Review of DSP Toxicity in Ireland: Long-Term Trend Impacts, Biodiversity and Toxin Profiles from a Monitoring Perspective


Abstract
:1. Introduction
2. Results
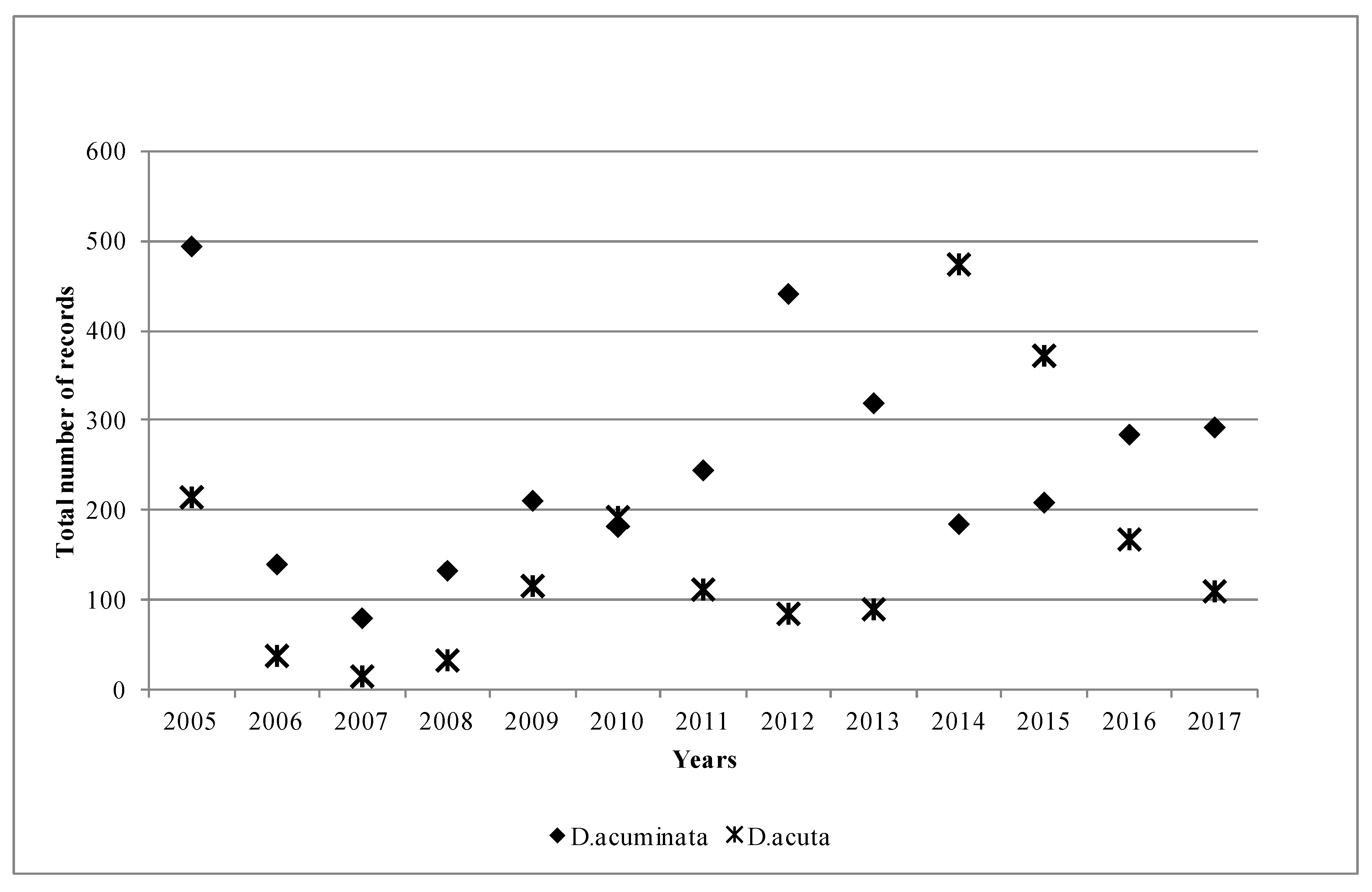
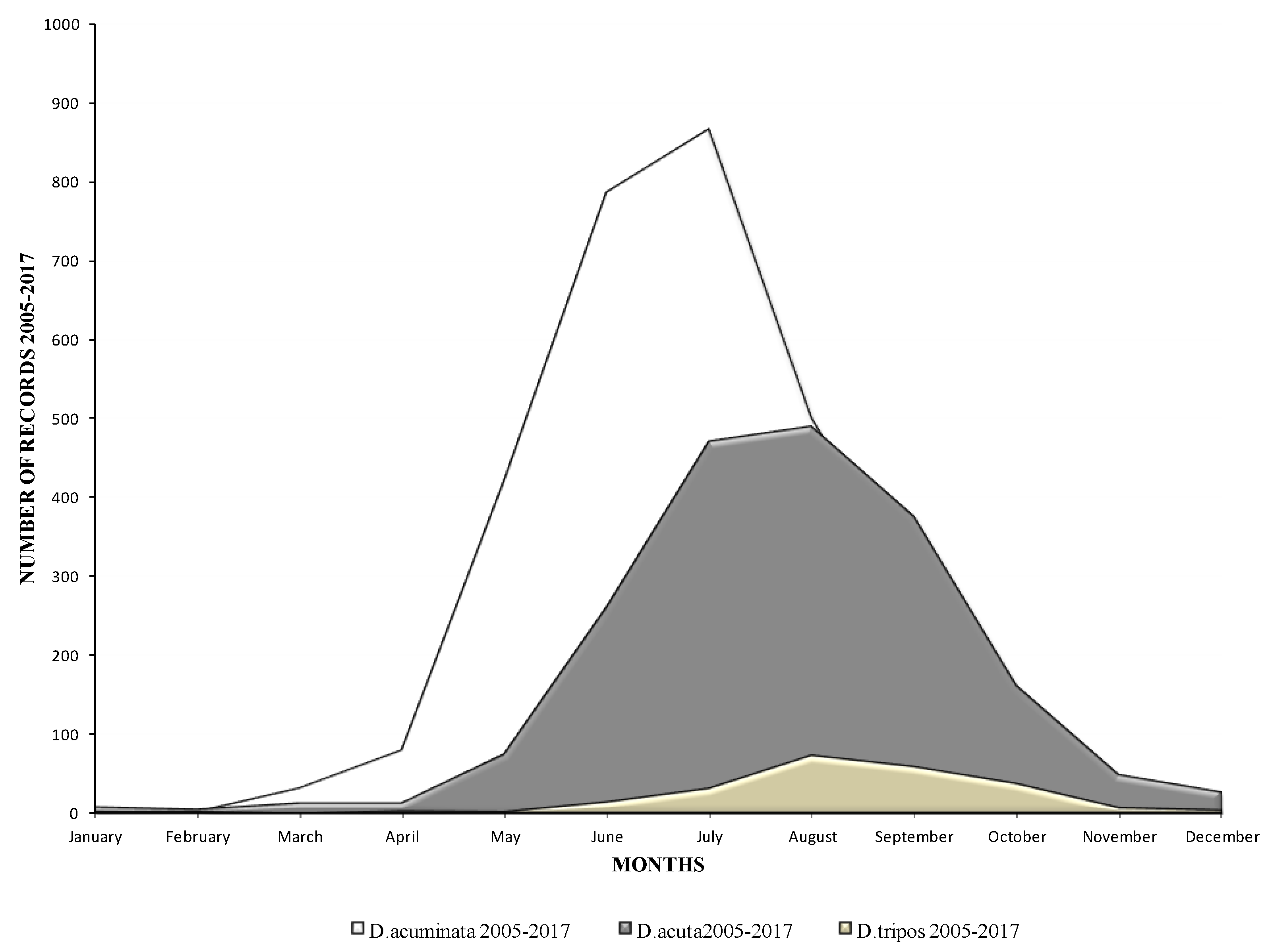
2.1. Phytoplankton Records from the HABs Database
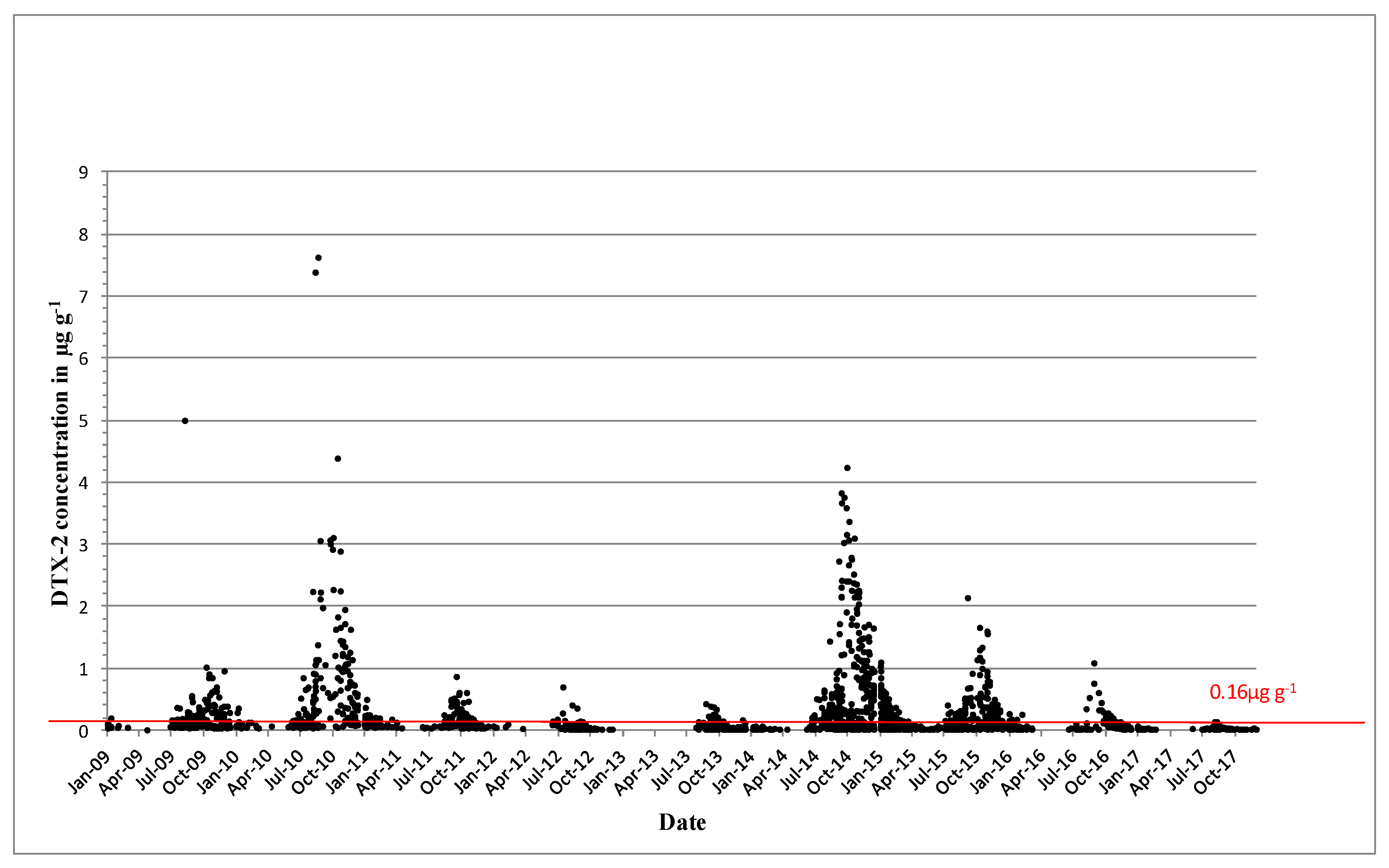
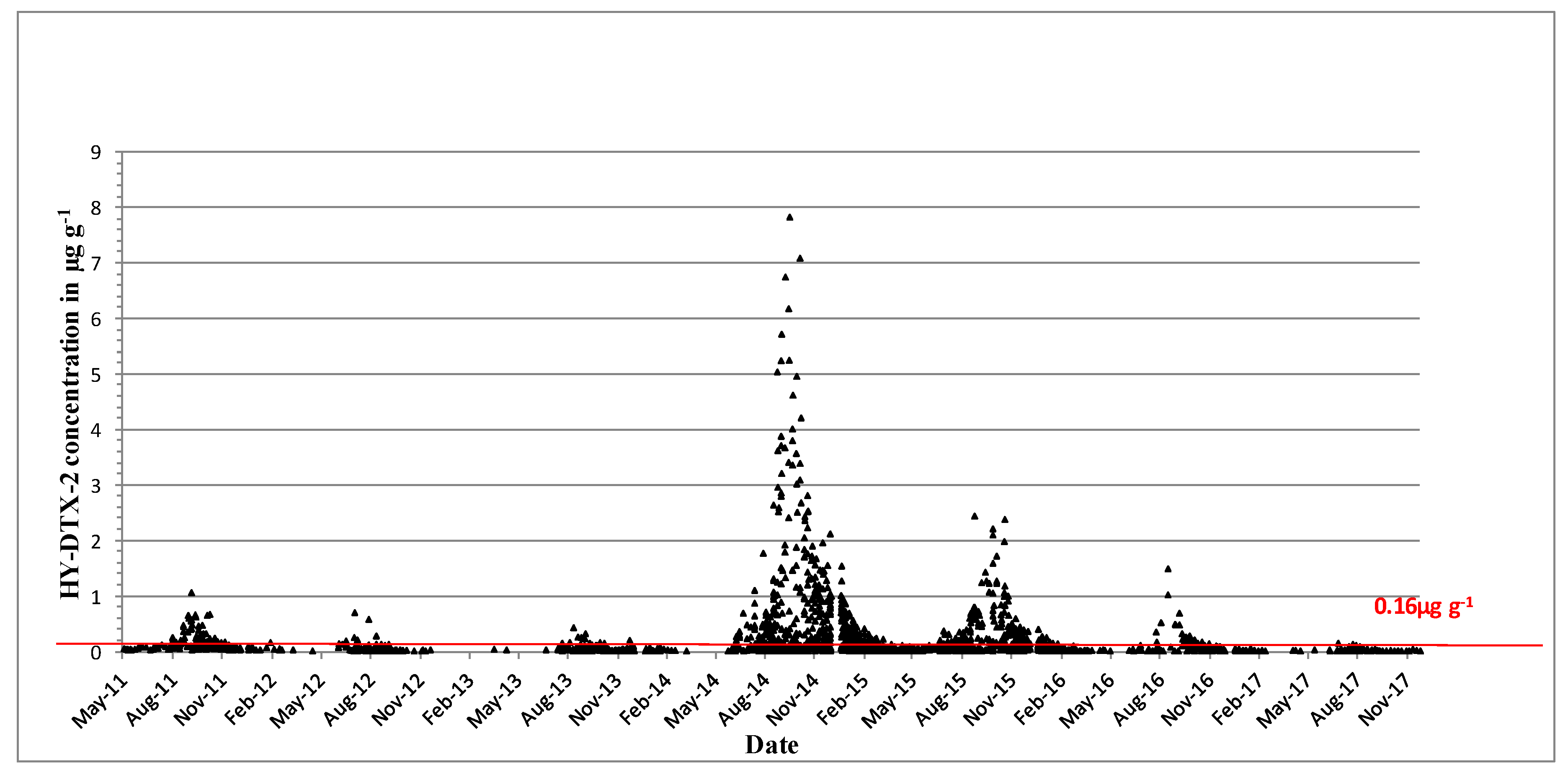
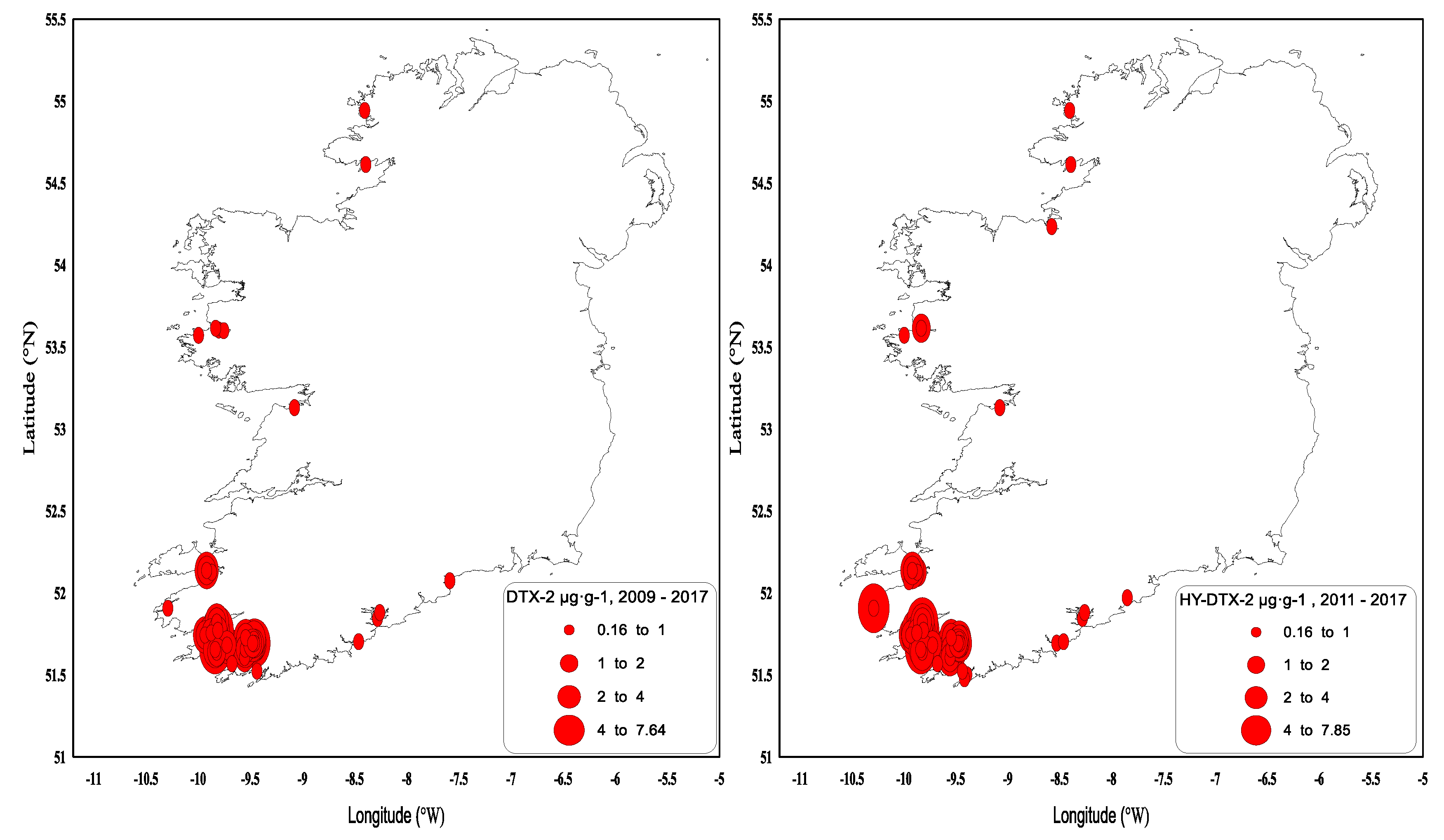
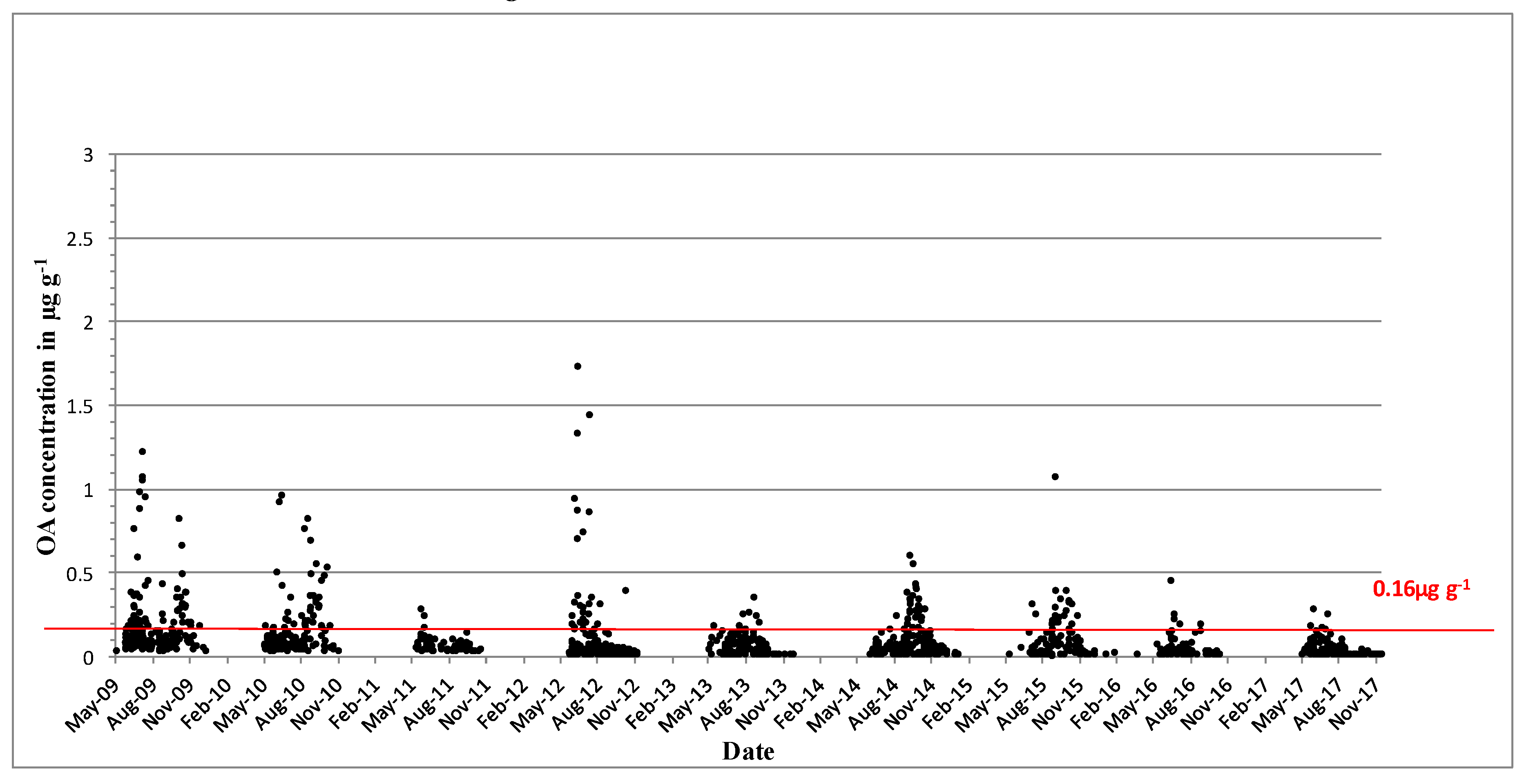
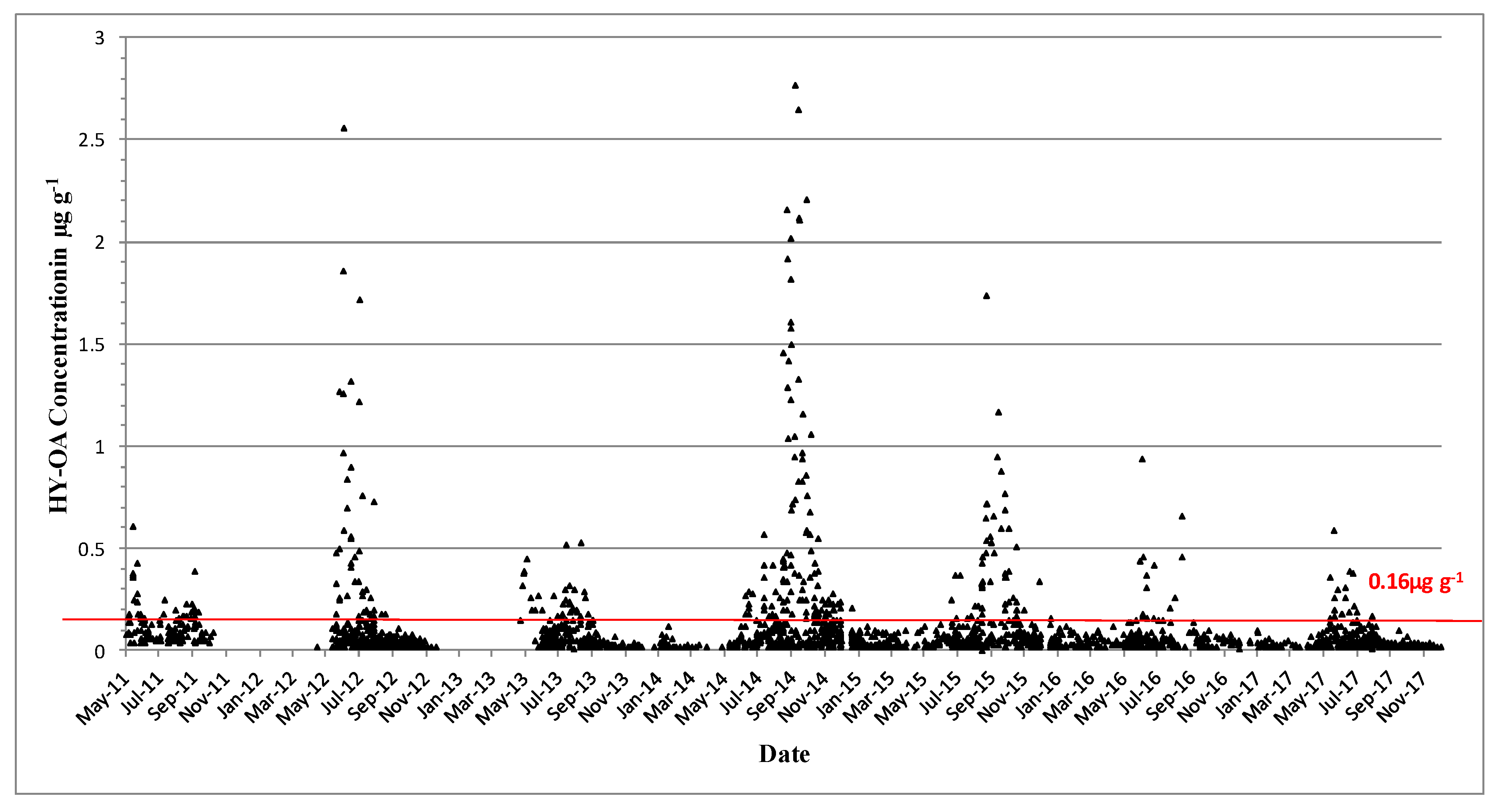
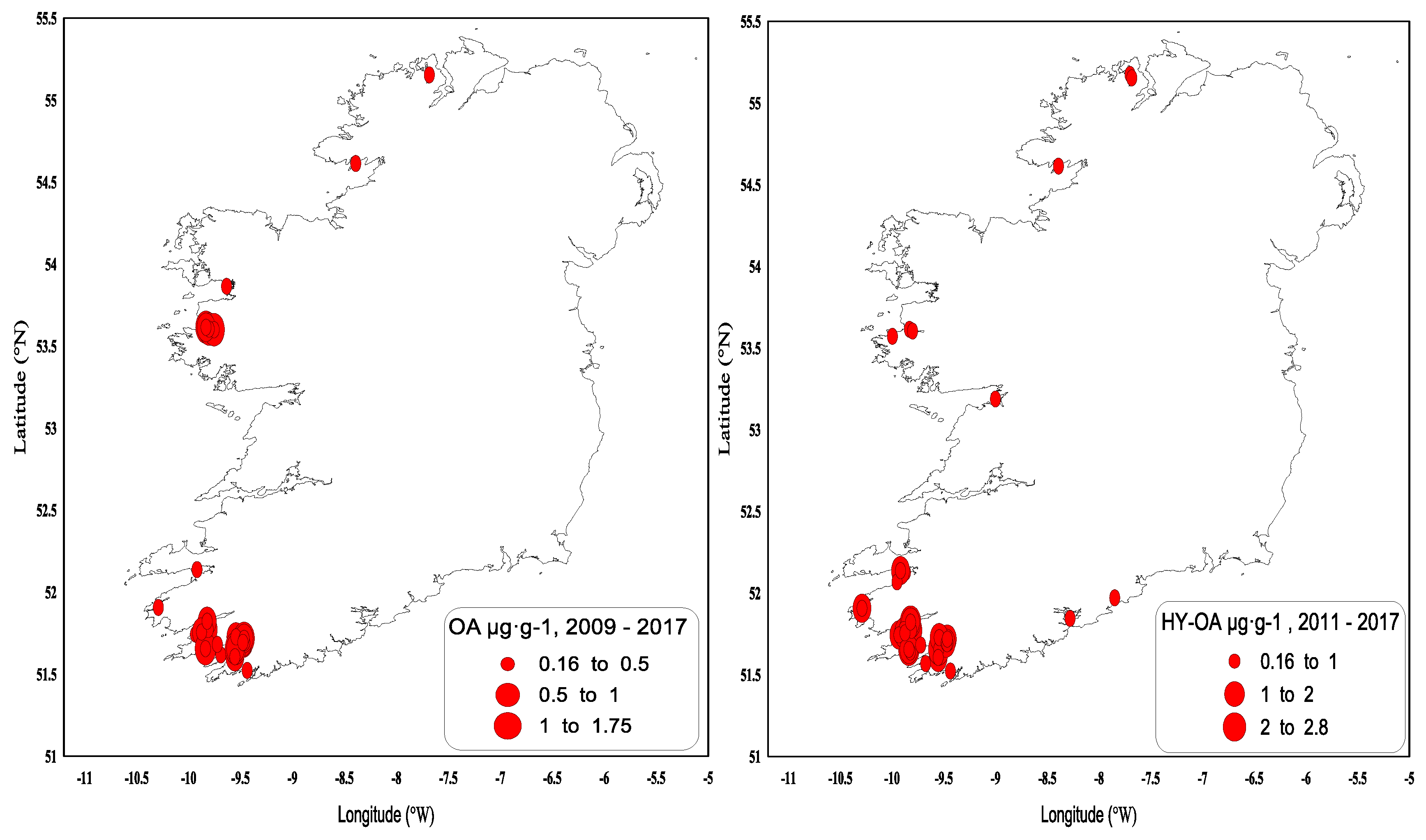
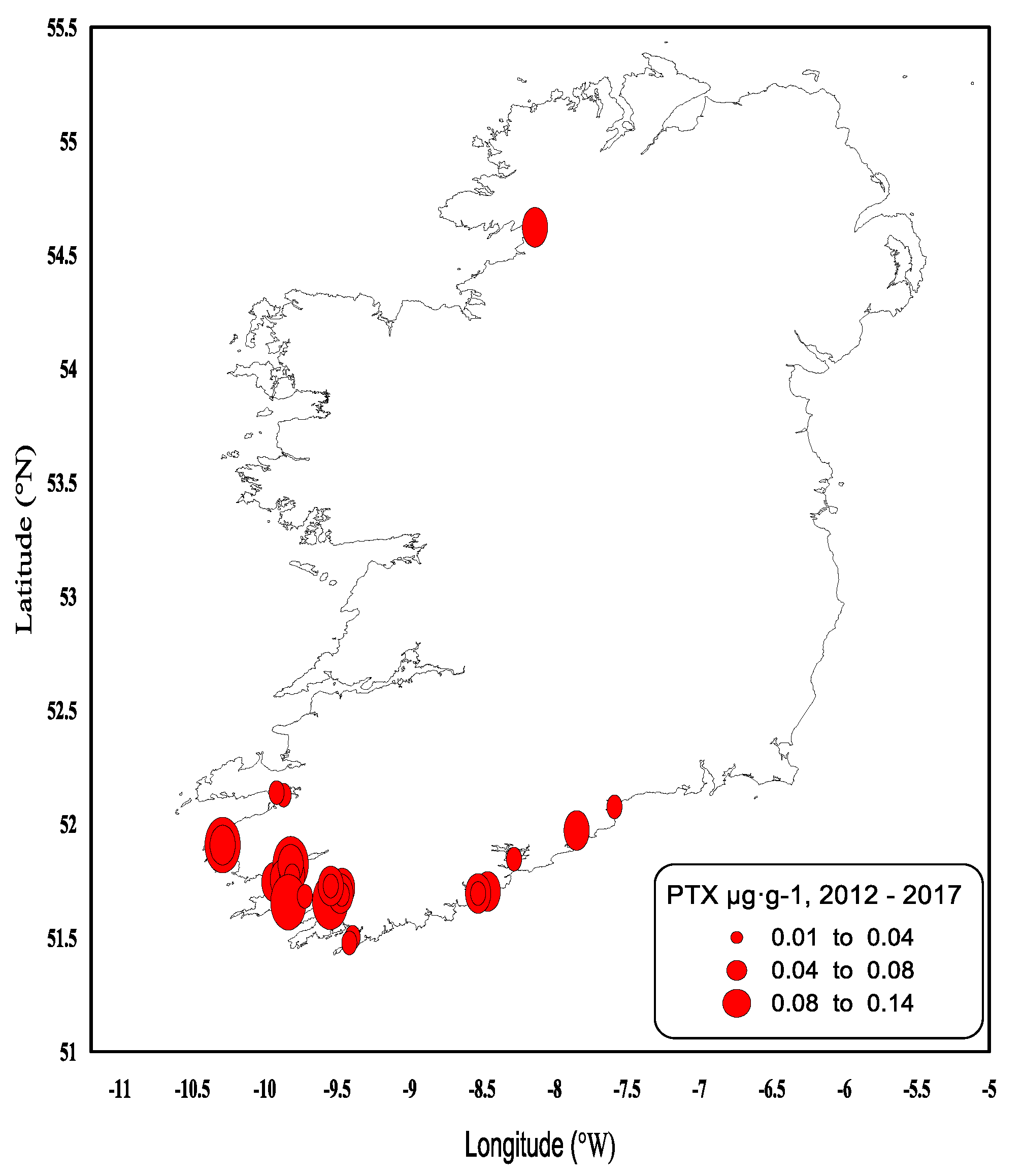
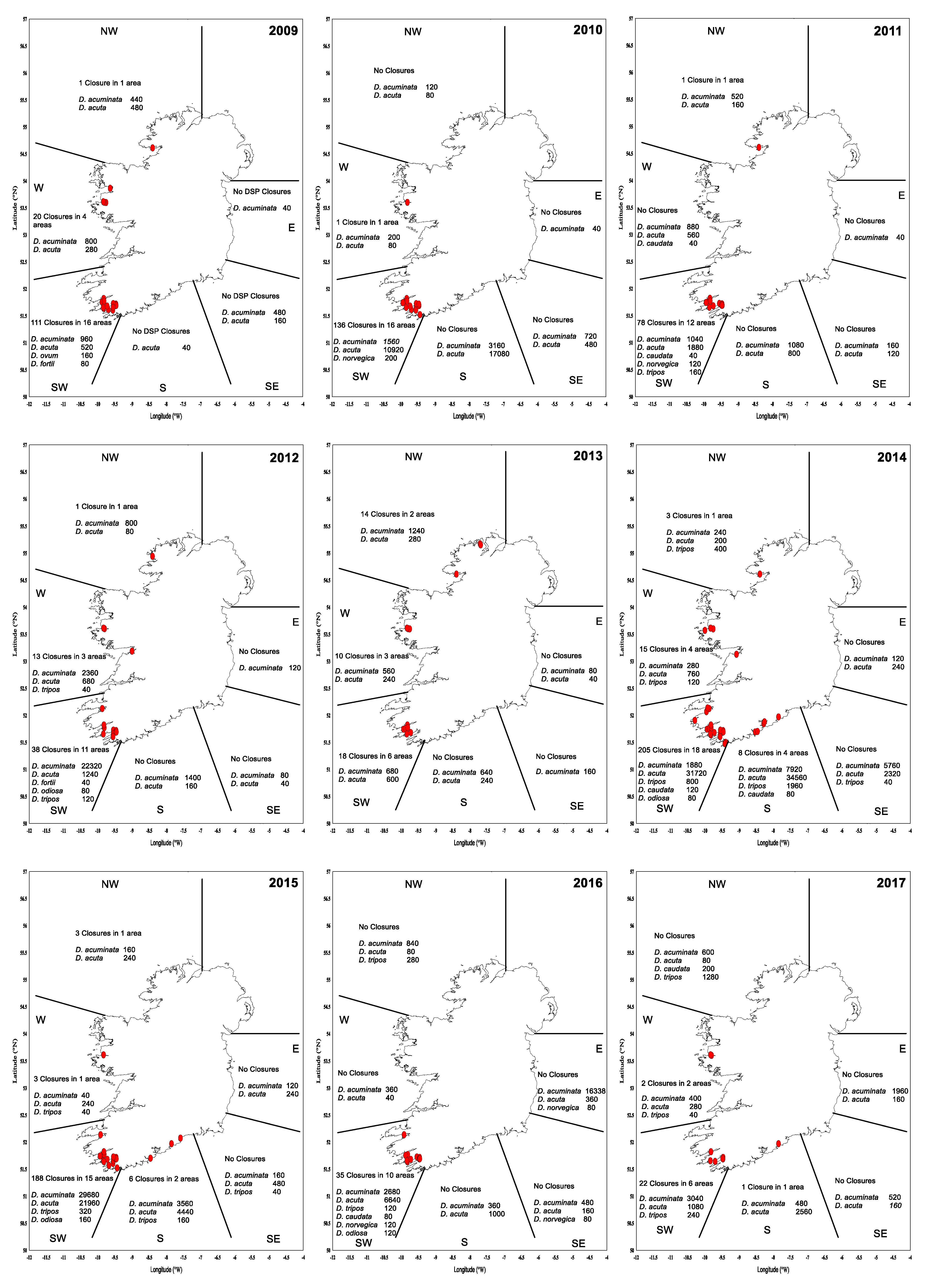
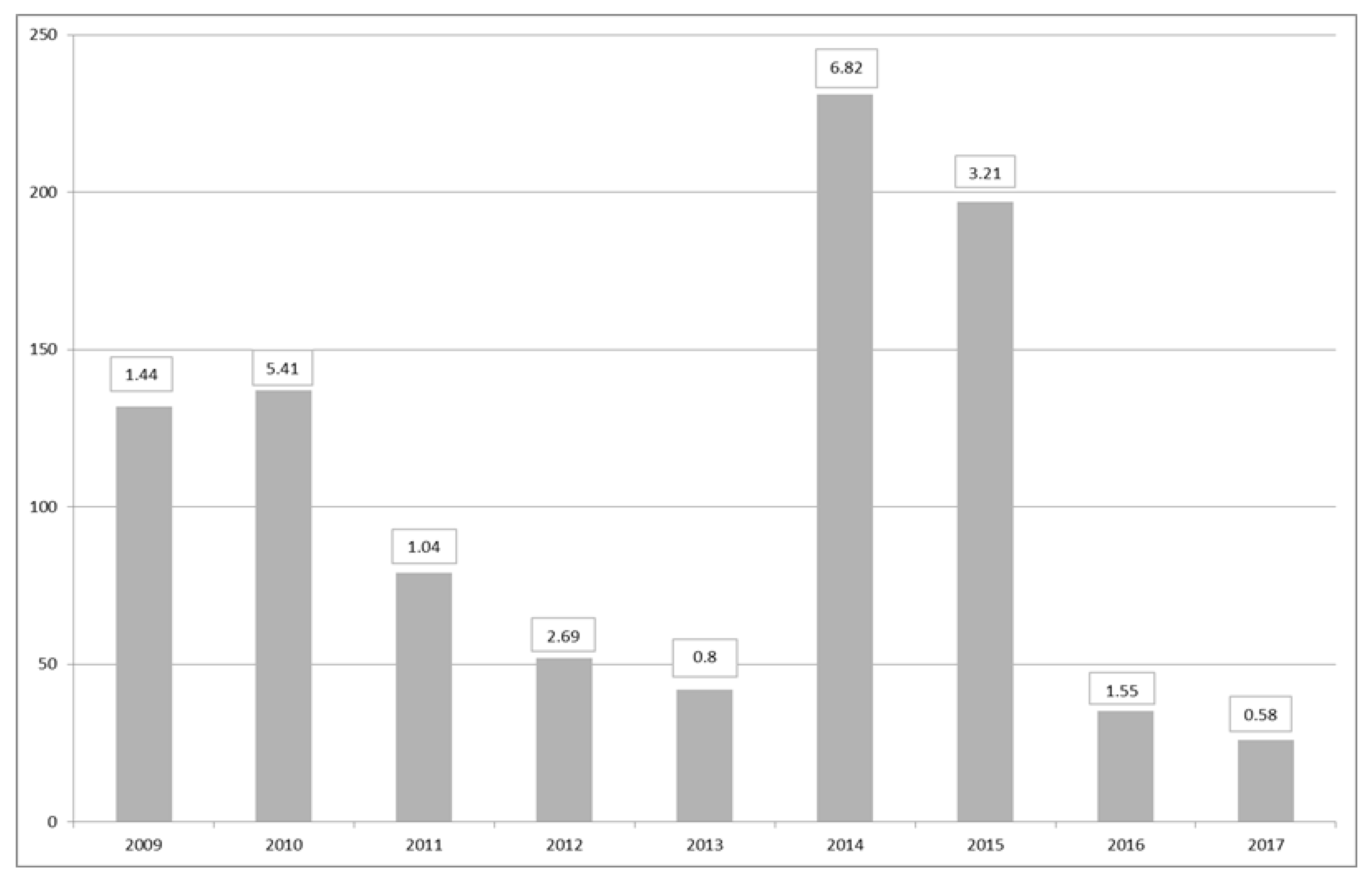
2.2. Shellfish Toxicity Data
3. Discussion
4. Materials and Methods
4.1. Biotoxin National Monitoring Programme
4.1.1. Sample Collection, Delivery and Lab Receipt
4.1.2. LC-MS/MS Methodology
4.1.3. Sample Results and Reporting
4.1.4. Chemistry Data Review and Result Interpretation
4.2. Phytoplankton National Monitoring Programme in Ireland
4.2.1. Sample Collection, Transport and Delivery
4.2.2. Sample Analysis
4.2.3. Sample Results and Reporting
4.2.4. Harmful Algal Bloom (HAB) Database Phytoplankton Data Extraction
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kat, M. The occurrence of Prorocentrum species and coincidental gastointestinal illness of mussel consumers. In Toxic Dinoflagellate Blooms, Developments in Marine Biology; Taylor, D.L., Seliger, H.H., Eds.; Elsevier/North-Holland: New York, NY, USA, 1979; Volume 1, pp. 215–220. [Google Scholar]
- Kat, M. Dinophysis acuminata blooms in the Dutch coastal area related to diarrhetic mussel poisoning in the Dutch Waddensea. Sarsia 1983, 68, 81–84. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku District. Bull. Jpn. Soc. Sci. Fish 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish 1980, 46, 1045–1411. [Google Scholar] [CrossRef]
- McMahon, T.; Silke, J.; Nixon, E.; Taffe, B.; Nolan, A.; McGovern, E.; Doyle, J. Seasonal variation in diarrhetic shellfish toxins in mussels from the southwest coast of Ireland in 1994. In Irish Marine Science 1995; Keegan, B.F., O’Connor, R., Eds.; Galway University Press: Galway, Ireland, 1995; pp. 417–432. [Google Scholar]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Hess, P.; Dell’ Aversano, C. Recent developments in the analysis of phycotoxins by liquid chromatography–mass spectrometry. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Millennium; De Koe, W.J., Samson, R.A., van Egmond, H.P., Gilbert, J., Sabino, M., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2002; pp. 383–392. [Google Scholar]
- Gerssen, P.P.J.; Mulder, M.A.; McElhinney, J.B. Liquid chromatography–tandem mass spectrometry method for the detection of marine lipophilic toxins under alkaline conditions. J. Chromatogr. A 2009, 9, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Shimatani, M.; Sugitani, H.; Oshima, Y.; Yasumoto, T. Isolation and structural elucidation of the causative toxin of the diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish 1982, 48, 549–552. [Google Scholar] [CrossRef]
- Hu, T.; Marr, J.; de Freitas, A.S.W.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C.; Pleasance, S. New diol esters isolated from cultures of the dinoflagellate Prorocentrum lima and Prorocentrum concavum. J. Nat. Prod. 1992, 55, 1631–1637. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Matsumoto, K.; Clardy, J. Diarrhetic shellfish poisoning. In Seafood Toxins, ACS Symposium Series; Ragelis, P., Ed.; American Chemical Society: Washington, DC, USA, 1984; pp. 207–214. [Google Scholar]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Draisci, R.; Lucentini, L.; Mascioni, A. Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 2000; pp. 289–324. [Google Scholar]
- Utermöhl, H. Zur Vervollkomnung der quantitativen phytoplankton-Methodik. Mitt. Int. Ver. Limnol. 1958, 9, 1–38. [Google Scholar]
- Raine, R.; O’Mahoney, J.; McMahon, T.; Roden, C. Hydrography and phytoplankton of waters off South-west Ireland. Estuar. Coast. Shelf Sci. 1990, 30, 579–592. [Google Scholar] [CrossRef]
- Edwards, A.; Jones, K.; Graham, J.M.; Griffiths, C.R.; MacDougall, N.; Patching, J.P.; Richard, J.M.; Raine, R. Transient Coastal Upwelling and Water Circulation in Bantry. Estuar. Coast. Shelf Sci. 1996, 42, 213–230. [Google Scholar] [CrossRef]
- Elliott, A.J.; Clarke, T. Seasonal stratification in the northwest European shelf seas. Cont. Shelf Res. 1991, 11, 467–492. [Google Scholar] [CrossRef]
- Raine, R.; McMahon, T. Physical dynamics on the continental shelf off southwestern Ireland and their influence on coastal phytoplankton blooms. Cont. Shelf Res. 1998, 18, 883–914. [Google Scholar] [CrossRef]
- Fernand, L.; Nolan, G.D.; Raine, R.; Chambers, C.E.; Dye, S.R.; White, M.; Brown, J. The Irish coastal current: A seasonal jet-like circulation. Cont. Shelf Res. 2006, 26, 1775–1793. [Google Scholar] [CrossRef] [Green Version]
- Raine, R.; McDermott, G.; Silke, J.; Lyons, K.; Nolan, G.; Cusack, C. A simple short range model for the prediction of harmful algal events in the bays of southwestern Ireland. J. Mar. Syst. 2010, 83, 150–157. [Google Scholar] [CrossRef]
- Rodriguez, F.; Escalera, L.; Reguera, B.; Rial, P.; Riobó, P.; de Jesús da Silva, T. Morphological variability, toxinology and genetics of the dinoflagellate Dinophysis tripos (Dinophysiaceae, Dinophysiales). Harmful Algae 2012, 13, 26–33. [Google Scholar] [CrossRef]
- Fabro, E.; Almandoz, G.O.; Ferrario, M.; Tillmann, U.; Cembella, A.; Krock, B. Distribution of Dinophysis species and their association with lipophilic phycotoxins in plankton from the Argentine Sea. Harmful Algae 2016, 59, 31–41. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Areas | |||||||
|---|---|---|---|---|---|---|---|
| Species Name | East | South East | South | South West | West | North West | Total Number of Observations |
| Dinophysis acuminata | 160 | 103 | 126 | 1417 | 737 | 555 | 3098 |
| Dinophysis acuta | 109 | 51 | 141 | 1241 | 277 | 119 | 1938 |
| Dinophysis tripos | 0 | 2 | 9 | 151 | 13 | 46 | 221 |
| Dinophysis caudata | 0 | 0 | 1 | 12 | 1 | 3 | 17 |
| Dinophysis norvegica | 3 | 3 | 0 | 4 | 0 | 0 | 10 |
| Dinophysis odiosa | 0 | 0 | 0 | 10 | 0 | 0 | 10 |
| Dinophysis fortii | 0 | 0 | 1 | 7 | 1 | 0 | 9 |
| Dinophysis hastata | 0 | 0 | 0 | 0 | 6 | 1 | 7 |
| Dinophysis ovum | 0 | 0 | 0 | 3 | 1 | 0 | 4 |
| Dinophysis nasuta | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| Normalised, Estimated Number of Observations by Geographical Area | |||
|---|---|---|---|
| Regions | D. acuminata | D. acuta | Ratio D. acuminata/D. acuta |
| East | 134 | 91 | 0.68 |
| Southeast | 73 | 36 | 0.49 |
| South | 158 | 190 | 1.20 |
| Southwest | 345 | 288 | 0.83 |
| West | 196 | 73 | 0.37 |
| Northwest | 190 | 44 | 0.23 |
| Species Name | Common Name | DTX-2 µg∙g−1 | HY-DTX-2 µg∙g−1 | OA µg∙g−1 | HY-OA µg∙g−1 |
|---|---|---|---|---|---|
| Mytilus edulis | Blue mussel | 7.63 | 7.84 | 1.74 | 2.77 |
| Pecten maximus | King scallop | 0.27 | 7.1 | 0.28 | 1.92 |
| Crassostrea gigas | Pacific oyster | 0.36 | 0.29 | <RL | <RL |
| Cerastoderma edule | Common cockle | <RL | 0.75 | <RL | 0.34 |
| Spisula solida | Surf clam | <RL | 0.16 | <RL | 0.22 |
| Years | Southwest | |||||||||||
| Jan. | Feb. | Mar. | Apr. | May | Jun. | Jul. | Aug. | Sep. | Oct. | Nov. | Dec. | |
| 2009 | - | - | - | - | - | 14 | 15 | 16 | 25 | 22 | 16 | 3 |
| 2010 | 2 | - | - | - | 4 | 13 | 16 | 28 | 15 | 20 | 28 | 10 |
| 2011 | - | - | - | - | 7 | 12 | 2 | 13 | 21 | 19 | 2 | - |
| 2012 | - | - | - | - | - | 12 | 15 | 9 | 2 | - | - | - |
| 2013 | - | - | - | - | - | - | 4 | 8 | 6 | - | - | - |
| 2104 | - | - | - | - | - | 10 | 12 | 23 | 49 | 34 | 41 | 36 |
| 2015 | 56 | 16 | 3 | - | - | 1 | 8 | 16 | 20 | 22 | 30 | 16 |
| 2016 | 4 | - | - | - | - | 8 | 4 | 5 | 6 | 8 | - | - |
| 2017 | - | - | - | - | 3 | 10 | 5 | 3 | - | - | - | - |
| Years | West | |||||||||||
| Jan. | Feb. | Mar. | Apr. | May | Jun. | Jul. | Aug. | Sep. | Oct. | Nov. | Dec. | |
| 2009 | - | - | - | - | - | 5 | 13 | 2 | - | - | - | - |
| 2010 | - | - | - | - | - | 1 | - | - | - | - | - | - |
| 2011 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2012 | - | - | - | - | - | 3 | 8 | 1 | 1 | - | - | - |
| 2013 | - | - | - | - | - | - | 1 | 6 | 3 | - | - | - |
| 2104 | - | - | - | - | - | - | - | 2 | 6 | 5 | 2 | - |
| 2015 | - | - | - | - | - | - | - | 3 | - | - | - | - |
| 2016 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2017 | - | - | - | - | - | - | - | 2 | - | - | - | - |
| Years | Northwest | |||||||||||
| Jan. | Feb. | Mar. | Apr. | May | Jun. | Jul. | Aug. | Sep. | Oct. | Nov. | Dec. | |
| 2009 | - | - | - | - | - | - | - | - | 1 | - | - | - |
| 2010 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2011 | - | - | - | - | - | - | 1 | - | - | - | - | - |
| 2012 | - | - | - | - | - | - | - | 1 | - | - | - | - |
| 2013 | - | - | - | - | - | 7 | 3 | 4 | - | - | - | - |
| 2104 | - | - | - | - | - | - | - | 1 | 2 | - | - | - |
| 2015 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2016 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2017 | - | - | - | - | - | - | - | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salas, R.; Clarke, D. Review of DSP Toxicity in Ireland: Long-Term Trend Impacts, Biodiversity and Toxin Profiles from a Monitoring Perspective. Toxins 2019, 11, 61. https://doi.org/10.3390/toxins11020061
Salas R, Clarke D. Review of DSP Toxicity in Ireland: Long-Term Trend Impacts, Biodiversity and Toxin Profiles from a Monitoring Perspective. Toxins. 2019; 11(2):61. https://doi.org/10.3390/toxins11020061
Chicago/Turabian StyleSalas, Rafael, and Dave Clarke. 2019. "Review of DSP Toxicity in Ireland: Long-Term Trend Impacts, Biodiversity and Toxin Profiles from a Monitoring Perspective" Toxins 11, no. 2: 61. https://doi.org/10.3390/toxins11020061
APA StyleSalas, R., & Clarke, D. (2019). Review of DSP Toxicity in Ireland: Long-Term Trend Impacts, Biodiversity and Toxin Profiles from a Monitoring Perspective. Toxins, 11(2), 61. https://doi.org/10.3390/toxins11020061