Metabolome Variation between Strains of Microcystis aeruginosa by Untargeted Mass Spectrometry


Abstract
:1. Introduction
2. Results
2.1. Identification of Cyanobacteria Metabolites
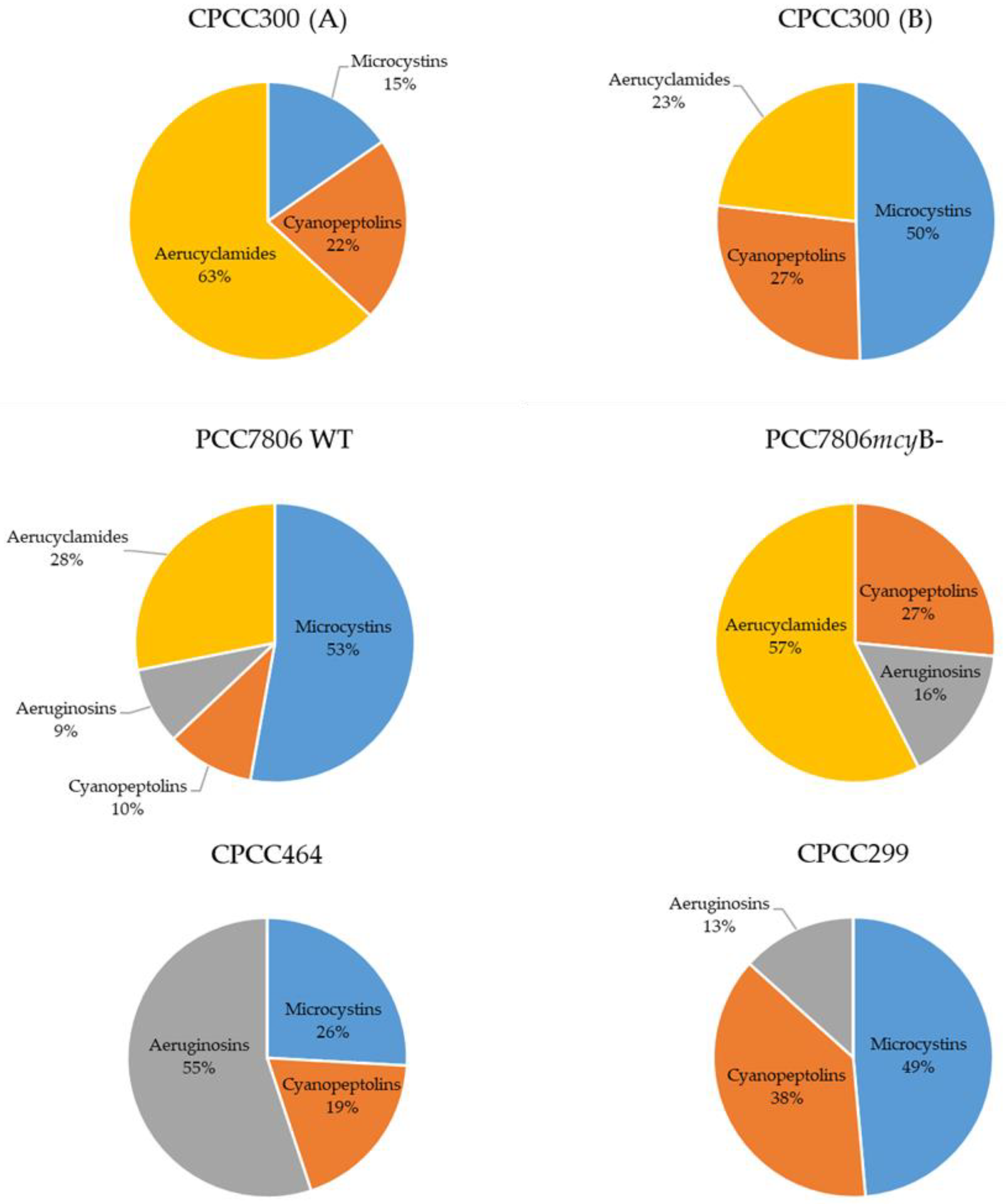
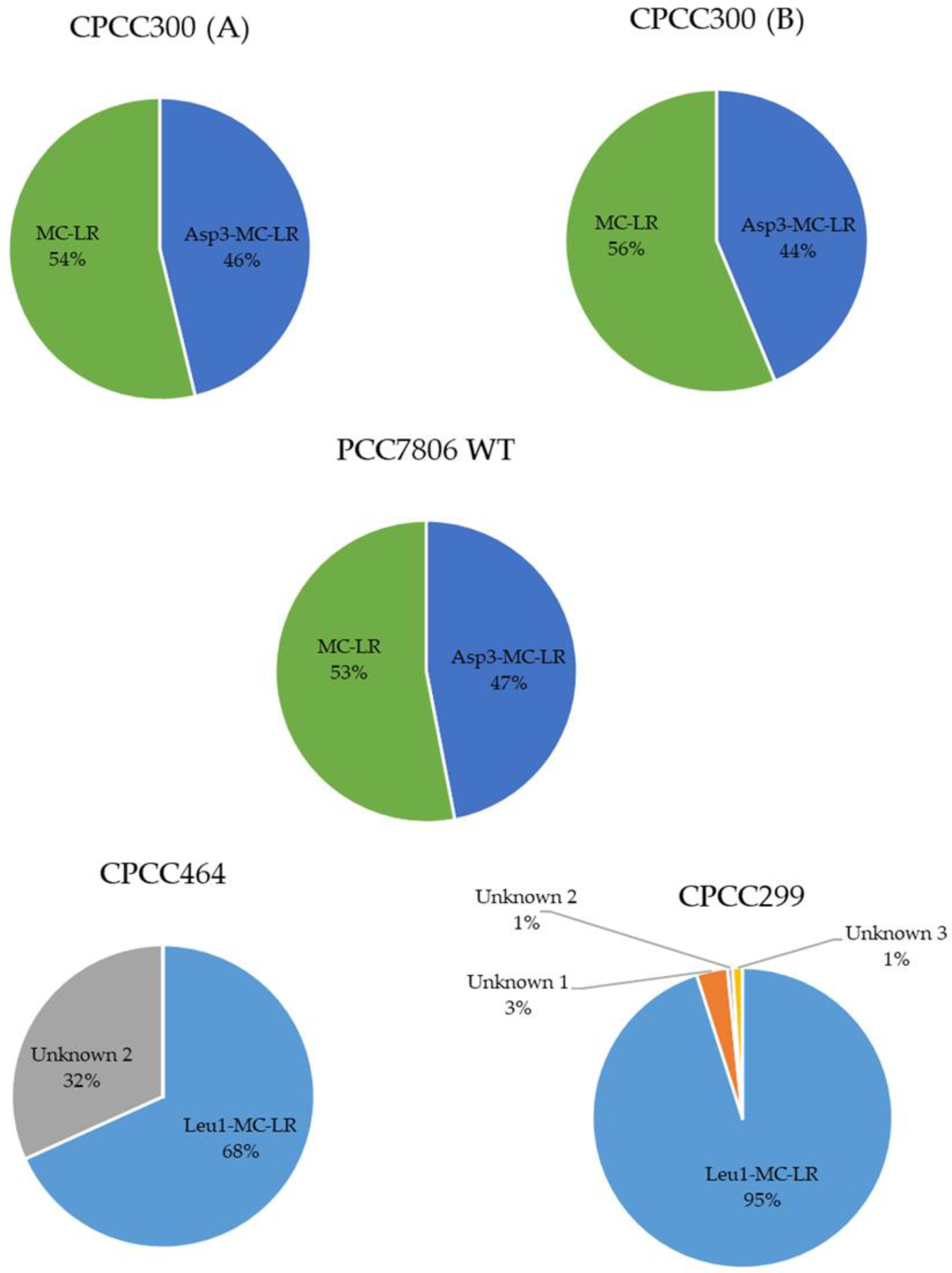
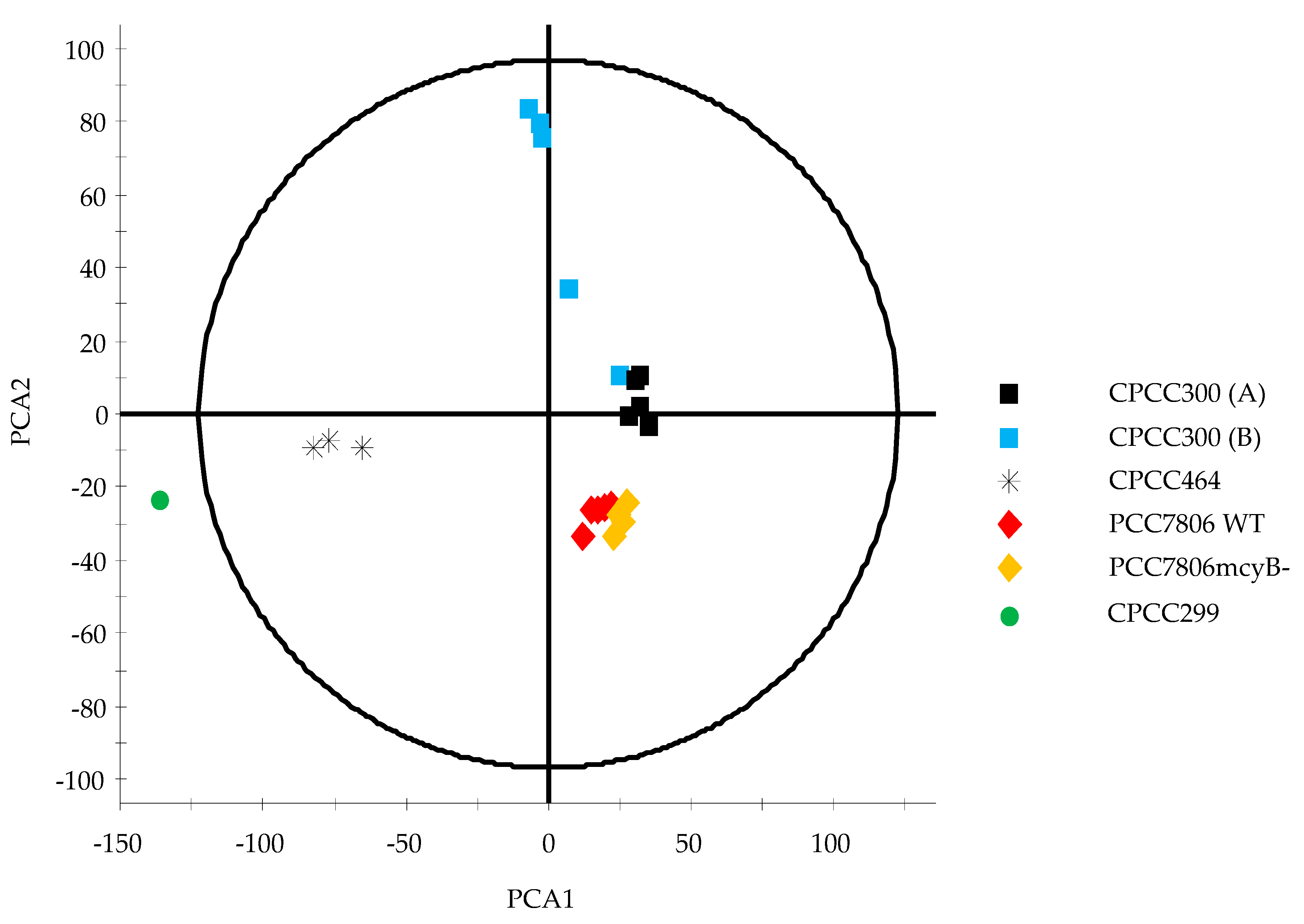
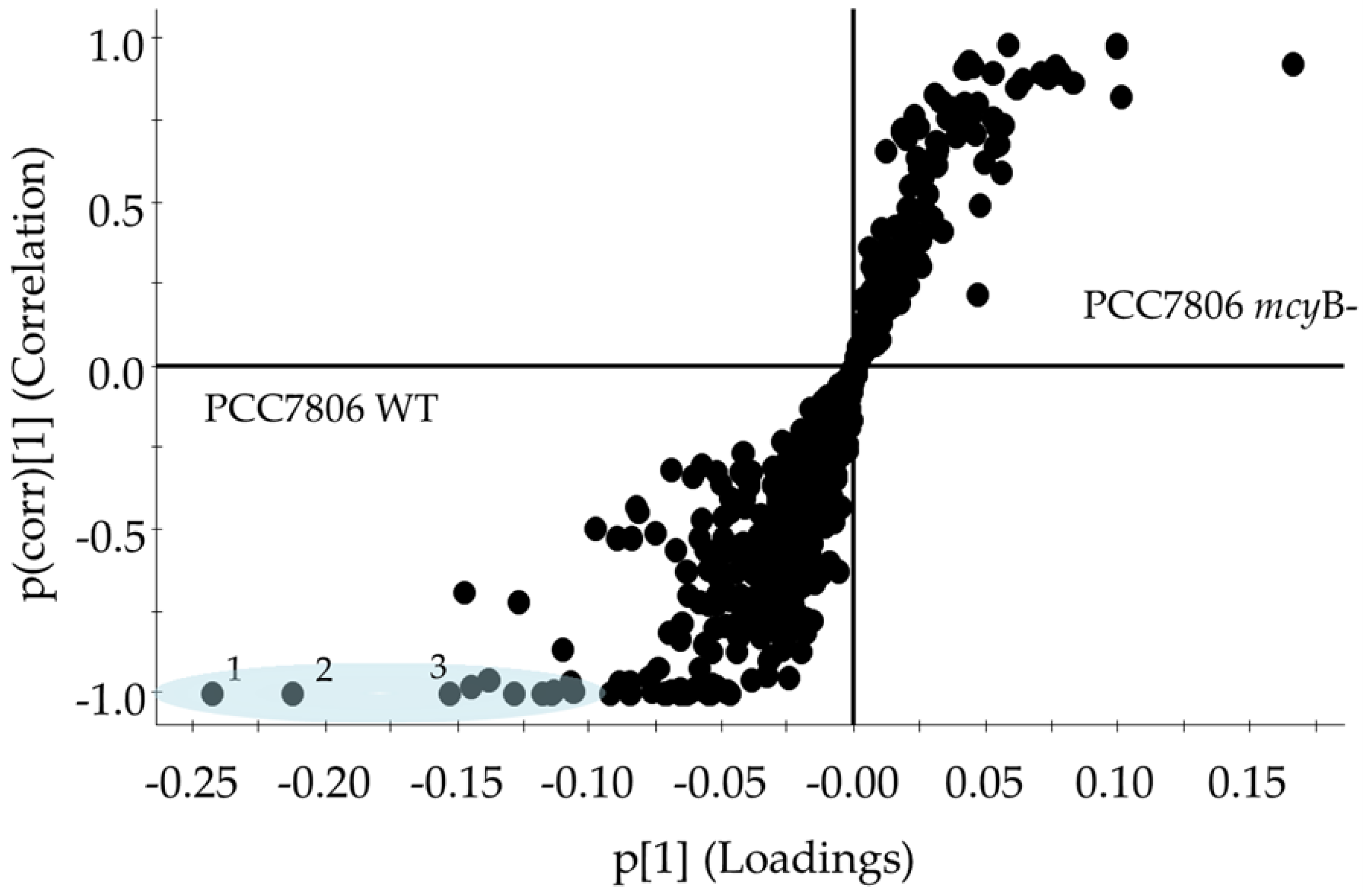
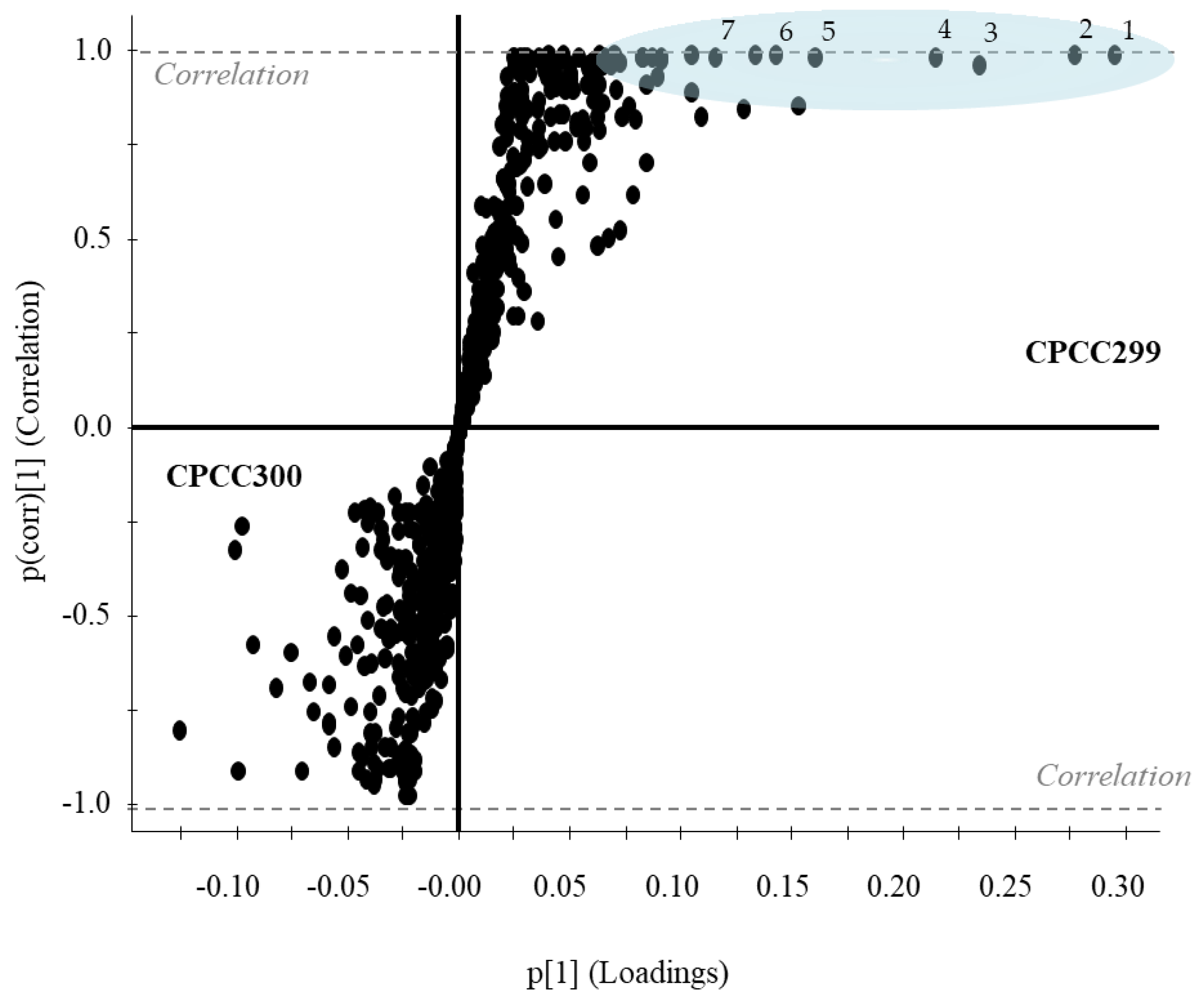
2.2. Metabolomes of M. aeruginosa Strains
3. Discussion
3.1. Metabolome Diversity in M. aeruginosa
3.2. Identification of Metabolites
4. Conclusions
5. Material and Methods
5.1. Culturing Techniques
5.2. Extraction
5.3. LC-MS Analyses
5.4. Pattern Recognition Statistical Analyses
5.5. Metabolite Identification
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
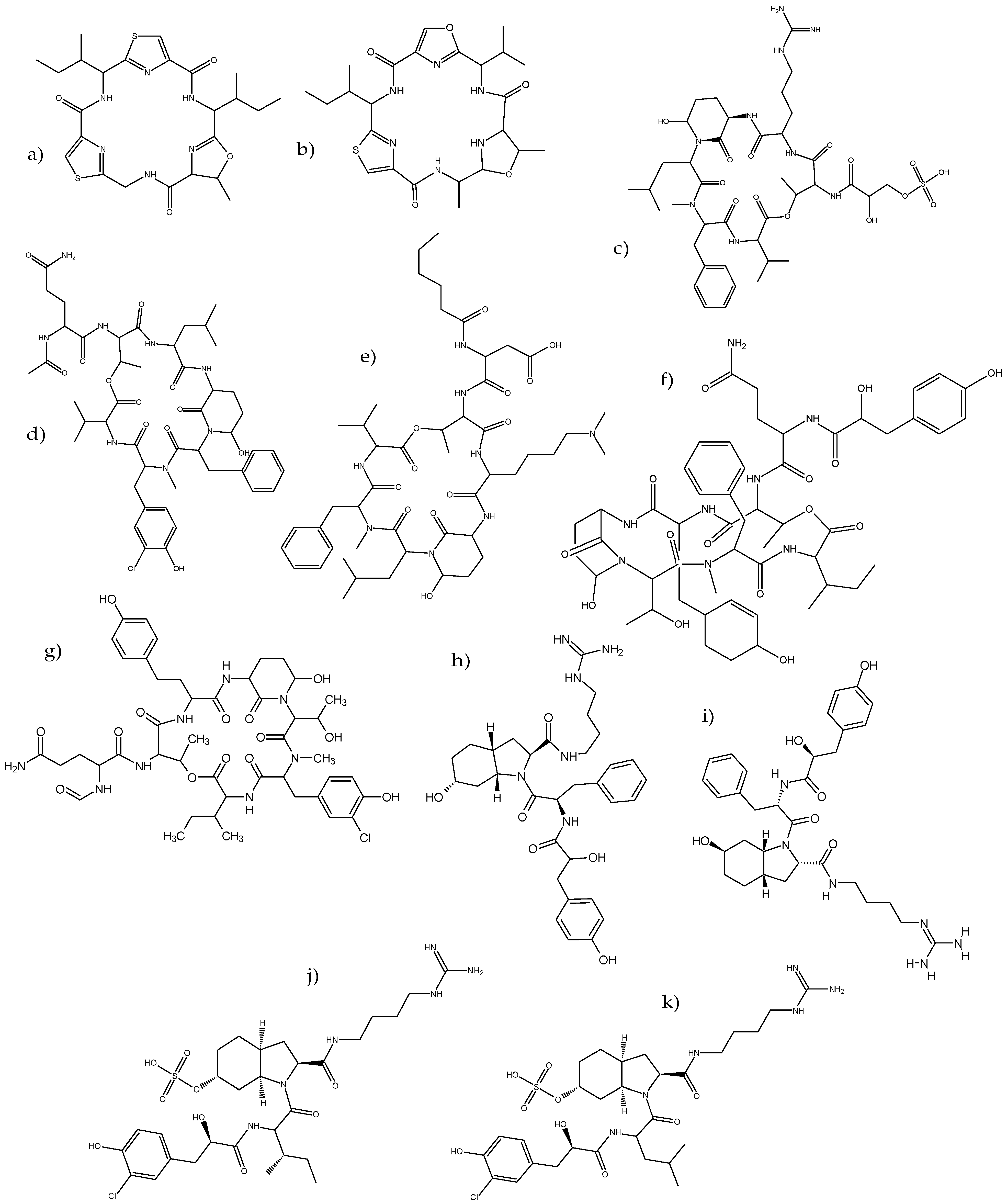{kind=link}
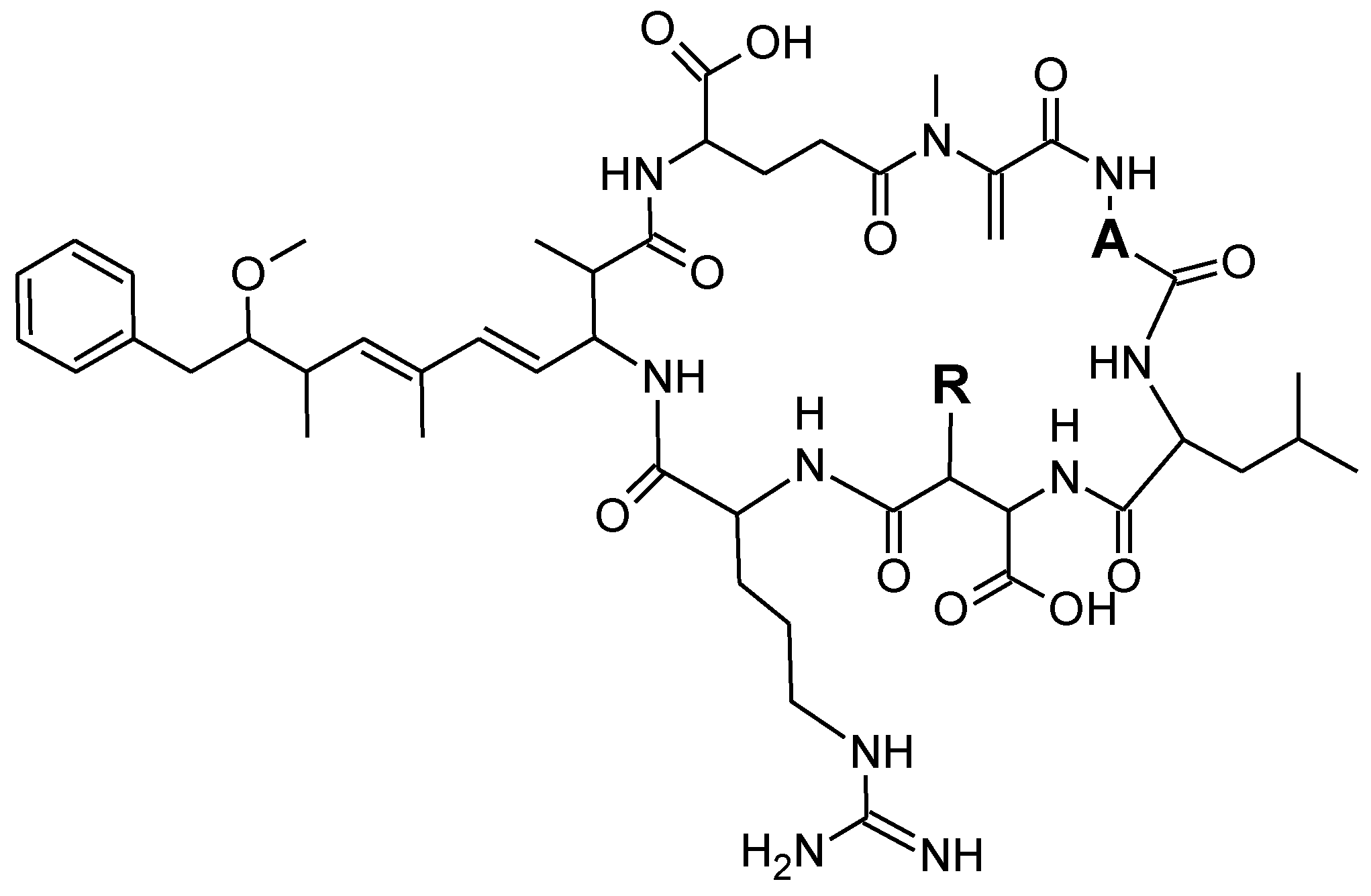
| Metabolites | A | R |
|---|---|---|
| MC-LR | D-Ala | CH3 |
| [Leu1]-MC-LR | D-Leu | CH3 |
| [Asp3]-MC-LR | D-Ala | H |

References
- Michalak, A.M.; Anderson, E.J.; Beletsky, D.; Boland, S.; Bosch, N.S.; Bridgeman, T.B.; Chaffin, J.D.; Cho, K.; Confesor, R.; Daloglu, I.; et al. Record-setting algal bloom in Lake Erie caused by agricultural and meteorological trends consistent with expected future conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6448–6452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falconer, I.R. Cyanobacterial Toxins of Drinking Water Supplies: Cylindrospermopsins and Microcystins; CRC Press: New York, NY, USA, 2004. [Google Scholar]
- Downing, J.A.; Watson, S.B.; McCauley, E. Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci. 2001, 58, 1905–1908. [Google Scholar] [CrossRef]
- Beaulieu, M.; Pick, F.R.; Gregory-Eaves, I. Nutrients and water temperature are significant predictors of cyanobacterial biomass in a 1147 lakes dataset. Limnol. Oceanogr. 2013, 58, 1736–1746. [Google Scholar] [CrossRef]
- Harris, T.D.; Smith, V.H. Do persistent organic pollutants stimulate cyanobacterial blooms? Inland Waters 2016, 6, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Pick, F.R. Blooming algae: A Canadian perspective on the rise of toxic cyanobacteria. Can. J. Fish. Aquat. Sci. 2016, 73, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Brooks, B.W.; Lazorchak, J.M.; Howard, M.D.A.; Johnson, M.V.V.; Morton, S.L.; Perkins, D.A.K.; Reavie, E.D.; Scott, G.I.; Smith, S.A.; Steevens, J.A. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems? Environ. Toxicol. Chem. 2016, 35, 6–13. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; World Health Organization, E&FN Spon: London, UK, 1999. [Google Scholar]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Pouria, S.; De Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.S.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.F.; Reichmuth, C.; Rouse, A.A.; Libby, L.A.; Dennison, S.E.; Carmichael, O.T.; Kruse-Elliott, K.T.; Bloom, J.; Singh, B.; Fravel, V.A.; et al. Algal toxin impairs sea lion memory and hippocampal connectivity, with implications for strandings. Science 2015, 350, 1445–1447. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; World Health Organization, E&FN Spon: London, UK, 1999. [Google Scholar]
- WHO. Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef] [Green Version]
- Dawson, R.M. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Rao, P.V.L. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Hollister, J.W.; Kreakie, B.J. Associations between chlorophyll a and various microcystin-LR health advisory concentrations. F1000Research 2016, 5, 151. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3: Tables of Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 526–537. [Google Scholar]
- Lazar, S.; Egoz, I.; Brandeis, R.; Chapman, S.; Bloch-Shilderman, E.; Grauer, E. Propagation of damage in the rat brain following sarin exposure: Differential progression of early processes. Toxicol. Appl. Pharmacol. 2016, 310, 87–97. [Google Scholar] [CrossRef]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Heal. Part B Crit. Rev. 2005, 8, 1–37. [Google Scholar] [CrossRef]
- Guyot, M.; Doré, J.C.; Devillers, J. Typology of secondary cyanobacterial metabolites from minimum spanning tree analysis. SAR QSAR Environ. Res. 2004, 15, 101–114. [Google Scholar] [CrossRef]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes Coding for Hepatotoxic Heptapeptides (Microcystins) in the Cyanobacterium Anabaena Strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Welker, M.; Von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Sinha, R.P. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Klassen, A.; Faccio, A.T.; Canuto, G.A.B.; da Cruz, P.L.R.; Ribeiro, H.C.; Tavares, M.F.M.; Sussulini, A. Metabolomics: Definitions and significance in systems biology. In Metabolomics: From Fundamentals to Clinical Applications; Springer: Basel, Switzerland, 2017. [Google Scholar]
- Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L.; Miller, T.R. Variable cyanobacterial toxin and metabolite profiles across six eutrophic lakes of differing physiochemical characteristics. Toxins 2017, 9, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayad, P.B.; Roy-Lachapelle, A.; Duy, S.V.; Prévost, M.; Sauvé, S. On-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for the analysis of cyanotoxins in algal blooms. Toxicon 2015, 108, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Roy-Lachapelle, A.; Fayad, P.B.; Sinotte, M.; Deblois, C.; Sauvé, S. Total microcystins analysis in water using laser diode thermal desorption-atmospheric pressure chemical ionization-tandem mass spectrometry. Anal. Chim. Acta 2014, 820, 76–83. [Google Scholar] [CrossRef]
- Sanz, M.; Andreote, A.P.D.; Fiore, M.F.; Dörr, F.A.; Pinto, E. Structural characterization of new peptide variants produced by cyanobacteria from the Brazilian Atlantic Coastal Forest using liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. Mar. Drugs 2015, 13, 3892–3919. [Google Scholar] [CrossRef]
- León, C.; Peñuela, G.A. Detected cyanotoxins by UHPLC MS/MS technique in tropical reservoirs of northeastern Colombia. Toxicon 2019, 167, 38–48. [Google Scholar] [CrossRef]
- Haddad, S.P.; Bobbitt, J.M.; Taylor, R.B.; Lovin, L.M.; Conkle, J.L.; Chambliss, C.K.; Brooks, B.W. Determination of microcystins, nodularin, anatoxin-a, cylindrospermopsin, and saxitoxin in water and fish tissue using isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1599, 66–74. [Google Scholar] [CrossRef]
- León, C.; Boix, C.; Beltrán, E.; Peñuela, G.; López, F.; Sancho, J.V.; Hernández, F. Study of cyanotoxin degradation and evaluation of their transformation products in surface waters by LC-QTOF MS. Chemosphere 2019, 229, 538–548. [Google Scholar] [CrossRef]
- Ortiz, X.; Korenkova, E.; Jobst, K.J.; MacPherson, K.A.; Reiner, E.J. A high throughput targeted and non-targeted method for the analysis of microcystins and anatoxin-A using on-line solid phase extraction coupled to liquid chromatography–quadrupole time-of-flight high resolution mass spectrometry. Anal. Bioanal. Chem. 2017, 409, 4959–4969. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.K.; Ravindra, R.; Van Wagoner, R.M.; Wright, J.L.C. Metabolomics-Guided Discovery of Microginin Peptides from Cultures of the Cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2018, 81, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Foss, A.J.; Miles, C.O.; Özen, M.; Demir, N.; Balcı, M.; Beach, D.G. Comprehensive multi-technique approach reveals the high diversity of microcystins in field collections and an associated isolate of Microcystis aeruginosa from a Turkish lake. Toxicon 2019, 167, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Okita, Y.; Matsuda, H.; Okino, T.; Murakami, M. Aeruginosins, protease inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron 1999, 55, 10971–10988. [Google Scholar] [CrossRef]
- Ishida, K.; Welker, M.; Christiansen, G.; Cadel-Six, S.; Bouchier, C.; Dittmann, E.; Hertweck, C.; De Marsac, N.T. Plasticity and evolution of aeruginosin biosynthesis in cyanobacteria. Appl. Environ. Microbiol. 2009, 75, 2017–2026. [Google Scholar] [CrossRef] [Green Version]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S. Determination of BMAA and three alkaloid cyanotoxins in lake water using dansyl chloride derivatization and high-resolution mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 5487–5501. [Google Scholar] [CrossRef]
- Leikoski, N.; Fewer, D.P.; Sivonen, K. Widespread occurrence and lateral transfer of the cyanobactin biosynthesis gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2009, 75, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Briand, E.; Bormans, M.; Gugger, M.; Dorrestein, P.C.; Gerwick, W.H. Changes in secondary metabolic profiles of Microcystis aeruginosa strains in response to intraspecific interactions. Environ. Microbiol. 2016, 18, 384–400. [Google Scholar] [CrossRef] [Green Version]
- Welker, M.; Maršálek, B.; Šejnohová, L.; von Döhren, H. Detection and identification of oligopeptides in Microcystis (cyanobacteria) colonies: Toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar] [CrossRef]
- Kultschar, B.; Dudley, E.; Wilson, S.; Llewellyn, C.A. Intracellular and extracellular metabolites from the cyanobacterium chlorogloeopsis fritschii, pcc 6912, during 48 hours of uv-b exposure. Metabolites 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Namikoshi, M.; Brittain, S.M.; Carmichael, W.W.; Murphy, T. [D-Leu1] microcystin-LR, a new microcystin isolated from waterbloom in a Canadian prairie lake. Toxicon 2001, 39, 855–862. [Google Scholar] [CrossRef]
- Pinto, R.C. Chemometrics Methods and Strategies in Metabolomics. In Metabolomics: From Fundamentals to Clinical Applications; Springer: Basel, Switzerland, 2017; pp. 163–190. [Google Scholar]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. A conversation on data mining strategies in LC-MS untargeted metabolomics: Pre-processing and pre-treatment steps. Metabolites 2016, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Sivonen, K.; Leikoski, N.; Fewer, D.P.; Jokela, J. Cyanobactins-ribosomal cyclic peptides produced by cyanobacteria. Appl. Microbiol. Biotechnol. 2010, 86, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Oberer, L.; Ino, T.; König, W.A.; Buschdtt, M.; Weckesser, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystins sp. PCC 7806. J. Antibiot. (Tokyo) 1993, 46, 1550–1556. [Google Scholar] [CrossRef] [Green Version]
- Leblanc Renaud, S. Microcystin Production and the Dominance of Toxigenic Strains of Cyanobacteria in Lake and Cultures Studies. Ph.D. Thesis, University of Ottawa, Ottawa, ON, Canada, 2009. [Google Scholar]
- Zhang, H.; Liu, Y.; Nie, X.; Liu, L.; Hua, Q.; Zhao, G.P.; Yang, C. The cyanobacterial ornithine-ammonia cycle involves an arginine dihydrolase article. Nat. Chem. Biol. 2018, 14, 575. [Google Scholar] [CrossRef]
- Portmann, C.; Blom, J.F.; Gademann, K.; Jüttner, F. Aerucyclamides A and B: Isolation and synthesis of toxic ribosomal heterocyclic peptides from the cyanobacterium Microcystis aeruginosa PCC7806. J. Nat. Prod. 2008, 71, 1193–1195. [Google Scholar] [CrossRef]
- Faltermann, S.; Zucchi, S.; Kohler, E.; Blom, J.F.; Pernthaler, J.; Fent, K. Molecular effects of the cyanobacterial toxin cyanopeptolin (CP1020) occurring in algal blooms: Global transcriptome analysis in zebrafish embryos. Aquat. Toxicol. 2014, 149, 33–39. [Google Scholar] [CrossRef]
- Pereira, D.A.; Giani, A. Cell density-dependent oligopeptide production in cyanobacterial strains. FEMS Microbiol. Ecol. 2014, 88, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Jüttner, F.; Watson, S.B.; von Elert, E.; Köster, O. β-Cyclocitral, a Grazer Defence Signal Unique to the Cyanobacterium Microcystis. J. Chem. Ecol. 2010, 36, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morán, X.A.G.; Gasol, J.M.; Arin, L.; Estrada, M. A comparison between glass fiber and membrane filters for the estimation of phytoplankton POC and DOC production. Mar. Ecol. Prog. Ser. 1999, 187, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.A. Analytical techniques for the detection of α-amino-β-methylaminopropionic acid. Analyst 2012, 137, 1991–2005. [Google Scholar] [CrossRef]
- Rohrlack, T.; Edvardsen, B.; Skulberg, R.; Halstvedt, C.B.; Utkilen, H.C.; Ptacnik, R.; Skulberg, O.M. Oligopeptide chemotypes of the toxic freshwater cyanobacterium Planktothrix can form subpopulations with dissimilar ecological traits. Limnol. Oceanogr. 2008, 53, 1279–1293. [Google Scholar] [CrossRef]
- Anderson, R.A. Algal Culturing Techniques, 1st ed.; Elsevier Academic Press: New York, NY, USA, 2005. [Google Scholar]
- Aranda-Rodriguez, R.; Tillmanns, A.; Benoit, F.M.; Pick, F.R.; Harvie, J.; Solenaia, L. Pressurized liquid extraction of toxins from cyanobacterial cells. Environ. Toxicol. 2005, 20, 390–396. [Google Scholar] [CrossRef]
- Ortelli, D.; Edder, P.; Cognard, E.; Jan, P. Fast screening and quantitation of microcystins in microalgae dietary supplement products and water by liquid chromatography coupled to time of flight mass spectrometry. Anal. Chim. Acta 2008, 617, 230–237. [Google Scholar] [CrossRef]






| Oligopeptide Families | Metabolites | Molecular Formula | Precursor Ion | Major Fragments | RT (min) | CPCC464 | CPCC299 | CPCC300 | PCC7806 | PCC7806 mcyB- |
|---|---|---|---|---|---|---|---|---|---|---|
| Microcystins | [Asp3]-MC-LR * | C48H72N10O12 | 981.5220 | 135.0820, 502.2665, 847.4663 | 2.84 | X | X | |||
| [Leu1]-MC-LR | C52H80N10O12 | 1037.6033 | 135.0808, 169.1345, 530.2979, 903.5250 | 3.06 | X | X | ||||
| MC-LR * | C49H74N10O12 | 995.5287 | 135.0784, 213.0799, 509.2578, 861.4597 | 2.84 | X | X | ||||
| Microcystin (Unknown 1) | N/A** | 1031.4791 | 135.0808, 282.1383 | 3.18 | X | |||||
| Microcystin (Unknown 2) | 1055.5521 | 135.0808, 539.2790 | 2.98 | X | X | |||||
| Microcystin (Unknown 3) | 1009.4980 | 135.0975, 282.1452, 404.1939 | 3.24 | X | ||||||
| Cyanobactins | Aerucyclamide B | C24H32N6O4S2 | 533.1835 | 140.0681, 337.2317, 437.1928 | 4.59 | X | X | X | ||
| Aerucyclamide C | C24H32N6O5S | 517.2213 | 140.0657, 425.2299, 437.1971 | 4.47 | X | |||||
| Cyanopeptolins | Cyanopeptolin 911 (CPT911) | C39H61N9O14 | 912.4331 | 157.0907, 199.1181 | 2.13 | X | X | |||
| Cyanopeptolin 911 (isomer) | 2.25 | X | X | |||||||
| Cyanopeptolin 954 (CPT954) | C46H63ClN8O12 | 955.4891 | 222.1122, 486.2253, 915.4907 | 3.62 | X | |||||
| Cyanopeptolin D (CPTD) | C48H76N8O12 | 957.5405 | 222.1091, 261.1078, 470.2648 | 3.13 | X | X | X | |||
| Aeruginopeptin 228B | C52H72N8O15 | 1049.4680 | 129.0720, 222.1122, 404.1939, 522.2386, 888.4375 | 2.79 | X | |||||
| Anabaenopeptilide 90B | C45H61ClN8O14 | 973.4313 | 140.0657, 181.1215, 222.1122, 973.4313 | 3.47 | X | X | ||||
| Aeruginosins | Aeruginosin KT608A/KT608B/Microcin SF608 | C32H44N6O6 | 609.2664 | 149.0242, 178.1235, 242.2856, 301.1415, 515.3187 | 5.53 | X | ||||
| Aeruginosin KT608A/KT608B/Microcin SF608 | 609.3372 | 113.9631, 178.1225, 222.1122, 260.0569 | 1.65 | X | ||||||
| Aeruginosin 98A/DA688 | C29H45ClN6O9S | 689.2940 | 609.3372, 280.2124 | 1.46 | X | X | ||||
| Aeruginosin 98A/DA688 | 689.2833 | 113.9631, 178.1225 222.1152, 481.2542 | 1.28 | X | X | |||||
| Other | N-acetylornithine | C7H14N2O3 | 175.1187 | 152.9637 | 0.40 | X | X |
| Strain (Abbreviation) | Origin | Max Growth Rates (d−1) | MC Producer |
|---|---|---|---|
| CPCC300 | Deposited by E. Prepas/A. Lam. Isolated by A. Lam from Pretzlaff Pond, Alberta, Canada, Aug. 1990. | 0.600 | Yes |
| CPCC464 | Deposited by D. Parker. Isolated by E.A.D. Allen/P. Gorham from Trampling Lake, SK, Canada, Aug. 1980. | 0.048 | Yes |
| CPCC299 | Deposited by E. Prepas/A.Lam. Isolated by A. Lam from Pretzlaff Pond, Alberta, Canada, Aug. 1990 | 0.024 | Yes |
| PCC7806 (WT) | Deposited by F.I. Kappers. Isolated from Braakman Reservoir, Netherlands, 1984. | 0.024 | Yes |
| PCC7806 mcyB- (mcyB-) | Deposited by E. Dittman, 1997. Genetically modified from PCC7806. | 0.072 | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racine, M.; Saleem, A.; Pick, F.R. Metabolome Variation between Strains of Microcystis aeruginosa by Untargeted Mass Spectrometry. Toxins 2019, 11, 723. https://doi.org/10.3390/toxins11120723
Racine M, Saleem A, Pick FR. Metabolome Variation between Strains of Microcystis aeruginosa by Untargeted Mass Spectrometry. Toxins. 2019; 11(12):723. https://doi.org/10.3390/toxins11120723
Chicago/Turabian StyleRacine, Marianne, Ammar Saleem, and Frances R. Pick. 2019. "Metabolome Variation between Strains of Microcystis aeruginosa by Untargeted Mass Spectrometry" Toxins 11, no. 12: 723. https://doi.org/10.3390/toxins11120723
APA StyleRacine, M., Saleem, A., & Pick, F. R. (2019). Metabolome Variation between Strains of Microcystis aeruginosa by Untargeted Mass Spectrometry. Toxins, 11(12), 723. https://doi.org/10.3390/toxins11120723