Antiparasitic Properties of Cantharidin and the Blister Beetle Berberomeloe majalis (Coleoptera: Meloidae)


 ,
, 
Abstract
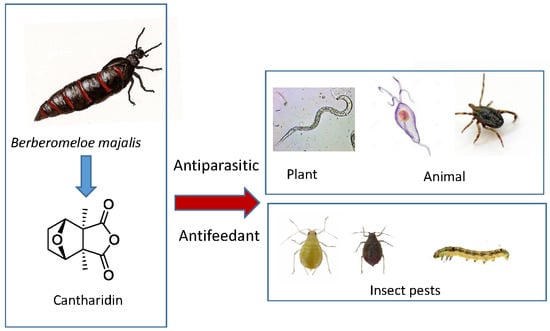
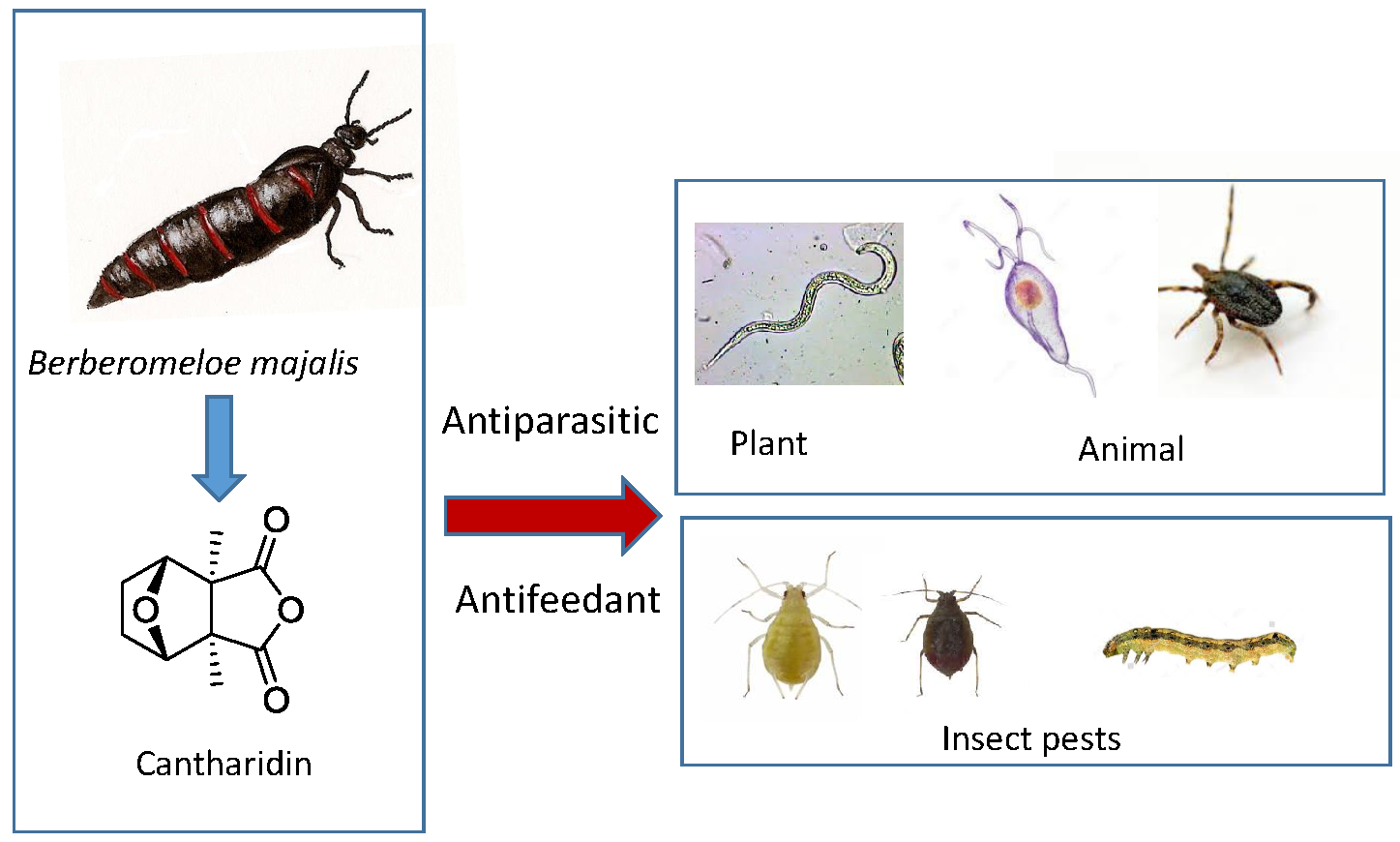
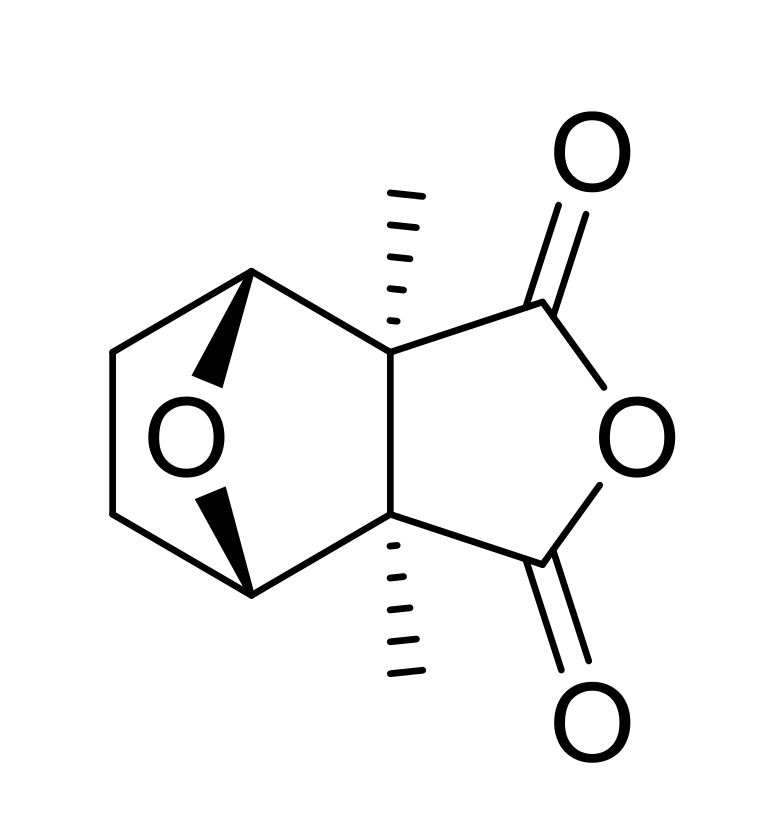
:
1. Introduction
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Insect Extracts
4.2. Cantharidin Quantification
4.3. Bioassays
4.4. Antiprotozoal Activity
4.5. Nematicidal Activity
4.6. Insect Bioassays
4.7. Ixodicidal Activity
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gathercoal, E.N.; Wirth, E.H. Pharmacognosy; Lea and Febiger: Philadelphia, PA, USA, 1936; pp. 1–852. [Google Scholar]
- Wang, G.S. Medical uses of mylabris in ancient China and recent studies. J. Ethnopharmacol. 1989, 26, 147–162. [Google Scholar] [CrossRef]
- Wilson, C.R. Methods for Analysis of Gastrointestinal Toxicants. In Comprehensive Toxicology, 2nd ed.; Hooser, S., Mc-Queen, C., Eds.; Elsevier Academic Press: London, UK, 2010; pp. 145–152. [Google Scholar]
- Moed, L.; Shwayder, T.A.; Chang, M.W. Cantharidin revisited: A blistering defense of an ancient medicine. Arch. Dermatol. 2001, 137, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Pajovic, B.; Radosavljevic, M.; Radunovic, M.; Radojevic, N.; Bjelogrlic, B. Arthropods and their products as aphrodisiacs—Review of literature. Eur. Rev. Med. Pharmaco. 2012, 16, 539–547. [Google Scholar]
- James, P.; Thorpe, N. Ancient Inventions; Ballantine Books; The Random House Publishing Group: New York, NY, USA, 1995; pp. 1–672. [Google Scholar]
- Ford, P.; Howell, M. The Beetle of Aphrodite and other Medical Mysteries; Random House: New York, NY, USA, 1985; pp. 1–358. [Google Scholar]
- Schaeffer, N. The Marquis de Sade: A Life; Harvard University Press: Cambridge, MA, USA, 2000; pp. 1–557. [Google Scholar]
- Robiquet, M. Expériences sur les cantharides. Annales de Chimie 1810, 76, 302–322. [Google Scholar]
- Aggrawal, A. Spanish Fly (Cantharides). In Textbook of Forensic Medicine and Toxicology; Avichal Publishing Company: New Delhi, India, 2014; p. 652. [Google Scholar]
- Blood, D.C.; Studdert, V.P.; Gay, C.C. Saunders Comprehensive Veterinary Dictionary, 3rd ed.; Elsevier: Philadelphia, PA, USA, 2007; pp. 1–2172. [Google Scholar]
- Anon. Cantharide. In Farlex Partner Medical Dictionary; Farlex: Huntingdon Valley, PA, USA, 2012; Available online: http://medical-dictionary.thefreedictionary.com/ (accessed on 31 January 2017).
- Taberner, P.V. Aphrodisiacs: The Science and the Myth; Springer Science & Business Media: Bristol, UK, 2012; pp. 1–278. [Google Scholar]
- Puerto-Galvis, C.E.; Vargas-Méndez, L.Y.; Kouznetsov, V.V. Cantharidin-Based Small Molecules as Potential Therapeutic Agents. Chem. Biol. Drug Des. 2013, 82, 477–499. [Google Scholar] [CrossRef] [PubMed]
- Karras, D.J.; Farrell, S.E.; Harrigan, R.A.; Henretig, F.M.; Gealt, L. Poisoning from “Spanish Fly” (cantharidin). Am. J. Emerg. Med. 1996, 14, 478–483. [Google Scholar] [CrossRef]
- Ghoneim, K. Cantharidin toxicosis to animal and human in the world: A review. Stand. Res. J. Toxicol. Environ. Health Sci. 2013, 1, 1–16. [Google Scholar]
- Cotovio, P.; Silva, C.; Marques, M.G.; Ferrer, F.; Costa, F.; Carreira, A.; Campos, M. Acute kidney injury by cantharidin poisoning following a silly bet on an ugly beetle. Clin. Kidney J. 2013, 6, 201–203. [Google Scholar] [CrossRef]
- Schmitz, D.G. Overview of Cantharidin Poisoning (Blister Beetle Poisoning). In The Merck Veterinary Manual; Aiello, S.E., Moses, M.A., Eds.; Merck Sharp & Dohme: Kenilworth, NJ, USA, 2013; Available online: http://www.merckvetmanual.com/ (accessed on 31 January 2017).
- Nickolls, L.C.; Teare, D. Poisoning by cantharidin. Br. Med. J. 1954, 2, 384–1386. [Google Scholar] [CrossRef]
- Froberg, B.A. Animals. In Criminal Poisoning: Clinical and Forensic Perspectives; Jones & Bartlett: Burlington, MA, USA, 2010; pp. 39–48. [Google Scholar]
- Al-Rumikan, A.; Al-Hamdan, N.A. Indirect cantharidin food poisoning caused by eating wild birds. Saudi Epidemiol. Bulll. 1999, 6, 25–26. [Google Scholar]
- Penrith, M.L.; Naudé, T.W. Mortality in chickens associated with blíster beetle consumption. J. S. Afr. Vet. Assoc. 1996, 67, 97–99. [Google Scholar]
- Rockett, J.; Bosted, S. Veterinary Clinical Procedures in Large Animal Practices; Cengage Learning: Boston, MA, USA, 2015; pp. 1–672. [Google Scholar]
- Honkanen, R.E. Cantharidin, another natural toxin that inhibits the activity of serine/threonine protein phosphatases types 1 and 2ª. FEBS Lett. 1993, 330, 283–286. [Google Scholar] [CrossRef]
- Baba, Y.; Hirukawa, N.; Sodeoka, M. Optically active cantharidin analogues possessing selective inhibitory activity on Ser/Thr protein phosphatase 2B (calcineurin): Implications for the binding mode. Bioorgan. Med. Chem. 2005, 13, 5164–5170. [Google Scholar] [CrossRef]
- Bologna, M.A.; Oliverio, M.; Pitzalis, M.; Mariottini, P. Phylogeny and evolutionary history of the blister beetles (Coleoptera, Meloidae). Mol. Phylogenet. Evol. 2008, 48, 679–693. [Google Scholar] [CrossRef]
- Carrel, J.E.; Eisner, T. Cantharidin: Potent feeding deterrent to insects. Science 1974, 183, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Nikbakhtzadeh, N.R.; Dettner, K.; Boland, W.; Gäde, G.; Dötterle, S. Intraspecific transfer of cantharidin within selected members of the family Meloidae (Insecta: Coleoptera). J. Insect. Physiol. 2007, 53, 890–899. [Google Scholar] [CrossRef]
- Eisner, T.; Conner, J.; Carrel, J.E.; McCormick, J.P.; Slagle, A.J.; Gans, C.; O’Reilly, J.C. Systemic retention of ingested cantharidin by frogs. Chemoecology 1990, 1, 57–62. [Google Scholar] [CrossRef]
- Bartram, S.; Boland, W. Chemistry and ecology of toxic birds. ChemBioChem 2001, 2, 809–811. [Google Scholar] [CrossRef]
- Wirtz, W.O.; Austin, D.H.; Dekle, G.W. Food habits of the common long-nosed armadillo Dasypus movemcinctus in Florida, 1960–1961. In Evolution and Ecology of Armadillos, Sloths, and Vermilinguas; Montgomery, G.G., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1985; pp. 439–451. [Google Scholar]
- Larson, N.P. The common toad as an enemy of blíster beetles. J. Econ. Entomol. 1943, 36, 480. [Google Scholar] [CrossRef]
- Kilham, L. Use of blíster beetle in bill-sweeping by White-brested Nuthatch. Auk Ornithol. Adv. 1971, 88, 175–176. [Google Scholar]
- Percino-Daniel, N.; Buckley, D.; García-París, M. Pharmacological properties of blister beetles (Coleoptera: Meloidae) promoted their integration into the cultural heritage of native rural Spain as inferred by vernacular names diversity, traditions, and mitochondrial DNA. J. Ethnopharmacol. 2013, 147, 570–583. [Google Scholar] [CrossRef]
- Sánchez-Barbudo, I.S.; Camarero, P.R.; García-Montijano, M.; Mateo, R. Possible cantharidin poisoning of a great bustard (Otis tarda). Toxicon 2012, 59, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Bravo, C.; Bautista, L.M.; García-París, M.; Blanco, G.; Alonso, J.C. Males of a Strongly Polygynous Species Consume More Poisonous Food than Females. PLoS ONE 2014, 9, e111057. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.A. Diseases and Medical Management of Houbara Bustards and other Otididae; Emirates Printing Press LLC: Dubai, UAE, 2008; pp. 1–494. [Google Scholar]
- Silvanose, C.; Samour, J.; Naldo, J.; Bailey, T.A. Oro-pharyngeal protozoa in captive bustards: Clinical and pathological considerations. Avian. Pathol. 1998, 27, 526–530. [Google Scholar] [CrossRef]
- Alonso, J.C.; Palacín, C. Avutarda—Otis tarda. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2015; Available online: http://www.vertebradosibericos.org/ (accessed on 31 January 2017).
- Cordero del Campillo, M.; Castañón-Ordóñez, L.; Reguera-Feo, A. Índice Catálogo de Zooparásitos Ibéricos, 2nd ed.; Universidad de León, Secretariado de Publicaciones: León, Spain, 1994; pp. 1–650. [Google Scholar]
- McCormick, J.P.; Carrel, J.E. Cantharidin biosynthesis and function in meloid beetles. In Pheromone Biochemistry; Prestwich, G.D., Bloomquist, H.F., Eds.; Harcourt, Brace, & Jovanovich: Orlando, FL, USA, 1987; pp. 307–350. [Google Scholar]
- Carrel, J.E.; McCairel, M.H.; Slagle, A.J.; Doom, J.P.; Brill, J.; McCormick, J.P. Cantharidin production in a blister beetle. Experientia 1993, 49, 171–174. [Google Scholar] [CrossRef]
- Mebs, D.; Pogoda, W.; Schneider, M.; Kauert, G. Cantharidin and demethylcantharidin (palasonin) content of blister beetles (Coleoptera: Meloidae) from southern Africa. Toxicon 2009, 53, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Whitman, D.W. Allelochemical interactions among plants, herbivores, and their predators. In Novel Aspects of Insect-Plant Interactions; Barbosa, P., Letrourneau, D., Eds.; John Wiley: New York, NY, USA, 1988; pp. 11–64. [Google Scholar]
- De Brum Vieira, P.; Brandt Giordani, R.; Macedo, A.J.; Tasca, T. Natural and synthetic compound anti-Trichomonas vaginalis: An update review. Parasitol. Res. 2015, 114, 1249–1261. [Google Scholar] [CrossRef]
- Yahya, M.; Fatemeh, G.; Abdolhosein, D.; Zohreh, S.; Zuhair, H. Effect of cantharidin on apoptosis of the Leishmania major and on parasite load in BALB/c mice. Res. J. Parasitol. 2013, 8, 14–25. [Google Scholar]
- Ghaffarifar, F. Leishmania major: In vitro and in vivo anti-leishmanial effect of cantharidin. Exp. Parasitol. 2010, 126, 126–129. [Google Scholar] [CrossRef]
- Bajsa, J.; McCluskey, A.; Gordon, C.P.; Stewart, S.G.; Sahu, R.; Duke, S.O.; Tekwan, B.L. The antiplasmodial activity of norcantharidin analogs. Bioorg. Med. Chem. Lett. 2010, 20, 6688–6695. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.E.; Tarleton, M.; Gordon, C.P.; Sakoff, J.A.; Gilbert, J.; McCluskey, A.; Gasser, R.B. Norcantharidin analogues with nematocidal activity in Haemonchus contortus. Bioorg. Med. Chem. Lett. 2011, 21, 3277–3281. [Google Scholar] [CrossRef]
- Campbell, B.E.; Hofmann, A.; McCluskey, A.; Gasser, R.B. Serine/threonine phosphatases in socioeconomically important parasitic nematodes-prospects as novel drug targets? Biotechnol. Adv. 2011, 29, 28–39. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, Y. Chronic sublethal effects of cantharidin on the diamondback moth Plutella xylostella (lepidoptera: Plutellidae). Toxins 2015, 7, 1962–1978. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y. Characterization of glutathione S-transferases from Sus scrofa, Cydia pomonella and Triticum aestivum: Their responses to cantharidin. Enzyme Microb. Technol. 2015, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lü, S.; Zhang, Y. Characterization of protein phosphatase 5 from three lepidopteran insects: Helicoverpa armigera, Mythimna separata and Plutella xylostella. PLoS ONE 2014, 9, e97437. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, J.; Zhang, Y. Cantharidin impedes the activity of protein serine/threonine phosphatase in Plutella xylostella. Mol. BioSyst. 2014, 10, 240–250. [Google Scholar] [CrossRef]
- Wang, M.; Nan, X.; Feng, G.; Yu, H.; Hu, G.; Liu, Y. Design, synthesis and bioactivity evaluation of novel acylthiourea derivatives of cantharidin. Ind. Crop. Prod. 2014, 55, 11–18. [Google Scholar] [CrossRef]
- Ruiz-Vásquez, L.; Olmeda, A.S.; Zúñiga, G.; Villarroel, L.; Echeverri, L.F.; González-Coloma, A.; Reina, M. Insect antifeedant and ixodicidal compounds from Senecio adenotrichius. Chem. Biodivers. 2017, 14, 1612–1880. [Google Scholar] [CrossRef] [PubMed]
- Till, J.S.; Majmudar, B.N. Cantharidin poisoning. South. Med. J. 1981, 74, 444–447. [Google Scholar] [CrossRef]
- Polettini, A.; Crippa, O.; Ravagli, A.; Saragoni, A. A fatal case of poisoning with cantharidin. Forensic. Sci. Int. 1992, 56, 37–43. [Google Scholar] [CrossRef]
- Martínez-Díaz, R.A.; Ibáñez-Escribano, A.; Burillo, J.; de las Heras, L.; del Prado, G.; Agulló-Ortuño, M.T.; Julio, L.F.; González-Coloma, A. Trypanocidal, trichomonacidal and cytotoxic components of cultivated Artemisia absinthium Linnaeus (Asteraceae) essential oil. Mem. Inst. Oswaldo Cruz. 2015, 110, 639–699. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Escribano, A.; Meneses Marcel, A.; Machado Tugores, Y.; Nogal Ruiz, J.J.; Arán Redó, V.J.; Escario García-Trevijano, J.A.; Gómez Barrio, A. Validation of a modified fluorimetric assay for the screening of trichomonacidal drugs. Mem. Inst. Oswaldo Cruz. 2012, 107, 637–643. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematocidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Burgueño-Tapia, E.; Castillo, L.; González-Coloma, A.; Joseph-Nathan, P. Antifeedant and phytotoxic activity of the sesquiterpene p-benzoquinone perezone and some of its derivatives. J. Chem. Ecol. 2008, 34, 766–771. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Extract | CTD (µg/mg) | Total CTD (mg) a | Distribution of Total CTD (%) | CTD Per Beetle (mg) b |
|---|---|---|---|---|
| Body | 41.2 | 1813 | 99.3 | 9.06 |
| Hemolymph | 295.0 | 5.9 | 0.7 | 0.03 |
| Total | - | 1819 | 100 | 9.1 |
| Concentration (µg/mL) | Body Extract | Hemolymph Extract | CTD | Metronidazole |
|---|---|---|---|---|
| 500 | 92.7 ± 0.8 | 99.8 ± 0.3 | - | - |
| 100 | 52.7 ± 4.6 | 77.5 ± 4.1 | 98.1 ± 0.3 | - |
| GI50 (µg/mL) (95% CL) | 75.7 (24.6–220.2) | 15.5 (1.4–36.2) | 5.6 (4.2–7.0) | 0.6 (0.3–1.4) |
| Treatment | Dose (µg/µL) | Mortality a % | Lethal Concentrations b | |
|---|---|---|---|---|
| LC50 (µg/mg) | LC90 (µg/mg) | |||
| Body | 1 | 74.9 ± 2.92 | nc | |
| Hemolymph | 1 | 84.05 ± 2.64 | 0.656 (0.626–0.687) | 1.108 (1.054–1.172) |
| CTD | 0.5 | 100 ± 0 | 0.0252(0.023–0.027) | 0.065 (0.061–0.070) |
| Treatment | Concentration (µg/cm2) | Rhopalosiphum Padi | Myzus Persicae |
|---|---|---|---|
| %SI b | |||
| Body | 50 | 94.35 ± 2.42 | 82.75 ± 8.28 |
| EC50 b | 6.7 (4.63–9.63) | 14.3 (8.1–25.3) | |
| Hemolymph | 50 | 96.84 ± 1.94 | 93.23 ± 5.0 |
| EC50 b | 0.8 (0.5–1.5) | 3.38 (1.98–5.77) | |
| CTD | 50 | 94.7 ± 3.5 | 91.50 ± 2.31 |
| EC50 b | 0.098 (0.031–0.3) | 0.211 (0.05–0.91) | |
| Treatment | Mortality a | Lethal Concentrations b | |
|---|---|---|---|
| LC50 (µg/mg) | LC90 (µg/mg) | ||
| Body | 81.7 ± 0.9 | 12.79 (10.84–14.93) | 23.93 (20.79–28.94) |
| Hemolymph | 70.2 ± 1.8 | 12.25 (10.65–13.94) | 21.05 (18.74–24.47) |
| CTD | 90 ± 0.1 | 12.84 (11.55–14.30) | 20.31 (18.32–23.11) |
| Nootkatone c | - | 4.02 (1.92–7.42) | 18.02 (13.60–29.16) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitman, D.W.; Andrés, M.F.; Martínez-Díaz, R.A.; Ibáñez-Escribano, A.; Olmeda, A.S.; González-Coloma, A. Antiparasitic Properties of Cantharidin and the Blister Beetle Berberomeloe majalis (Coleoptera: Meloidae). Toxins 2019, 11, 234. https://doi.org/10.3390/toxins11040234
Whitman DW, Andrés MF, Martínez-Díaz RA, Ibáñez-Escribano A, Olmeda AS, González-Coloma A. Antiparasitic Properties of Cantharidin and the Blister Beetle Berberomeloe majalis (Coleoptera: Meloidae). Toxins. 2019; 11(4):234. https://doi.org/10.3390/toxins11040234
Chicago/Turabian StyleWhitman, Douglas W., Maria Fe Andrés, Rafael A. Martínez-Díaz, Alexandra Ibáñez-Escribano, A. Sonia Olmeda, and Azucena González-Coloma. 2019. "Antiparasitic Properties of Cantharidin and the Blister Beetle Berberomeloe majalis (Coleoptera: Meloidae)" Toxins 11, no. 4: 234. https://doi.org/10.3390/toxins11040234
APA StyleWhitman, D. W., Andrés, M. F., Martínez-Díaz, R. A., Ibáñez-Escribano, A., Olmeda, A. S., & González-Coloma, A. (2019). Antiparasitic Properties of Cantharidin and the Blister Beetle Berberomeloe majalis (Coleoptera: Meloidae). Toxins, 11(4), 234. https://doi.org/10.3390/toxins11040234