Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus



Abstract
:1. Introduction
2. Results
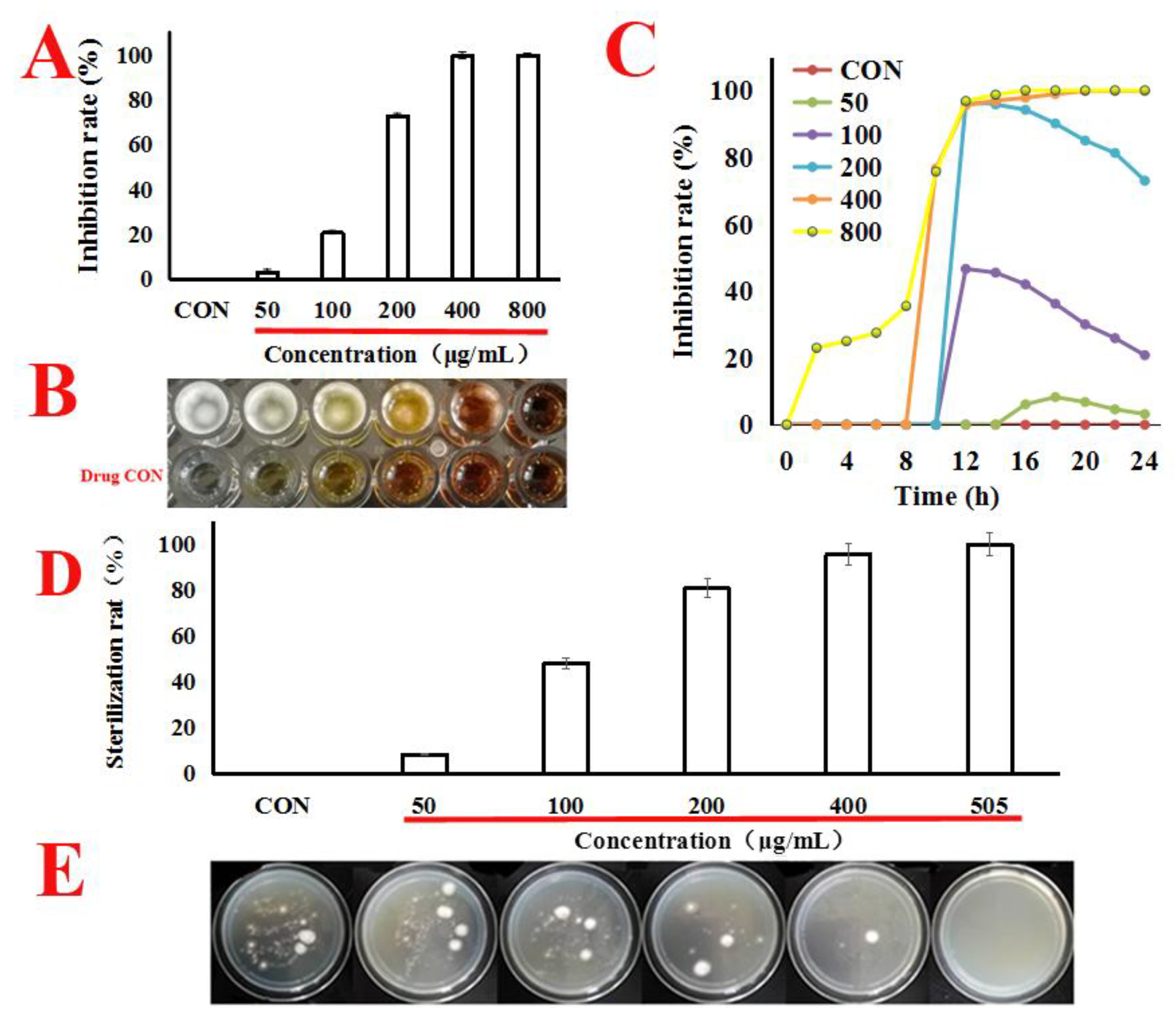
2.1. Quercetin Inhibited the Proliferation of A. flavus
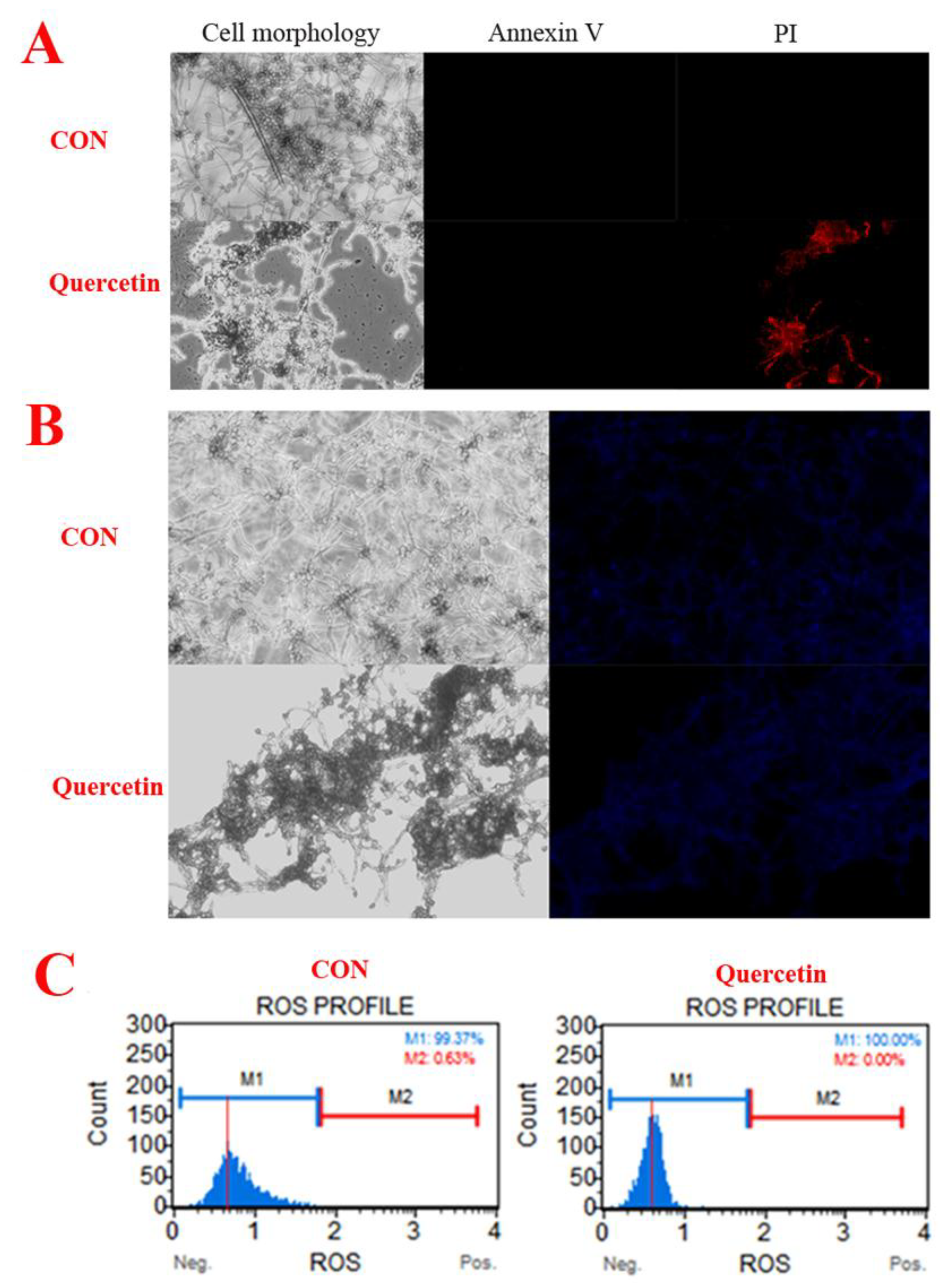
2.2. Morphological Changes of A. flavus
2.3. Cell Apoptosis
2.4. RNA-Seq Data
2.5. Identification and Functional Annotation
2.6. Expression Analysis of Conidial Development- and A. flavus Growth-Related Genes in Response to Quercetin
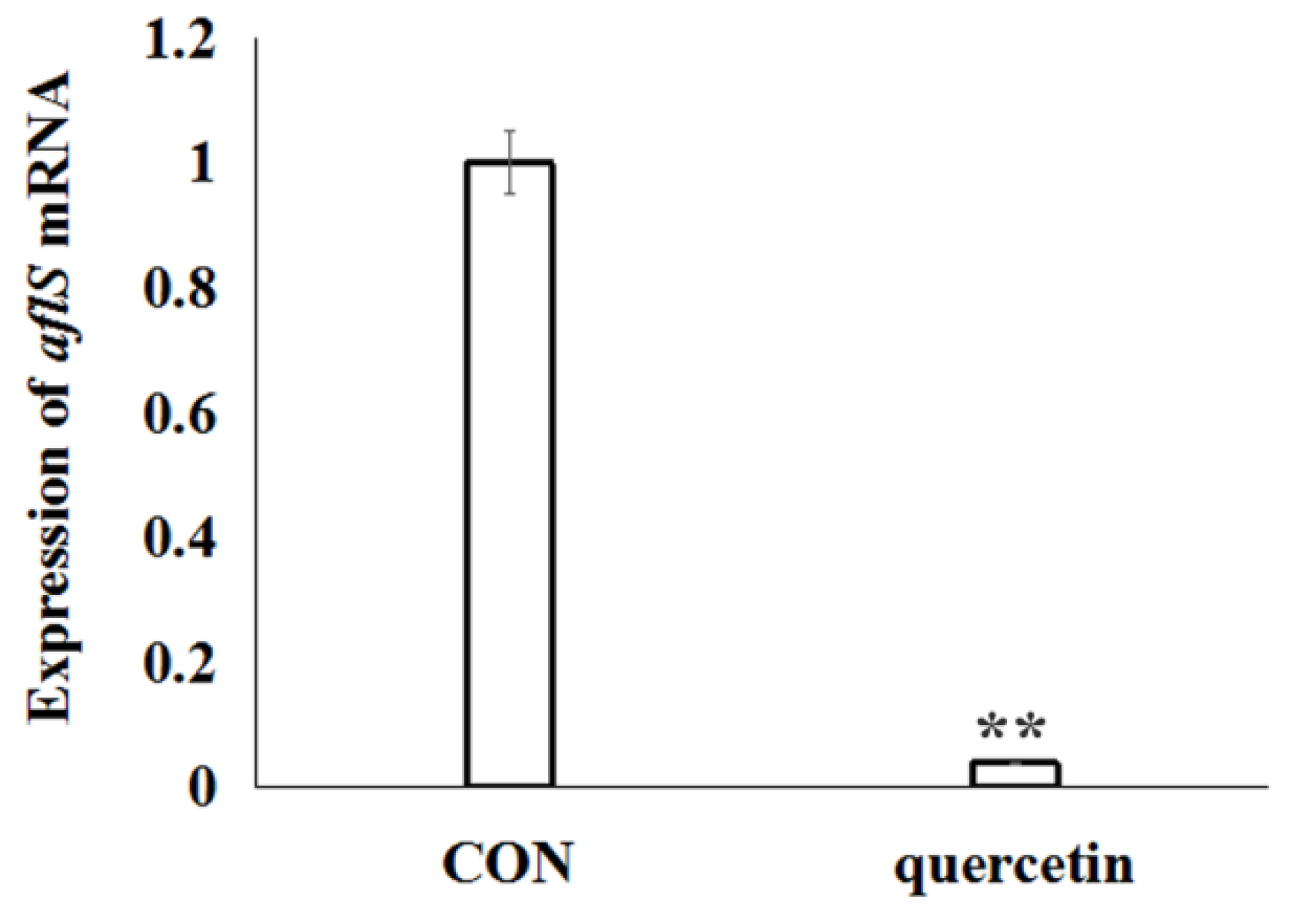
2.7. Expression Analysis of A. flavus AF Biosynthesis-Related Genes in Response to Quercetin
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Fungus Strain and Cultivation
4.3. Anti-Proliferative Activity
4.4. Morphological Changes of A. flavus
4.5. Phosphatidylserine (PS) Externalization
4.6. Hochest 33342
4.7. Measurement of Reactive Oxygen Species
4.8. cDNA Preparation and Illumina Sequencing
4.9. RNA-Seq and Enrichment Analysis of Differentially Expressed Genes
4.10. Validation of RNA-Seq Analysis by Quantitative Real-Time PCR (qRT-PCR)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cary, J.W.; Han, Z.; Yin, Y.; Lohmarc, J.M.; Shantappa, S.; YHarris-Coward, P. Transcriptome analysis of Aspergillus flavus reveals veA-dependent regulation of secondary metabolite gene clusters, including the novel aflavarin cluster. Eukaryot. Cell. 2015, 14, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Kelle, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.F.; Gibbons, J.G.; Lee, M.K.; Han, K.H.; Hong, S.B.; Yu, J.H. Controlling aflatoxin contamination and propagation of Aspergillus flavus by a soy-fermenting Aspergillus oryzae strain. Sci. Rep. 2018, 8, 16871. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; Snini, S.P.; Puel, O.; Mathieu, F. Streptomyces roseolus, A promising biocontrol agent against Aspergillus flavus, the main Aflatoxin B₁producer. Toxins 2018, 30, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Liu, J.; Pan, F.F.; Shi, D.D.; Wen, Z.G.; Yang, P.L. Quercetin and aconitine synergistically induces the human cervical carcinoma HeLa cell apoptosis via endoplasmic reticulum (ER) stress pathway. PLoS ONE 2018, 13, e0191062. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Zhang, H.; Yang, J.; Hu, L.B.; MO, H.Z. Inhibition Effect of Quercetin on Aflatoxin B1 Production. Mod. Food Sci. Technol. 2014, 30, 56–62. [Google Scholar]
- Wang, X.Y.; Wang, Y.Z.; Zhou, Y.G.; Wei, X.L. Farnesol induces apoptosis-like cell death in the pathogenic fungus Aspergillus flavus. Mycologia 2014, 106, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Lei, Y.; Yan, L.Y.; Cheng, K.; Dai, X.F.; Wan, L.Y. Deep sequencing analysis of transcriptomes in Aspergillus flavus in response to resveratrol. BMC Microbiol. 2015, 15, 182. [Google Scholar] [CrossRef]
- Lv, C.; Wang, P.; Ma, L.X.; Zheng, M.M.; Liu, Y.; Xing, F.G. Large-scale comparative analysis of Eugenol-Induced/Repressed genes expression in Aspergillus flavus using RNA-seq. Front. Microbiol. 2018, 9, 1116. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Q.; Zhao, X.X.; Zhi, Q.Q.; Zhao, M.; He, Z.M. Transcriptomic profiling of Aspergillus flavus in response to 5-azacytidine. Fungal Genet. Biol. 2013, 56, 78–86. [Google Scholar] [CrossRef]
- Li, X.M.; Luo, X.G.; Si, C.L.; Wang, N.; Zhou, H.; He, J.F. Antibacterial active compounds from Hypericum ascyron L. induce bacterial cell death through apoptosis pathway. Eur. J. Med. Chem. 2015, 96, 436–444. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.F.; de Giovani, W.F. Antioxidant properties of complexes of flavonoids with metal ions. Redox Rep. 2004, 9, 97–104. [Google Scholar] [CrossRef]
- Yoon, J.S.; Chae, M.K.; Lee, S.Y.; Lee, E.J. Anti-inflammatory effect of quercetin in a whole orbital tissue culture of Graves’ orbitopathy. Br. J. Ophthalmol. 2012, 96, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. J. Vis. Exp. 2011, 50, 2597. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.O.; Tiffany, M.F.; Qualls, C.R.; Key, C.R. The rising incidence of adenocarcinoma relative to squamous cell carcinoma of the uterine cervix in the United States-a 24-year population-based study. Gynecol. Oncol. 2000, 78, 97–105. [Google Scholar] [CrossRef]
- Chang, T.W.; Chen, C.C.; Chen, K.Y.; Su, J.H.; Chang, J.H.; Chang, M.C. Ribosomal phosphoprotein P0 interacts with GCIP and overexpression of P0 is associated with cellular proliferation in breast and liver carcinoma cells. Oncogene 2008, 27, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Akanuma, G.; Nanamiya, H.; Natori, Y.; Yano, K.; Suzuki, S.; Omata, S. Inactivation of ribosomal protein genes in Bacillus subtilis reveals importance of each ribosomal protein for cell proliferation and cell differentiation. J. Bacteriol. 2012, 194, 6282–6291. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Ito, S.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H. A-tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS. Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef]
- Wang, H.M.; Lei, Y.; Yan, L.Y.; Wan, L.Y.; Ren, X.P.; Chen, S.L. Functional genomic analysis of Aspergillus flavus interacting with resistant and susceptible peanut. Toxins 2016, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar]
- Calvo, A.M.; Cary, J.W. Association of fungal secondary metabolism and sclerotial biology. Front. Microbiol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.S.; Yue, Y.W.; Fu, Y.S.; Fang, Z.; Xu, Z.L.; Ma, G.L. Exploration of the regulatory mechanism of secondary metabolism by comparative transcriptomics in Aspergillus flavus. Front. Microbiol. 2018, 9, 1568–1583. [Google Scholar]
- Benvenuto, M.; Sileri, P.; Rossi, P.; Masuelli, L.; Fantini, M.; Nanni, M. Natural humoral immune response to ribosomal P0 protein in colorectal cancer patients. J. Trans. Med. 2015, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Remacha, M.; Jimenez-Diaz, A.; Santos, C.; Briones, E.; Zambrano, R.; Rodriguez-Gabriel, M.A. Proteins P1, P2, and P0, components of the eukaryotic ribosome stalk. New structural and functional aspects. Biochem. Cell Biol. 1995, 73, 959–968. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Zhang, B.C.; Zhang, A.R.; Wu, T.T.; Liu, J.; Yu, L.F.; Wang, W.-X.; Gao, J.-F.; Fang, D.-C.; Rao, Z.-G. Co-transduction of ribosomal protein L23 enhances the therapeutic efficacy of adenoviral-mediated p53 gene transfer in human gastric cancer. Oncol. Rep. 2013, 30, 1989–1995. [Google Scholar] [CrossRef]
- Qi, Y.K.; Li, X.; Chang, C.K.; Xu, F.; He, Q.; Zhao, Y.S. Ribosomal protein L23 negatively regulates cellular apoptosis via the RPL23/Miz-1/c-Myc circuit in higher-risk myelodysplastic syndrome. Sci. Rep. 2016, 7, 2323. [Google Scholar] [CrossRef]
- Pidutti, P.; Federici, F.; Brandi, J.; Manna, L.; Rizzi, E.; Marini, U. Purification and characterization of ribosomal proteins L27 and L30 having antimicrobial activity produced by the Lactobacillus salivarius SGL03. J. Appl. Microbiol. 2017, 124, 398–407. [Google Scholar] [CrossRef]
- Smolock, E.M.; Korshunov, V.A.; Glazko, G.; Qiu, X.; Gerloff, J.; Berk, B.C. Ribosomal Protein L17, RpL17, is an Inhibitor of Vascular Smooth Muscle Growth and Carotid Intima Formation. Circulation 2012, 126, 2418–2427. [Google Scholar] [CrossRef] [Green Version]
- Mina, H.; Kim, H.R.; Kim, I. Ribosomal protein L19 overexpression activates the unfolded protein response and sensitizes MCF7 breast cancer cells to endoplasmic reticulum stress-induced cell death. Biochem. Biophs. Res. Commun. 2014, 450, 673–678. [Google Scholar]
- Bee, A.; Brewer, D.; Beesley, C.; Dodson, A.; Forootan, S.; Dickinson, T. SiRNA knockdown of ribosomal protein gene RPL19 abrogates the aggressive phenotype of human prostate cancer. PLoS ONE 2011, 6, e22672. [Google Scholar] [CrossRef]
- Cao, B.; Fang, Z.L.; Liao, P.; Zhou, X.; Xiong, J.P.; Zeng, S. Cancer-mutated ribosome protein L22 (RPL22/eL22) suppresses cancer cell survival by blocking p53-MDM2 circuit. Oncotarget 2017, 8, 90651–90661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.J.; Li, J.; Jia, Y.X.; Wu, J.J.; Yuan, L.; Li, M.J. Silencing of ribosomal protein L34 (RPL34) inhibits the proliferation and invasion of esophageal cancer cells. Oncol. Res. 2017, 25, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Ding, L.J.; Wei, Z.T.; Zhang, Y.D.; Li, Y.; Luo, Q.H. Ribosomal protein L34 promotes the proliferation, invasion and metastasis of pancreatic cancer cells. Oncotarget 2016, 7, 85259–85272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Gou, Y.; Wang, Q.; Jin, H.; Cui, L.; Zhang, Y. Downregulation of RPL6 by siRNA inhibits proliferation and cell cycle progression of human gastric cancer cell lines. PLoS ONE 2011, 6, e26401. [Google Scholar] [CrossRef]
- Guo, X.; Shi, Y.; Gou, Y.; Li, J.; Han, S.; Zhang, Y. Human ribosomal protein S13 promotes gastric cancer growth through down-regulating p27(Kip1). J. Cell Mol. Med. 2011, 15, 296–306. [Google Scholar] [CrossRef]
- Li, Y.; Dong, W.; Shi, Y.; Deng, F.; Chen, X.; Wan, C.; Zhou, M.; Zhao, L.; Fu, Z.F.; Peng, G. Rabies virus phosphoprotein interacts with ribosomal protein L9 and affects rabies virus replication. Virology 2016, 488, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Word, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 7, 621–628. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Length | Means-CK | Means-QT | log2(Fold Change) | Probability | Description |
|---|---|---|---|---|---|---|
| AFLA_020210 | 1362 | 370.1433333 | 64.65666667 | −2.517213007 | 0.885380738 | sexual development transcription factor NsdD |
| AFLA_048650 | 2100 | 156.1633333 | 38.95 | −2.00336052 | 0.844491613 | sexual development transcription factor SteA |
| AFLA_018540 | 1068 | 67.02666667 | 14.44666667 | −2.213998536 | 0.81499661 | G protein complex alpha subunit GpaB |
| AFLA_046990 | 2370 | 200.3033333 | 75.34333333 | −1.410634662 | 0.8049291 | APSES transcription factor StuA |
| AFLA_083110 | 255 | 766.0933333 | 24.60333333 | −4.960594472 | 0.950709 | conidiation-specific protein Con-10 |
| AFLA_044790 | 309 | 525.2933333 | 124.2433333 | −2.079954833 | 0.873234869 | conidiation-specific family protein |
| NO. | Gene Name | Means-CK | Means-QT | log2(Fold Change) | Up-Down-Regulation (QT/CK) | Description |
|---|---|---|---|---|---|---|
| 1 | AFLA_047870 | 2358.77 | 2.84 | −1.920818 | Down | 40S ribosomal protein S1 |
| 2 | AFLA_034340 | 976.7166667 | 6.739345 | −2.16111509 | Down | 40S ribosomal protein S10b |
| 3 | AFLA_050800 | 723.8966667 | 1.4697534 | −1.69897 | Down | 40S ribosomal protein S12 |
| 4 | AFLA_075030 | 802.26 | 9.62712 | −1.92081875 | Down | 40S ribosomal protein S13 |
| 5 | AFLA_044170 | 765.6133333 | 3.8280665 | −2.30102999 | Down | 40S ribosomal protein S14 |
| 6 | AFLA_044530 | 2185.433333 | 34.966928 | −1.795880017 | Down | 40S ribosomal protein S15 |
| 7 | AFLA_044110 | 763.1666667 | 9.9211671 | −1.886056647 | Down | 40S ribosomal protein S16 |
| 8 | AFLA_125890 | 2183.286667 | 43.66574 | −1.69897 | Down | 40S ribosomal protein S17 |
| 9 | AFLA_009000 | 1360.496667 | 34.012425 | −1.602059991 | Down | 40S ribosomal protein S18 |
| 10 | AFLA_050650 | 898.7666667 | 20.6716341 | −1.638272164 | Down | 40S ribosomal protein S19 |
| 11 | AFLA_117990 | 651.8966667 | 23.4682812 | −1.443697016 | Down | 40S ribosomal protein S2 |
| 12 | AFLA_043150 | 1318.843333 | 44.840662 | −1.468521082 | Down | 40S ribosomal protein S20 |
| 13 | AFLA_092120 | 8890.06 | 515.62348 | −1.236572001 | Down | 40S ribosomal protein S21 |
| 14 | AFLA_083740 | 680.39 | 6.8039 | −2.067506415 | Down | 40S ribosomal protein S22 |
| 15 | AFLA_021730 | 846.6866667 | 3.3867468 | −2.399868277 | Down | 40S ribosomal protein S23 |
| 16 | AFLA_071320 | 1901.14 | 7.60456 | −2.397940008 | Down | 40S ribosomal protein S24 |
| 17 | AFLA_083050 | 5390.12 | 29.64566 | −2.259637311 | Down | 40S ribosomal protein S25 |
| 17 | AFLA_127800 | 4272.09 | 26.059749 | −2.214670164 | Down | 40S ribosomal protein S26 |
| 18 | AFLA_083470 | 140.9033333 | 2.5362594 | −1.744727494 | Down | 40S ribosomal protein S28 |
| 19 | AFLA_101020 | 1328.043333 | 55.77806 | −1.37675071 | Down | 40S ribosomal protein S4 |
| 20 | AFLA_084620 | 1535.576667 | 16.89347 | −1.958607314 | Down | 40S ribosomal protein S5 |
| 21 | AFLA_026830 | 2783.25 | 125.24625 | −1.346787486 | Down | 40S ribosomal protein S6 |
| 22 | AFLA_029020 | 3209.673333 | 8954.783333 | −1.48148606 | Down | 40S ribosomal protein S8 |
| 23 | AFLA_101160 | 832.2533333 | 1.747731999 | −2.677780753 | Down | 40S ribosomal protein S9 |
| 24 | AFLA_030140 | 259.6833333 | 929.63 | 1.839903203 | Down | 60S acidic ribosomal protein P0 |
| 25 | AFLA_127860 | 264.4766667 | 10.05011333 | −1.420216403 | Down | 60S acidic ribosomal protein P1 |
| 26 | AFLA_044520 | 984.8466667 | 2.659086 | −2.568636235 | Down | 60S acidic ribosomal protein P2 |
| 27 | AFLA_068000 | 765.0433333 | 1.606590999 | −2.677780705 | Down | 60S ribosomal protein L1 |
| 28 | AFLA_112090 | 1100.716667 | 7.1546583 | −2.187086643 | Down | 60S ribosomal protein L11 |
| 29 | AFLA_080140 | 1298.606667 | 37.659593 | −1.537602002 | Down | 60S ribosomal protein L12 |
| 30 | AFLA_115110 | 943.4 | 9.24532 | −2.008773924 | Down | 60S ribosomal protein L13 |
| 31 | AFLA_056250 | 879.56 | 22.86856 | −1.585026652 | Down | 60S ribosomal protein L14 |
| 32 | AFLA_029260 | 633.1733333 | 16.14591999 | −1.593459819 | Down | 60S ribosomal protein L15, putative |
| 33 | AFLA_050000 | 1516.85 | 32.612275 | −1.66756154 | Down | 60S ribosomal protein L16 |
| 34 | AFLA_041990 | 1251.423333 | 4533.676667 | 1.857111595 | Up | 60S ribosomal protein L17 |
| 35 | AFLA_047440 | 2671.72 | 35.266704 | −1.879426068 | Down | 60S ribosomal protein L18 |
| 36 | AFLA_046970 | 2952.823333 | 8912.88 | 1.593796639 | Up | 60S ribosomal protein L19 |
| 37 | AFLA_048810 | 2272.563333 | 163.6245599 | −1.136677139 | Down | 60S ribosomal protein L2 |
| 38 | AFLA_029450 | 670.29 | 7.37319 | −1.958607314 | Down | 60S ribosomal protein L20 |
| 39 | AFLA_101150 | 2427.973333 | 41.2755466 | −1.769551078 | Down | 60S ribosomal protein L21 |
| 40 | AFLA_079880 | 233.58 | 1312.223333 | 2.490024624 | Up | 60S ribosomal protein L22 |
| 41 | AFLA_092370 | 1086.093333 | 13.25033866 | −1.913640169 | Down | 60S ribosomal protein L23 |
| 42 | AFLA_092370 | 1387.563333 | 70.76572998 | −1.292429823 | Down | 60S ribosomal protein L24 |
| 43 | AFLA_048140 | 2383.7 | 165.66715 | −1.155522824 | Down | 60S ribosomal protein L25 |
| 44 | AFLA_110470 | 188.4966667 | 9.481382335 | −1.298432014 | Down | 60S ribosomal protein L25, putative |
| 45 | AFLA_060150 | 2232.186667 | 13.39312 | −2.221848749 | Down | 60S ribosomal protein L26 |
| 46 | AFLA_127220 | 862.0833333 | 13.44849999 | −1.806875401 | Down | 60S ribosomal protein L27 |
| 47 | AFLA_054760 | 1163.723333 | 8.0296909 | −2.161150909 | Down | 60S ribosomal protein L28 |
| 48 | AFLA_103770 | 1127.98 | 10.039022 | −2.050609993 | Down | 60S ribosomal protein L28 |
| 49 | AFLA_134740 | 1447.586667 | 50.66553335 | −1.455931955 | Down | 60S ribosomal protein L3 |
| 50 | AFLA_045790 | 2011.47 | 43.246605 | −1.66756154 | Down | 60S ribosomal protein L30 |
| 51 | AFLA_045790 | 1371.076667 | 4046.166667 | −1.561246503 | Down | 60S ribosomal protein L31 |
| 52 | AFLA_003480 | 11,949.94667 | 33,519.81333 | 1.488009936 | Up | 60S ribosomal protein L32 |
| 53 | AFLA_086630 | 1485.253333 | 2.673455999 | −2.744727494 | Down | 60S ribosomal protein L33 |
| 54 | AFLA_086630 | 2061.426667 | 18.55284 | −2.04575749 | Down | 60S ribosomal protein L34 |
| 55 | AFLA_086630 | 1270.083333 | 5.715374999 | −2.346787486 | Down | 60S ribosomal protein L35 |
| 56 | AFLA_112390 | 832.9033333 | 4.164516667 | −2.301029995 | Down | 60S ribosomal protein L36 |
| 57 | AFLA_112390 | 3332.77 | 39.99324 | −1.920818753 | Down | 60S ribosomal protein L37 |
| 58 | AFLA_112390 | 1112.88 | 14.244864 | −1.89279003 | Down | 60S ribosomal protein L38 |
| 59 | AFLA_112390 | 1971.986667 | 29.77699867 | −1.821023052 | Down | 60S ribosomal protein L42 |
| 60 | AFLA_112390 | 3816.61 | 106.483419 | −1.554395796 | Down | 60S ribosomal protein L43 |
| 61 | AFLA_018700 | 1022.05 | 17.885875 | −1.732828271 | Down | 60S ribosomal protein L5 |
| 62 | AFLA_068420 | 3303.43 | 87.540895 | −1.576754126 | Down | 60S ribosomal protein L6 |
| 63 | AFLA_041710 | 1208.193333 | 5.255640998 | −2.361510743 | Down | 60S ribosomal protein L7 |
| 64 | AFLA_033980 | 1874.96 | 88.12312 | −1.327902142 | Down | 60S ribosomal protein L8 |
| 65 | AFLA_088370 | 686.0866667 | 3323.653333 | 2.276307178 | Up | 60S ribosomal protein L9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-M.; Li, Z.-Y.; Wang, Y.-D.; Wang, J.-Q.; Yang, P.-L. Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins 2019, 11, 154. https://doi.org/10.3390/toxins11030154
Li X-M, Li Z-Y, Wang Y-D, Wang J-Q, Yang P-L. Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins. 2019; 11(3):154. https://doi.org/10.3390/toxins11030154
Chicago/Turabian StyleLi, Xiu-Mei, Zhong-Yuan Li, Ya-Dong Wang, Jin-Quan Wang, and Pei-Long Yang. 2019. "Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus" Toxins 11, no. 3: 154. https://doi.org/10.3390/toxins11030154
APA StyleLi, X. -M., Li, Z. -Y., Wang, Y. -D., Wang, J. -Q., & Yang, P. -L. (2019). Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins, 11(3), 154. https://doi.org/10.3390/toxins11030154