Eptifibatide: a review of its use in patients with acute coronary syndromes and/or undergoing percutaneous coronary intervention. Drugs 2005, 65, 2009–2035. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.C.; Tcheng, J.E. Eptifibatide: a potent inhibitor of the platelet receptor integrin glycoprotein IIb/IIIa. Expert Opin. Pharmacother. 2002, 3, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.T.; Stafford, A.R.; Leslie, B.A.; Kim, P.Y.; Fredenburgh, J.C.; Weitz, J.I. Batroxobin binds fibrin with higher affinity and promotes clot expansion to a greater extent than thrombin. J. Biol. Chem. 2013, 288, 16862–16871. [Google Scholar] [CrossRef] [PubMed]
- Lodha, A.; Kamaluddeen, M.; Akierman, A.; Amin, H. Role of hemocoagulase in pulmonary hemorrhage in preterm infants: a systematic review. Indian J. Pediatrics 2011, 78, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-J.; Brighton, T.A. Ximelagatran: Direct thrombin inhibitor. Vasc. Health Risk Manag. 2006, 2, 49–58. [Google Scholar] [CrossRef]
- Han, S.M.; Weaver, F.A.; Comerota, A.J.; Perler, B.A.; Joing, M. Efficacy and safety of alfimeprase in patients with acute peripheral arterial occlusion (PAO). J. Vasc. Surg. 2010, 51, 600–609. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, E.E.; Andrews, R.K. The cut of the clot(h): Snake venom fibrinogenases as therapeutic agents. J. Thromb. Haemost. 2008, 6, 1360–1362. [Google Scholar] [CrossRef]
- Kunalan, S.; Othman, I.; Syed Hassan, S.; Hodgson, W.C. Proteomic Characterization of Two Medically Important Malaysian Snake Venoms, Calloselasma rhodostoma (Malayan Pit Viper) and Ophiophagus hannah (King Cobra). Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Johnstone, I.B.; Martin, C.A. Comparative effects of the human protein C activator, Protac, on the activated partial thromboplastin clotting times of plasmas, with special reference to the dog. Can. J. Vet. Res. Rev. Can. De Rech. Vet. 2000, 64, 117–122. [Google Scholar]
- Fukuda, K.; Doggett, T.; Laurenzi, I.J.; Liddington, R.C.; Diacovo, T.G. The snake venom protein botrocetin acts as a biological brace to promote dysfunctional platelet aggregation. Nat. Struct. Mol. Biol. 2005, 12, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, D.; Ben Ammar, Y.; Miyata, T.; Takeda, S. Structural basis of coagulation factor V recognition for cleavage by RVV-V. Febs Lett. 2011, 585, 3020–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suntravat, M.; Yusuksawad, M.; Sereemaspun, A.; Pérez, J.C.; Nuchprayoon, I. Effect of purified Russell’s viper venom-factor X activator (RVV-X) on renal hemodynamics, renal functions, and coagulopathy in rats. Toxicon 2011, 58, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Lange, U.; Olschewski, A.; Nowak, G.; Bucha, E. Ecarin chromogenic assay: An innovative test for quantitative determination of direct thrombin inhibitors in plasma. Hamostaseologie 2005, 25, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Lövgren, A. Recombinant snake venom prothrombin activators. Bioengineered 2013, 4, 153–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Target Microbe | Factor (Protein/Peptide) | Source (Snake Specie) |
|---|---|---|---|
| Antibacterial | Gram-positive and gram-negative bacteria | L-amino acid oxidase | Bothrop alternatus, Bothrop pirajai, Bothrop asper, Bothrop leucurus, Crotalus adamanteus, Daboia russelli russelli, Ophiophagus hannah and Pseudechis australis |
| Phospholipase (PLA2) | Bothrop asper, Bothrop jararacussu, Bothrop pirajai, Bothrop moojeni, Bungarus faciatus and Crotalus durissus terrificus | ||
| Antiviral | Measles, yellow fever and dengue viruses | Crotoxin and PLA2 | Crotalus durissus terrificus |
| Sendai virus | Cytotoxins | Naja nigricollis | |
| Human Immunodeficiency Virus (HIV) | L-amino acid oxidase | Trimeresurus stejnegeri, Bungarus candidus, Naja naja and Naja kaouthia | |
| Hepatitis C virus (HCV) | Crotoxin, PLA2 and crotapotin | Crotalus durissus terrificus | |
| Antifungal | Candida parapsilosis and Sporothrix schenckii | Metalloproteinases and PLA2 | Crotalus durissus cumanensis |
| Crotamine | Crotalus durissus terrificus | ||
| Antiparasitic | Leishmania species, Plasmodium falciparum and Trypanosoma cruzi | L-amino-acid oxidases | Brothrops snakes |
| Stage | Protein/Peptide | Lead Source | Pharmacology | Indication |
|---|---|---|---|---|
| FDA Approved | Captopril | Bothrops jararaca | Inhibiting angiotensin-converting enzyme. | Hypertension |
| Aggrastat (Tirofiban) | Echis carinatus | Glycoprotein IIb/IIIa inhibitors. | Heart attack | |
| Integrilin (Eptifibatide) | Sistrurus miliarus barbouri | Glycoprotein (GP) IIb/IIIa inhibitors. | Acute coronary syndrome | |
| Defibrase/Reptilase (Batroxobin) | Bothrops atrox & B. moojeni | Converts fibrinogen into fibrin. | Stroke, pulmonary embolism, deep vein thrombosis and myocardial infarction | |
| Hemocoagulase | Bothrops atrox | Catalyzes the coagulation of the blood. | Plastic surgery, abdominal surgery, and human vitrectomy | |
| Exanta (Ximelagatran) | Cobra venom | Direct thrombin inhibitors. | Thromboembolic complications of atrial fibrillation | |
| In clinical trials | Alfimeprase | Agkistrodon contortrix | Thrombolytic activity. | Acute peripheral arterial occlusion |
| Viprinex (Ancrod) | Agkistrodon rhodostoma | Defibrinogenating agent. | Acute ischemic stroke |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed Abd El-Aziz, T.; Soares, A.G.; Stockand, J.D. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins 2019, 11, 564. https://doi.org/10.3390/toxins11100564
Mohamed Abd El-Aziz T, Soares AG, Stockand JD. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins. 2019; 11(10):564. https://doi.org/10.3390/toxins11100564
Chicago/Turabian StyleMohamed Abd El-Aziz, Tarek, Antonio Garcia Soares, and James D. Stockand. 2019. "Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving" Toxins 11, no. 10: 564. https://doi.org/10.3390/toxins11100564
APA StyleMohamed Abd El-Aziz, T., Soares, A. G., & Stockand, J. D. (2019). Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins, 11(10), 564. https://doi.org/10.3390/toxins11100564